
Abstract
Recent research from cognitive psychology and cognitive neuroscience has suggested that the control mechanisms by which people are able to regulate task performance can be dissociated into evaluative and executive components. One process, implemented in the anterior cingulate cortex of the brain, monitors the amount of conflict that occurs during information processing; another process, implemented in the dorsolateral prefrontal cortex, is involved with maintaining the requirements of the task at hand and with biasing information processing in favor of appropriate responses. In the current article, we review this theory and some of the research that has supported it, including its implication for understanding cognitive disturbances in clinical disorders such as schizophrenia and obsessive-compulsive disorder. We conclude by addressing several interesting possibilities for future research.
Whenever one performs a task, one has to make sure that one selects the relevant information (stimuli, actions) and not get distracted by stimuli or thoughts that are irrelevant to the task. Such distraction might lead to inappropriate actions, such as errors. How the brain manages to do this is the central question in this paper: specifically, how people manage to pay more attention after they have either made an error or almost made an error.
One of the key aspects of cognitive control is how flexible it is. The issue of how people monitor and correct for errors has become a popular topic of inquiry in cognitive research; not only can this issue offer insight into the flexible nature of control and self-monitoring, but understanding the cognitive and neural basis of these functions can possibly shed substantial light on cognitive dysfunction of self-monitoring and control in clinical disorders such as schizophrenia and obsessive-compulsive disorder (OCD).
ATTENTIONAL CONTROL AND THE PREFRONTAL CORTEX
Many phenomena from cognitive and social psychology are thought to depend on automatic processes. Processes can be automatic when they are innate or highly practiced, because they are part of a strongly activated schema, or because they are imposed by a powerful social context or a strong motivational or emotional state, among other reasons.
Of all the stimuli that one encounters, only a limited set is relevant to what one is trying to accomplish; nevertheless, both relevant and irrelevant stimuli compete for access to the response system. Such competition can be difficult to overcome when the processing of irrelevant information is relatively automatic. When the irrelevant information is associated with a response, reliance on automatic processing might facilitate the task if this response is desirable. However, when the irrelevant, automatically processed information is associated with an inappropriate response, the resulting conflict between appropriate and inappropriate responses might be difficult to overcome. In such conditions, errors are likely to occur. Frequently used experimental paradigms to study such effects include interferences tasks. In such tasks, the participant has to make a forced-choice response to a stimulus while simultaneously trying not to be influenced by some other aspect of the stimulus, the irrelevant stimulus dimension. For instance, in the Stroop task, participants have to respond to the color in which a word is presented. When this word is identical to the target color, for instance the word “red” printed in red, color and word are said to be “congruent” and performance is relatively fast and accurate. When the irrelevant word consists of another color, for instance the word “blue” printed in red, color and word are considered to be “incongruent” and performance is slower and less accurate. This is thought to occur because word reading is relatively more automatic than color naming; therefore the word is hard to ignore, and it activates the associated response. During incongruent trials, the responses associated with the color compete with those associated with the word.
When conflicts occur, attentional control is needed to overcome the conflict by selecting the relevant information and suppressing the processing of irrelevant information. Control is often conceptualized as the ability to represent and maintain the task requirements, to support the processing of information relevant to the goals of the current task, and to suppress irrelevant information. Many neuropsychological and neuroimaging studies have shown that an important brain structure that supports this ability is the prefrontal cortex (PFC; e.g., Miller & Cohen, 2001). Presumably, this brain area is able to influence the processing of information in other, more posterior parts of the brain, such that the processing of relevant information is enhanced and the processing of irrelevant information is suppressed (either directly or indirectly). Greater engagement of the PFC should therefore lead to less distraction and more task-appropriate behavior. The PFC is also involved in the control of other forms of higher-order cognition. For instance, many neurophysiological and neuroimaging studies have shown that this region is able to maintain task-relevant information for brief periods of time, which has supported the notion that this region constitutes a neural basis for working memory. Also, the PFC is involved in task preparation and switching between different tasks. Therefore, it is thought to be involved in many aspects of the executive control over our thoughts and actions.
PERFORMANCE MONITORING
The notion that control regulates other types of information processing begs the question, how is control itself regulated? Without a clear answer to this question, the notion of attentional control becomes homuncular, posing the same questions as it is supposed to answer. Therefore, an answer to this question is fundamental to understanding the flexible nature of attentional control.
We and other researchers have argued that the amount of conflict occurring in the information-processing system plays a central role in regulating how much control is exerted and when control is withdrawn. The anterior cingulate cortex (ACC) of the brain, located on the medial surface of the frontal lobe, has (possibly among other functions) the role of detecting situations in which there are conflicts within the information-processing stream and which therefore require attention to be resolved. Following the detection of conflict, the PFC is alerted, and it can then exert control and resolve the conflict (see Fig. 1). Neuroimaging research has shown that the ACC is activated both during error trials and during correct, incongruent trials (e.g., Kerns et al., 2004; van Veen & Carter, 2002).
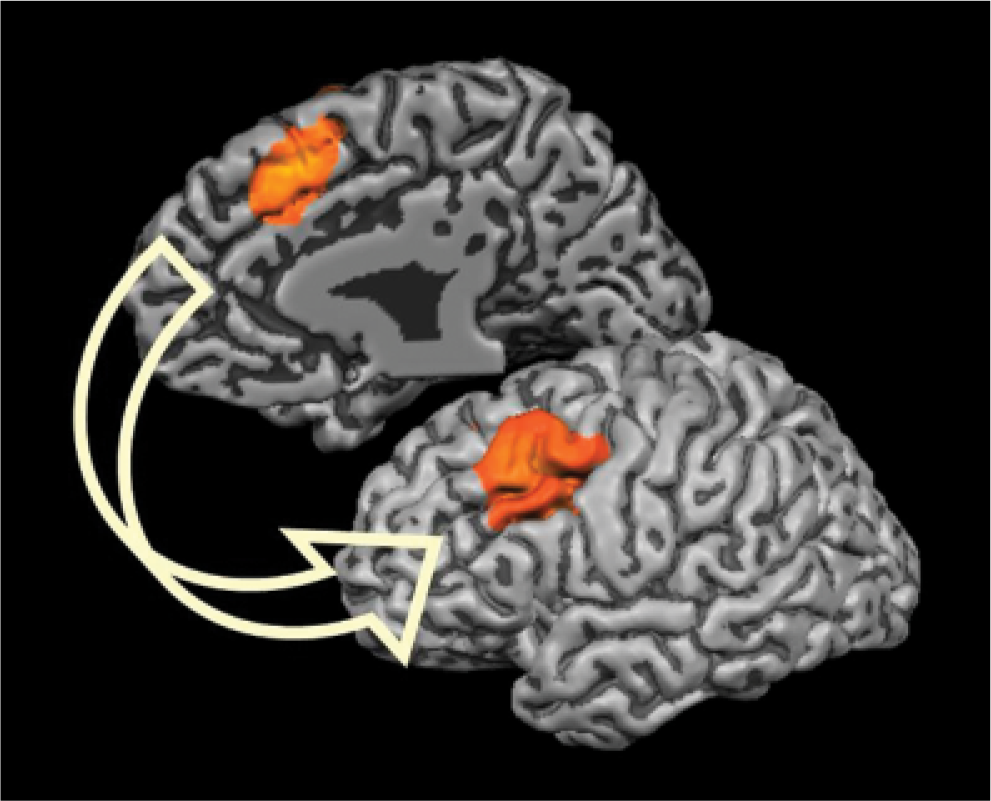
The anterior cingulate cortex (ACC; upper left, on the medial wall of the right hemisphere) and the prefrontal cortex (PFC; lower right, on the lateral side of the left hemisphere). The ACC is proposed to detect the presence of conflict and to alert the PFC to resolve the conflict.
Many labs, including ours, have used interference tasks to study automatic response activation and control engagement. Psychophysiological studies have focused on the timing of the activation of the responses in tasks like these. When the irrelevant stimulus (the word, in the Stroop task) is processed relatively automatically, the associated response is activated early following stimulus onset, whereas the response associated with the relevant (target) stimulus dimension is activated relatively late (e.g., Gratton, Coles, & Donchin, 1992). Thus, in experiments in which participants need to respond to a relevant stimulus with a particular hand and the irrelevant and relevant stimulus dimensions are incongruent (i.e., the relevant response is associated with the opposite hand), the incorrect response is activated first. This activation of the incorrect response is followed by activation of the correct response. If the initial incorrect activation does not manage to reach response threshold and the correct response manages to override the incorrect response, the response to the trial ends up being correct.
Errors in speeded-response tasks are typically fast, impulsive responses based on incomplete stimulus evaluation. The timing of response activation during error trials does not differ much from that during correct trials, the main exception being that during error trials the initial activation of the incorrect response does reach response threshold. The subsequent activation of the correct response is manifested as the tendency to “correct” the error.
Because of the timing of response activations, the timing of ACC activation with respect to the response differs; during correct incongruent trials, both responses are maximally coactivated prior to the response, whereas during error trials, both responses are maximally coactivated briefly following the response. Research on event-related potentials (ERPs; cognition-related electrical brain waves at the scalp surface) has indeed suggested that the maximum ACC activation during correct incongruent trials occurs prior to the response and that during erroneous trials it occurs immediately after the response (van Veen & Carter, 2002).
We have studied how activation of these neural systems relates to cognition and performance in interference tasks. In interference tasks like the Stroop task, the engagement of control following conflict is manifested as a reduced interference effect on trials that follow incongruent trials and a relatively increased interference effect on trials that follow congruent trials (Gratton et al., 1992; Kerns et al., 2004). In other words, there is a smaller effect of the irrelevant stimulus dimension, and less conflict, following an incongruent trial. The conflict detected during incongruent trials engages control, and by this increased control the subsequent irrelevant stimulus is suppressed to a greater extent. This results in less interference during incongruent trials preceded by incongruent trials; but it also results in less facilitation if the subsequent trial is a congruent one. Psychophysiological research has indeed shown that the activation of the incorrect response during incongruent trials on which the participant responds correctly is greater when it is preceded by a congruent trial (e.g., Stürmer, Leuthold, Soetens, Schröter, & Sommer, 2002), supporting the notion that conflict is greater during such trials. Furthermore, this phenomenon continues to be true when controlling for repetition priming effects (e.g., Kerns et al., 2004; Stürmer et al., 2002). Functional magnetic resonance imaging (fMRI) studies have shown that the ACC activation during incongruent trials is greater when such trials are preceded by congruent trials than when preceded by other incongruent trials (e.g., Kerns et al., 2004). Again, this phenomenon continues to be true when controlling for repetition priming.
Likewise, when participants make an error during speeded-response tasks, control is more strongly engaged on the next trial. Participants are typically more cautious on the next trial; their responses are slower and more accurate, and this “post-error slowing” has also repeatedly been associated with control processes. Importantly, we have also shown increased PFC activation on trials following conflict and error trials, and we have shown a correlation between the activation of the ACC on conflict and error trials and of the PFC on the subsequent control trials (Kerns et al., 2004)—supporting the presence of a “conflict-control loop.”
CLINICAL APPLICATIONS
Disturbances in the function of this conflict-control loop have been implied in a number of psychiatric illnesses, including schizophrenia and OCD. Impaired performance of schizophrenia patients during interference tasks has been linked to reduced activation of both the ACC and PFC. A recent study of normal and schizophrenic performance in the Stroop task has shown that the ACC of schizophrenia patients shows impaired activation to both conflict and error trials (Kerns et al., 2005). Importantly, schizophrenia patients also showed corresponding impairments of control adjustments in their behavior; both the post-conflict modulation of the interference effect and post-error slowing were reduced in the schizophrenia patients, compared to healthy control subjects. Consistently, both ERP components associated with conflict and errors and postulated to be generated by the ACC are reduced in schizophrenia (Kopp & Rist, 1999). Conversely, conflict-related and error-related activation of the ACC in OCD patients appears to be increased (Ursu, Jones, Shear, Stenger, & Carter, 2003). These and other results suggest that impairments in conflict monitoring play an important role in the control deficits that have been associated with schizophrenia and OCD.
FUTURE DIRECTIONS
The theory laid out in the present paper needs to be put further to the test, to address all the implications of behavioral, fMRI, and ERP data in cognitive and clinical research. One important direction for future research that has raised some interest is whether engagement of the conflict-control loop is dependent on whether or not conflict occurs consciously and, by extension, whether ACC activation is linked to conscious experience (Mayr, 2004). Many studies, including subliminal-priming studies, have suggested that conflict can occur in the absence of awareness; however, studies have not been conclusive as to whether this conflict also engages the ACC or whether such conflict is associated with a subsequent increase in attention. It would be very interesting to see additional research on the role of motivation and awareness of conflict in modulating ACC function, especially in the context of clinical populations.
Not all evidence reported in the literature has provided supportive evidence for the notion that conflict-related ACC activation leads to engagement of control. Recent data obtained from patients with damage to the ACC have suggested that attentional performance might not differ much between patients and controls (Fellows & Farah, 2005). Future research needs to address whether these data replicate and whether the present theory can account for these data or whether revisions are needed. After all, people with ACC damage might use different strategies, involving different networks, than those without. Similarly, future research needs to address the relationship between the theory discussed in this paper and alternative theories of ACC functioning, such as the view that the ACC's response to conflict and errors is part of a reinforcement-learning system (Holroyd & Coles, 2002); it needs to be addressed which of these theories can best account for the existing data, or whether these theories are complimentary rather than competing notions.
Finally, a possibly important topic for further research—one that we are particularly excited about—is to what extent the conflict-control loop plays a role in different psychological phenomena than the ones described in the present paper. If this loop is such an important part of our cognitive architecture, as we believe it is, than its functioning should be evident in other phenomena than those observed in performance in speeded forced-choice response tasks. Indeed, researchers from various backgrounds have suggested that this loop plays a role in a diverse range of phenomena in cognitive and social psychology. For instance, within the domain of causal reasoning, Fugelsang & Dunbar (2005) have suggested that the ACC and PFC are engaged when new data about causal relations conflict with a plausible theory that is being considered, and requires attention. Greene and colleagues have argued that during moral judgment, in situations when the “emotional” response to a moral dilemma competes with the cognitive, “rational” response, the ACC detects this conflict and engages the PFC to resolve the conflict in favor of the cognitive response (Greene, Nystrom, Engell, Darley, & Cohen, 2004). Anderson et al. (2004) have suggested that during thought suppression, the ACC detects the conflict between memory intrusions and the intention to suppress them and alerts the PFC to actively suppress the unwanted thoughts. Similarly, these areas have also been suggested to play a role in cognitive dissonance; Harmon-Jones (2004) has hypothesized that the ACC detects conflicts (dissonance) between actions and attitudes and alerts the PFC to reduce this dissonance. It would be interesting and informative to see what other areas of research the theory applies to; whether control in these different situations is also regulated by the amount of conflict during information processing; and whether these functions are also carried out by the same brain regions, the ACC and the PFC. As such, investigating whether the theory will generalize to other areas of investigation will also render the theory more falsifiable.