
Abstract
Verbal working memory consists of separable capacities for the retention of phonological and semantic information. Within the phonological domain, there are independent capacities for retaining input-phonological codes and output-phonological codes. The input-phonological capacity does not appear to be critical for language comprehension but is involved in verbatim repetition and long-term learning of new words. The semantic capacity is critical for both comprehension and production and for the learning of new semantic information. Different neural structures appear to underlie these capacities, with a left-parietal region involved in input-phonological retention and a left-frontal region involved in semantic retention.
Most cognitive psychologists take for granted the notion that language comprehension draws on working memory (the system that stores information temporarily and allocates attention to the processing of this information). Traditionally, memory span (or the number of words that can be recalled in order) has been used as a measure of the storage-capacity component of working memory, often referred to as short-term memory (STM) capacity. Neurally intact individuals have a span of about five words. Thus, one might expect that a brain-damaged patient or a child with a developmental disorder who had a memory span of only one to two words would, as a result, have problems with understanding language. Many studies from our lab have shown, however, that whether or not there is a connection between reduced span and difficulties with language processing depends on the specific source of the STM deficit and the specific type of language task (Freedman & Martin, 2001; Hanten & Martin, 2000; Martin & He, 2004). We have developed a multiple-component model of verbal STM to accommodate these findings (Martin, in press). This multiple-component approach was first applied to understanding the relation between STM and sentence comprehension and repetition, but has been expanded more recently to encompass the relation between STM and language production and verbal learning. We have also begun to investigate more closely the neural architecture that underlies these different capacities through lesion studies and functional neuroimaging.
PHONOLOGICAL VERSUS SEMANTIC STORAGE COMPONENTS
A large body of findings indicates that individuals use a phonological (i.e., speech-sound) code to maintain information in short-term memory. For example, memory span is smaller for lists of phonologically similar words (e.g., rhyming words) than for phonologically dissimilar words and smaller for lists of multisyllable words than for single-syllable words. These effects hold irrespective of whether the lists are presented auditorily or visually. These findings have led to models of verbal STM in which words are stored in a phonological form and rehearsed subvocally. A number of individuals with brain damage or developmental disorders whose memory-span performance implicates a disruption of phonological storage have been reported (Hanten & Martin, 2000; Martin, in press; Vallar & Papagno, 1995). That is, despite showing accurate perception and production of speech sounds, these patients fail to show the standard phonological effects on span, and have particular difficulty repeating lists of nonwords (e.g., “dember,”“plim,” etc.)—an ability that depends mainly on retaining phonological information. For example, patient EA had a word span of one to two items, but could only repeat a nonword if it had three phonemes or fewer. However, not all patients with reduced span show this pattern. Some show standard phonological effects but fail to show normal retention of semantic information, despite having excellent knowledge of word meanings. For example, these patients fail to show the normal advantage for words over nonwords on span tasks—suggesting that they cannot make use of the lexical and semantic information in words to aid their recall (Martin & He, 2004). These differing STM patterns indicate that the retention of phonological and semantic information depends on separate capacities, which may be differentially affected by brain damage.
A further differentiation of STM capacities appears to be required within the phonological domain. We and other researchers have argued that there is one capacity for the retention of “input” phonological codes—those derived from speech perception—and another for the retention of “output” phonological codes—those generated internally that serve as the basis for speech production (Martin, Lesch, & Bartha, 1999). Supporting this contention are patients who perform well on STM tasks when they do not have to reproduce the list (as on probe tasks), but perform poorly on all list-repetition tasks. A case study that we reported (Martin et al., 1999) showed this pattern. Patient MS is a classic anomic patient in that he is a fluent speaker but has difficulty producing names for objects and producing nouns in spontaneous speech, though he shows good comprehension of the words he cannot produce. In picture naming, he often produces circumlocutions—that is, he accurately describes the object to be named, but cannot produce its name. On list-memory tasks that did not involve reproducing the list (e.g., deciding whether two lists were the same or different), he performed at a normal level. However, when asked to reproduce lists, he performed poorly and often produced circumlocutions like those he produced in naming. Thus, MS demonstrated preserved input-phonological capacity (supporting his excellent performance on probe and matching tasks), but poor access to output phonology, which impaired his performance in overt recall.
STM AND LANGUAGE PROCESSING
We and others have found that patients with an input-phonological-storage deficit perform remarkably well on sentence comprehension, even for sentences in which many words intervene between words that must be linked (such as between a noun and a reflexive pronoun; Butterworth, Campbell, & Howard, 1986; Hanten & Martin, 2000; Martin & He, 1994). Thus, the general conclusion in the literature is that semantic and syntactic codes are accessed quickly, with the meaning of the sentence constructed word-by-word as each word is perceived, to the extent that is possible. Retention of the phonological codes for the words is not necessary to support sentence comprehension (Hanten & Martin, 2000; Martin, in press). A semantic-STM deficit, in contrast, does lead to difficulty comprehending certain sentence types—specifically, those that require the maintenance of several individual word meanings simultaneously prior to their integration with other words (Martin, in press; Martin & He, 2004).
On the language-production side, patients with input-phonological-STM deficits have demonstrated normal language production on a variety of tasks (Martin, in press). Patients with output-phonological deficits produce phonologically disordered output (see Martin et al., 1999); however, there has been little systematic analysis of their production deficits at the phrase and sentence levels. Patients with semantic-STM deficits have been shown to have considerable difficulty in phrase and sentence production (Martin, in press). For example on a task that required the production of adjective–noun and adjective–adjective–noun phrases to describe pictures, a patient with an input-phonological-STM deficit (EA) performed at a normal level whereas two patients with a semantic-STM deficit (AB and ML) performed very poorly (despite their being able to produce the adjectives and nouns in isolation). For example, in trying to produce “short hair,” patient AB said, “Well … that's hair. It's short. That's short. … I can't get it.” To explain these results, we have argued that speakers plan their utterances on a phrase-by-phrase basis. They access and maintain lexical-semantic representations for all of the content words in a phrase simultaneously prior to beginning articulation of the utterance. Patients with a semantic-STM deficit have difficulty maintaining several lexical-semantic representations simultaneously, leading to halting, piecemeal speech.
STM, VERBATIM RECALL, AND LEARNING
From the discussion above, the question arises as to what the role of the input-phonological STM might be, given the good language comprehension and production of patients with such STM deficits. In fact, severe deficits on some language tasks have been observed for these patients—specifically, on those tasks that require a verbatim phonological record. These patients are very impaired in verbatim sentence repetition, often producing a paraphrase of a sentence rather than the exact wording. The patients with phonological-STM deficits have also been shown to have difficulty in learning new phonological forms—that is, new words. In a recent study, we showed that patients with a phonological-STM deficit have difficulty learning foreign translations of English words (Freedman & Martin, 2001). However, they do better in learning a new meaning for an old word (e.g., rip→“old horse”). Patients with a semantic-STM deficit show the reverse—that is, doing better on learning new phonological forms but worse on learning new meanings. Thus, there appears to be a specific relation between holding information in either a phonological or semantic form in STM and being able to create a long-term-memory representation for that information.
These findings on learning have implications for vocabulary acquisition in children and adults with neurological disorders. Children with reduced phonological or semantic capacity either due to brain injury or developmental disorders are likely to have difficulty acquiring words at a normal rate (Gathercole & Baddeley, 1990). Stroke patients with reduced phonological- or semantic-STM capacities may also have difficulty reacquiring lost word meanings or pronunciations because of these capacity restrictions. Work in other laboratories has been investigating whether word-learning methodologies that emphasize the priming of either semantic or phonological representations may be differentially effective depending on the nature of the patients' STM deficit (e.g., N. Martin, Fink, Laine, & Ayala, 2004).
NEUROANATOMICAL BASIS OF VERBAL STM
Reviews of studies of patients with input-phonological-STM deficits indicate that these patients have lesions in the left temporal and parietal lobes of the brain (see Fig. 1), with the region of greatest overlap across all patients with this disorder being in the left supramarginal gyrus (Vallar & Papagno, 1995). The cases in our lab with semantic-STM deficits have had left inferior frontal lesions, though the most striking cases also had damage in adjacent parietal regions. The left supramarginal gyrus and the left inferior frontal regions involved in STM differ from those thought to be involved in knowledge representations for phonological and semantic information. Specifically, the representations of word meanings (at least for concrete nouns) appear to be localized in left middle and inferior temporal regions, and phonological representations in left superior temporal regions (see Martin, 2003, for discussion). Consequently, one might hypothesize that during short-term retention of several words, semantic representations in the inferior and middle temporal lobe are activated and temporarily connected to regions in the inferior frontal lobe for maintenance whereas input-phonological representations in the superior temporal lobe are activated and temporarily connected to the supramarginal gyrus (Hanten & Martin, 2000; see Fig. 1). A functional neuroimaging study from our lab, using normal undergraduate participants, examined input-phonological and semantic retention by contrasting performance on a rhyme-probe task (i.e., judging whether a probe word rhymes with a list item) and a synonym-probe task (i.e., judging whether a probe word is a synonym of a list item; Martin, Wu, Jackson, Freedman, & Lesch, 2003). As shown in Figure 2, a large left inferior and midfrontal region showed a strong effect of STM load, as it was more highly activated for both tasks when the list length was four items compared to one item. Some differences in the activation patterns for the two tasks were obtained, however, as shown in Figure 3. Consistent with the patient findings, an inferior parietal region was more strongly activated in the phonological task than in the semantic task. Although a frontal region was more activated in the semantic task, this activation was not significant when using a conservative criterion for statistical significance that adjusted for the many comparisons of activation in specific brain regions for the two tasks. Neuroimaging findings from other labs have, however, found significant left frontal involvement in maintaining semantic representations, contrasting with left parietal involvement in maintaining input-phonological representations (Shivde & Thompson-Schill, 2004), though the specific regions differed somewhat from those reported in our study. There is little patient lesion data or evidence from neuroimaging from which to draw conclusions about the localization of output-phonological storage as distinct from output representations for words and motor programming. Further research is clearly needed to separate the localization of processing and storage for output phonology.
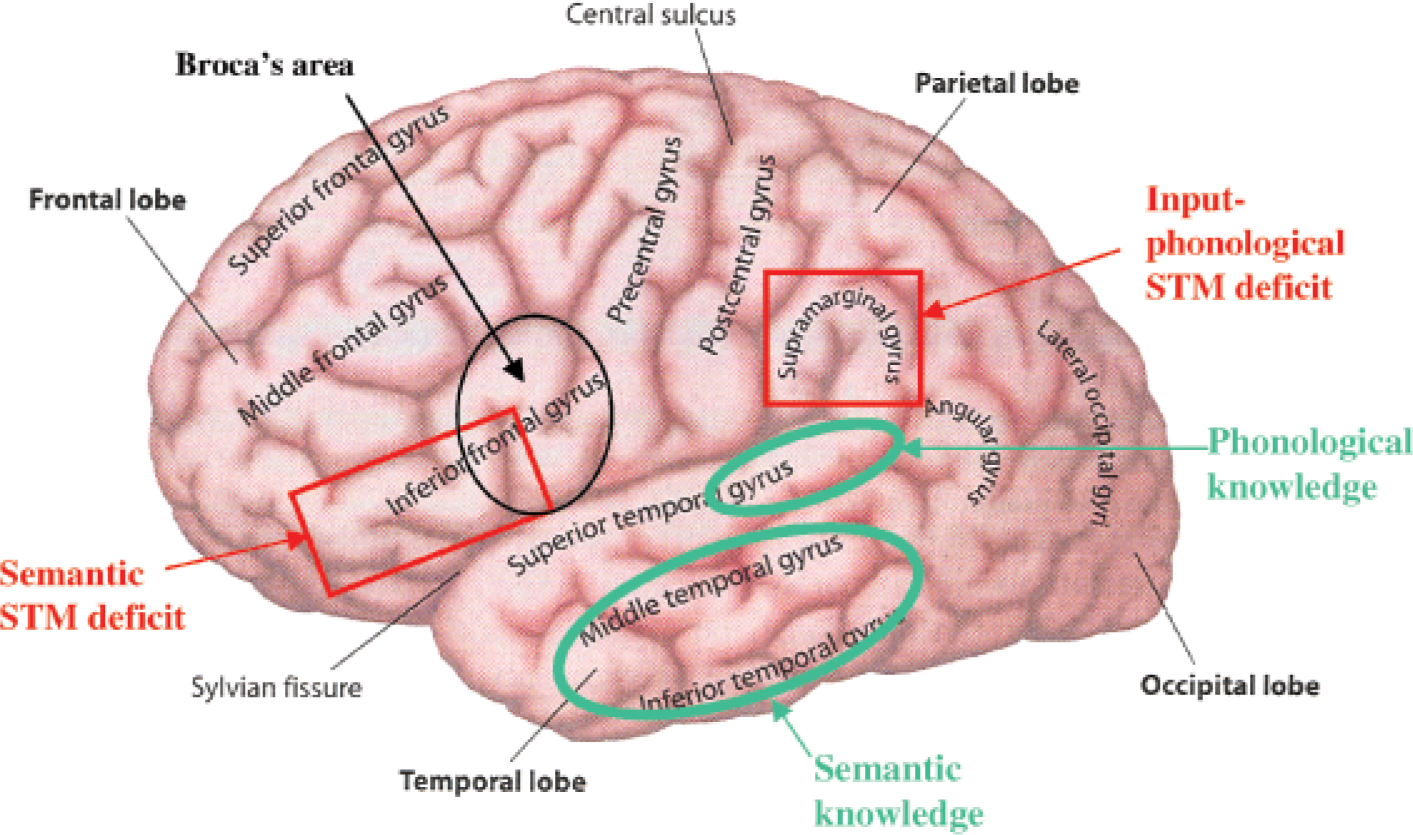
Cortical areas of the left hemisphere involved in phonological and semantic storage and processing. Areas circled in green are regions thought to be involved in the representation of phonological and semantic knowledge. Areas outlined in red are regions identified as being involved in semantic- and phonological-short-term memory (STM) deficits and, consequently, thought to be involved in short-term storage. Broca's area (outlined in black) is also shown, due to its overlap with the region hypothesized to be involved in semantic STM. Figure is adapted from Cognitive neuroscience: The biology of the mind (p. 45), by M.S. Gazzaniga, R.B. Ivry, & G.R. Mangun, 1998, New York: W.W. Norton & Co.
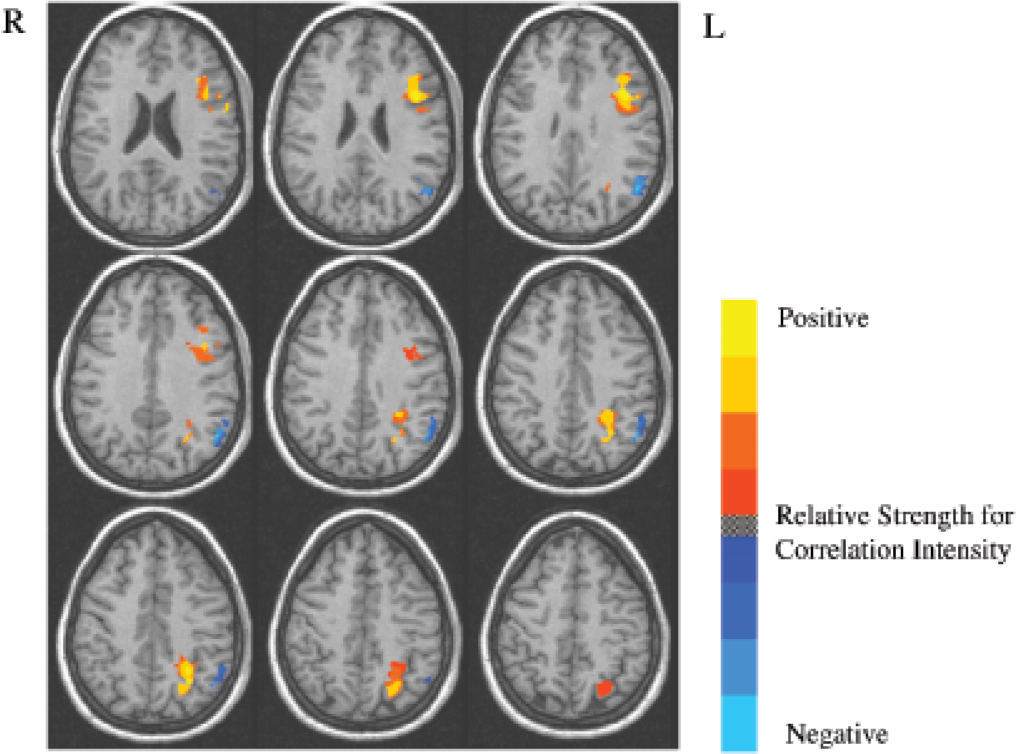
Brain activation patterns (horizontal slices 3 mm apart; from 22 – 46 mm above the horizontal plane separating the hypothalamus from subthalamic regions) showing contrast in short-term-memory (STM) load during rhyme- and synonym-probe tasks. Yellow/red areas were more activated in both tasks when they involved lists of four items than when they involved just a single item, whereas blue areas were more activated in the one-item conditions than in the four-item conditions. Reprinted from “An event-related fMRI investigation of phonological vs. semantic short-term memory,” by R.C. Martin, D. Wu, D. Jackson, M. Freedman, & M. Lesch, Journal of Neurolinguistics, Vol. 16, p. 350, copyright 2003, with permission from Elsevier.
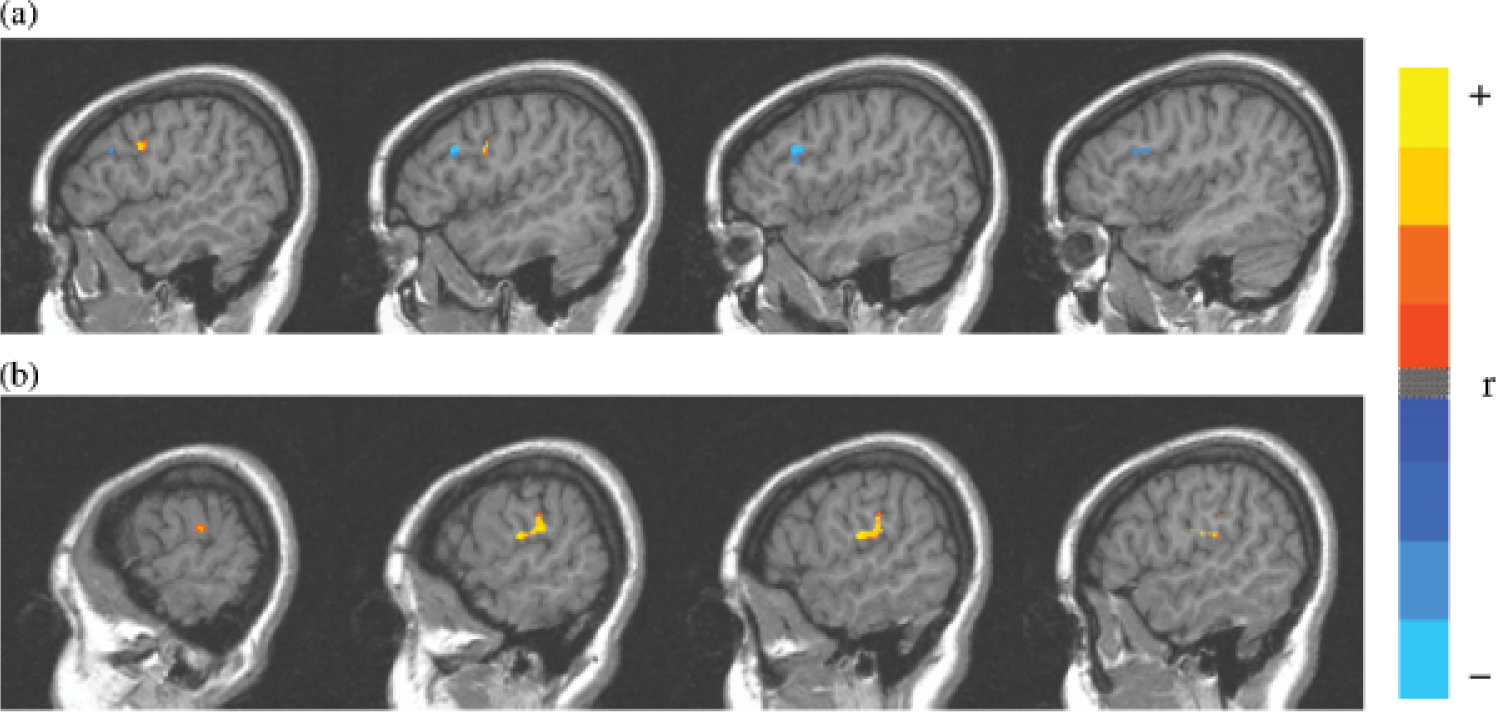
Brain activation patterns (vertical slices, 3 mm apart from 53 – 62 mm to the left of the plane dividing the left hemisphere from the right) showing contrasting activation in regions of interest for synonym and rhyme tasks. Greater anterior frontal activation was observed for the synonym conditions (indicated as the blue cluster; a), whereas greater posterior activation was observed for the rhyme conditions (indicated as the yellow/red cluster; b). Reprinted from “An event-related fMRI investigation of phonological vs. semantic short-term memory,” by R.C. Martin, D. Wu, D. Jackson, M. Freedman, & M. Lesch, Journal of Neurolinguistics, Vol. 16, p. 352, copyright 2003, with permission from Elsevier.
In discussions of the involvement of inferior frontal regions in semantic-STM deficits, questions often arise regarding the extent of overlap of these regions with Broca's area (see Fig. 1) and the extent to which such semantic-STM patients would be classified as Broca's aphasics. Patients with semantic STM might be seen as qualifying for the syndrome given their good single-word comprehension and their halting and fragmented speech. However, unlike the prototypical Broca's aphasic, these patients are not agrammatic speakers (that is, they do not omit grammatical words and inflections) and do not have problems with articulation (Martin & He, 2004). As discussed at length elsewhere, there is limited value in classifying patients into traditional syndromes because of the heterogeneity of deficits across patients in any of these groups (see Coltheart, 2001). In anatomical terms, the role of Broca's area in causing the deficits typically ascribed to Broca's aphasia is questionable, as a wide lesion involving the underlying cortex appears to be required (see Martin, 2003, for discussion). Patients who are classified as Broca's aphasics may have lesions that encompass the region involved in semantic-STM deficits and some of their speech-production symptoms (e.g., reduced phrase length) may derive from a semantic-STM deficit. However, they typically have additional deficits in speech motor programming and grammatical aspects of speech not found in patients with isolated semantic-STM deficits.
FUTURE DIRECTIONS
Further neuroimaging studies are underway to determine whether the left inferior frontal and supramarginal gyri play the roles in language comprehension and production implied by the patient findings. In comprehension, we predict that the left inferior frontal region will be more activated during the processing of sentences in which several word meanings have to be maintained simultaneously prior to their integration than it will be during processing of sentences in which integration is immediate. We also predict that the same region will be more activated when producing phrases that contain many content words than it will be when producing utterances in which the same content appears in different phrases. The left supramarginal gyrus would not be predicted to show this same pattern, given the good comprehension and production observed in some patients with phonological-STM deficits. Preliminarily results appear promising.
Another recent direction is to examine the specific role of the left inferior frontal gyrus in semantic STM. Recently, on the basis of the results of a case study, we have hypothesized that a semantic-STM deficit may not derive from a rapid loss of semantic information, but instead may result from difficulty inhibiting irrelevant semantic information (Hamilton & Martin, 2005). Such a proposal would be consistent with other hypotheses concerning the role of the frontal lobes in inhibitory processing and the consequences for memory and language of a decline in this processing with aging (Zacks & Hasher, 1994). Our case study revealed a specific deficit in inhibition in the verbal but not the nonverbal domain, suggesting that separate brain regions may be involved in inhibiting different kinds of material. Clearly, additional patient and neuroimaging evidence would be needed to support this claim.
Footnotes
Acknowledgements
The research reported here has been supported by a National Institutes of Health grant (DC-00218) to Rice University.