
Abstract
Research on young animals and humans has demonstrated the critical importance of the fetal stage as a formative period in normal development. However, the significance of these findings has not always been incorporated into our thinking when trying to elucidate the origins of health and disease. It is not only that babies react to the state of the mother and to salient environmental events while still in the uterus. This stimulation and priming seems to be essential for guiding the optimal maturation of the nervous, endocrine, and immune systems. Experiences during prenatal life also program the regulatory set points that will govern physiology in adulthood. During this malleable maturational phase, these biological processes should be viewed as flexible “learning systems” that guide the developmental trajectory toward health or derail it toward pathology. Our studies on infant primates have shown that the competence of their immune responses and the structure and activity of certain brain regions, as well as many aspects of behavior and emotional reactivity, are strongly affected by the pregnancy conditions of their mothers.
For much of the last century, the concept of tabula rasa was an influential, albeit controversial, idea in developmental and psychological theory. The notion that an individual starts out as a clean slate at birth is usually credited to the 17th-century philosopher John Locke. Its emphasis on the importance of postnatal experience is certainly sound, as are the ethical implications: that everyone should be assured of the same opportunities in life. However, an objective look at the evidence does not support the conclusion that development begins at birth and that all infants are identical at delivery, emerging from the womb as a blank canvas awaiting the paintbrush. Any parent can attest to the fact that newborn babies vary greatly in size, alertness, and even temperament. Perhaps more significantly, the developmental trajectory toward health or certain illnesses had already been set in motion while the baby was in the womb. This provocative conclusion is encapsulated in a new term, prenatal programming, which derives from epidemiological research on patterns of disease in human populations and from mechanistic studies in animals (Barker, 1995). The findings indicate that many processes governing the regulation of our physiology are encoded during the fetal stage (Wintour et al., 2003). In addition, when pregnancy and fetal development are significantly perturbed, the normal course of maturation may then be shunted in a pathological direction, undermining the original plan for a healthy adult phenotype.
PRENATAL PROGRAMMING
The potential malleability of the immature fetus should be viewed as a type of adaptation and learning from experience. However, this responsiveness and the refinements more typically associated with postnatal life are occurring in utero before the moment of birth. It is not only that the fetus responds to salient environmental stimuli and to the well-being of its mother; the normal development of its brain and other physiological systems appears to be critically dependent on this priming. We know the most about metabolic and growth adjustments in response to poor nutrition. A sustained period of marginal nutrition during pregnancy can change the baby, such that if raised later in an abundant and rich environment it will be more prone to obesity and thereby conferred with an increased risk for diabetes and cardiovascular disease (Murphy, Smith, Giles, & Clifton, 2006).
In many ways, the developing brain and the lymphoid tissues that will become the infant's immune system should be viewed similarly as “learning systems,” even during the fetal period. Many factors, beyond just the genes, influence which neurons will survive and which types of white blood cells will populate the thymus gland. Only a small portion of these immature T lymphocytes are maintained to become the regulators of immunity in the adult. Cells that might react against healthy tissues are deleted by a self-imposed death (apoptosis). Many of the developing neural, endocrine, and immune processes are designed in this way: to be malleable and responsive to contextual cuing and to be pruned back if not working correctly. Within a fairly large, tolerable range, this plasticity is adaptive, guiding and enabling infant development, but it can also be a weak link if the fetus is pushed beyond the normal boundaries for health and the alterations become the basis for later vulnerabilities after birth.
DEVELOPMENTAL ETIOLOGY OF ASTHMA
The significance of taking this perspective can be seen in the evolution of medical thinking about the prenatal origins of a childhood illness such as asthma (Fig. 1). The incidence of asthma is rising dramatically worldwide, now affecting about 8% of children in the United States. Clinicians have come to appreciate that asthma's origins can be found in the fetal stage (Warner, Jones, Jones, & Warner, 2000). A bias toward allergies and asthma may already be manifest in newborns with higher levels of certain antibodies (IgE) and lymphocytes that react in a particular manner. In fact, many infants probably have begun to react to food proteins and plant allergens crossing the placenta even before birth. Postnatal warning signs for asthma are also evident in the propensity of some young children for allergies and atopic skin disorders, such as eczema. These responses and symptoms alert us to the fact that many prenatal factors have now been linked to asthma, including premature birth, number of prior pregnancies, maternal infection and smoking, as well as social and economic disadvantage.
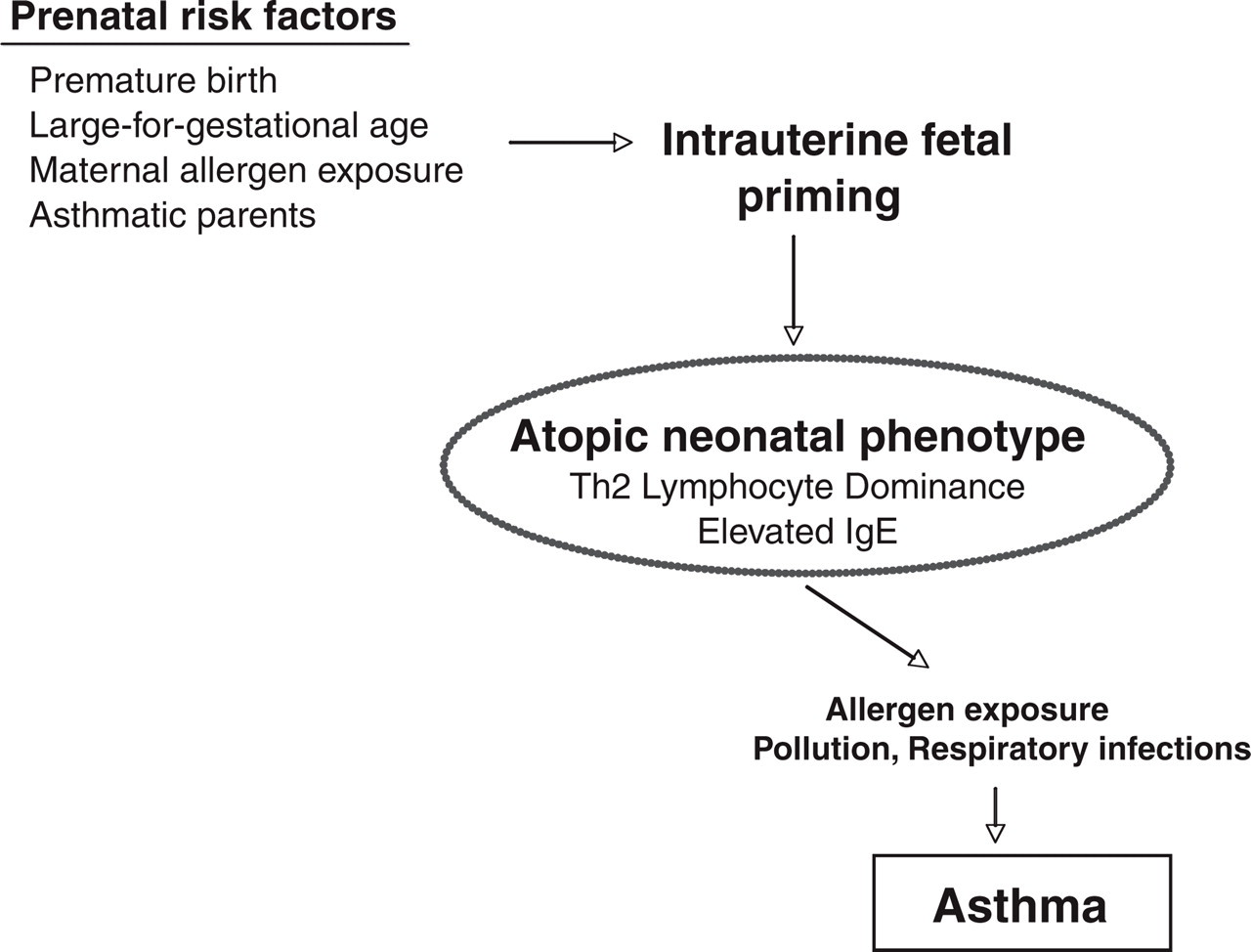
Risk factors for asthma, including those that can be traced back to fetal life. The latter include premature birth and the reactivity of lymphocytes at birth (e.g., a bias of the neonate's cells to secrete IgE and certain types of cytokines when stimulated). Allergens, pollution, and upper respiratory infections during rearing further aggravate an allergy- and asthma proneness already evident in certain young infants.
NONHUMAN PRIMATE MODELS
Apart from the relevance for illness, an appreciation of the subject of “prenatal origins” is critical for gaining insight into other developmental processes. For example, it has been known for many decades that exposure of a fetus to reproductive hormones initiates the sexual differentiation of genital anatomy and brain tissue, upon which cultural factors will later act to sculpt gender differences in postnatal behavior and physiology. Even a genetic female can be transformed to look like an anatomic male by exposure to a sufficient amount of male sex hormone before birth (Berenbaum, 2006). In animals, at least, she will continue to behave in a more masculine manner into adulthood. Over the last 20 years, our laboratory has studied how a wide range of other prenatal conditions affect development in primate infants. We investigated the long-term influence of fetal growth rates, maternal diet, placental transfer of nutrients, and stress. In addition, we assessed the damaging effect of prenatal exposure to too much corticosteroid hormone. We provide a selective review here to demonstrate the potency and persistence of alterations initiated during the fetal stage of development.
One unique feature of our research is that it involved a rhesus monkey colony in existence for over 50 years. Thus, it was possible to review breeding and pedigree records extending back over 5 generations and literally thousands of pregnancies (Price & Coe, 2000). From this information, we extracted familial and environmental influences on birth weight, showing that a fetal growth pattern evident in one pregnancy could have a lingering intergenerational influence on subsequent birth weights in the descendents. For example, a female monkey born at a low birth weight was much more likely to have a small baby herself. Additional analyses of growth patterns between birth and puberty established that the growth rate evident at birth was extremely influential in determining a female's age at menarche and the timing of her first conception (Coe & Shirtcliff, 2004). Thus, slow- or fast-growth trajectories set before birth had many ramifications. As is known to be the case in humans, larger females were likely to reach maturity at a younger age; the larger female monkeys were also likely to conceive sooner than the smaller ones.
PRENATAL ANTECEDENTS OF IRON DEFICIENCY ANEMIA
Many factors affect fetal growth, but certainly maternal diet and the placental transfer of nutrients have a particularly strong influence. We became especially interested in the relationship between maternal iron, infant growth, and the maturation of brain and immune functions. One compelling reason was that iron deficiency is the most common micronutrient problem worldwide, believed to impact over 1 to 2 billion people. Relevant to our hypotheses about the importance of the prenatal period, babies must obtain substantial amounts of iron before birth, because the growing infant is challenged later to obtain enough from breast milk. The solution for mammalian young is to acquire large amounts prenatally through transfer of maternal iron across the placenta. If iron stores are low at birth, both human and monkey babies are at high risk for an iron deficiency anemia (IDA) by 4 to 8 months of age. Many maternal and pregnancy factors predispose for this outcome, including if the mother is young or anemic herself. IDA is also a serious problem in premature babies who may entirely miss their iron allotment. In addition, babies from pregnancies complicated by diabetes and high blood pressure should be monitored closely for a proneness to iron deficiency (Rao & Georgieff, 2002). Our research added another pregnancy factor to consider: maternal stress. Infants born to monkey mothers stressed daily for 6 weeks (or 25% of their 24-week gestation) were destined to become anemic by 4 to 6 months of age, as their growth-related needs exceeded iron stores and dietary intake (Fig. 2; Coe, Lubach, & Shirtcliff, 2007).
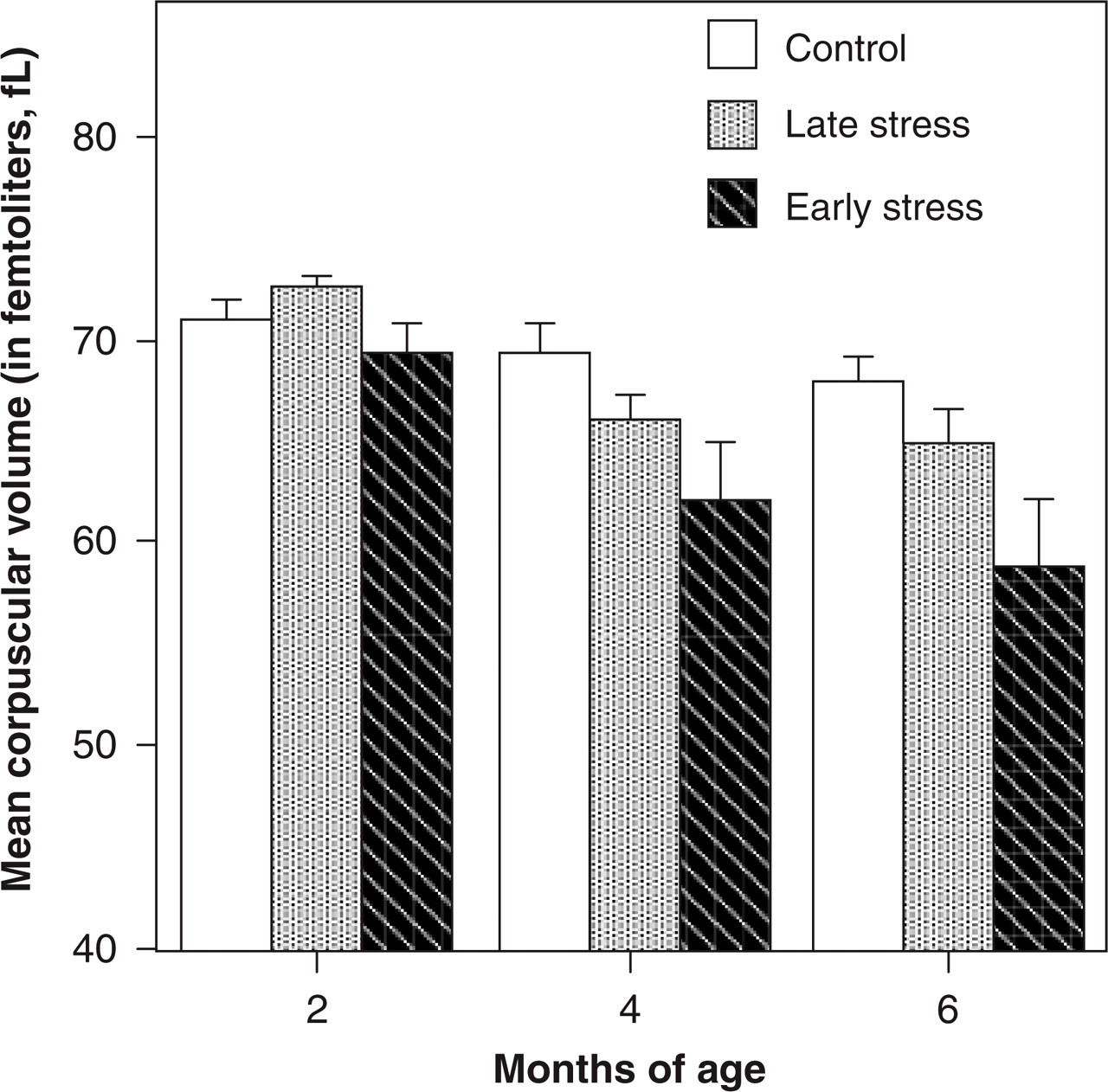
Mean corpuscular volume (MCV) at 2, 4, and 6 months of age for monkeys stressed early or late in gestation and for a control group. Early-stress and late-stress conditions spanned 25% of the 24-week pregnancy and involved a daily arousal induced by 3 acoustical startles during a 10-minute period (days 50–92 and 105–147 after conception, respectively). A greater risk for a low MCV indicative of iron deficiency anemia was one of many effects of prenatal stress, occurring because of lower iron stores at birth. Shown here is the marked decline in the volume of red blood cells as iron was depleted in the growing infant.
The discovery that maternal stress predisposed for infant anemia was important for many reasons. Iron is an essential nutrient needed for normal brain development. It is involved in the synthesis of both neurotransmitters and myelin (the fatty sheath around nerves; Lozoff et al., 2006). Evaluations of anemic animals have documented long-term effects on attentional and cognitive processes; in children, a history of anemia has been shown to influence later school performance. The adverse impact of prenatal stress on iron homeostasis in monkeys added to a long list of other deficits. Infant monkeys from disturbed pregnancies behave differently at birth; they have immature motor reflexes and don't visually orient as well. They remain more anxious and stress-responsive as they develop and, when introduced into small social groups after weaning, become the submissive animals among their peers.
The types of bacteria that reside in the gut were also tracked in undisturbed and prenatally stressed infants (Bailey, Lubach, & Coe, 2004). As infants leave the sterile womb, they have no gut bacteria, but within a day millions begin to colonize the gastrointestinal tract. Prenatally stressed monkeys established lower concentrations of two types of protective bacteria, Lactobacilli and Bifidobacteria, an effect that lasted through 6 months of age. This deficit was associated with a greater susceptibility to Shigella, a type of harmful intestinal bacteria that causes diarrhea. Assessments of the immune responses of prenatally stressed monkeys indicated that their cells also grew differently in culture. Even at 2 years of age, lymphocytes from prenatally stressed monkeys could still be distinguished from those of nonstressed monkeys. When stimulated with lipopolysaccharide, a common protein from bacteria, their cells secreted below-normal levels of proinflammatory cytokines (important for mounting an immune response during the initial stage of infection).
NEURAL IMPACT OF STRESS AND PRENATAL CORTICOSTEROIDS
Other signs of abnormal development after prenatal stress were evident in sensitive regions of the monkeys' brains. The size of the hippocampus was affected (Coe et al., 2003), as has been found in rodents. At 3 years of age, the hippocampi were approximately 10% smaller in prenatally disturbed monkeys. In addition, when the numbers of new neurons were quantified, there was 32% less growth ongoing in the dentate gyrus of the hippocampus. The magnitude was similar to the neural effects observed in infant monkeys exposed to the potent glucocorticoid drug dexamethasone (Dex) after it was administered to their pregnant mothers (Uno et al., 1990). Many studies have been conducted to assess the consequences of prenatal exposure to Dex, because the placental transfer of cortisol, its natural hormone homologue, is often hypothesized to mediate the deleterious effects of stress (Coe & Lubach, 2005). Research on corticosteroid drugs is also of clinical relevance because a 2-day course of Dex or betamethasone is typically given to an expectant woman who may deliver early or to the baby born premature. Thus, these findings from animals are potentially germane to more than 100,000 babies born prematurely each year just in the United States.
Dex certainly does have beneficial effects, serving to accelerate the maturation of lung function and to lessen the likelihood of hemorrhaging in the smaller blood vessels of the brain, but there is reason for some concern about lingering side effects. In animals, Dex jolts many physiological systems and may have a “reprogramming” type of action (Matthews, 2000). The findings suggest it may change the regulatory set points of the hypothalamic-pituitary-adrenal (HPA) axis (Seckl, 1997). Prenatal exposure may result in the maintenance of higher adrenal hormone levels or a propensity for longer-lasting cortisol responses to stressors. When we assessed the lymphocytes of 1-year-old monkeys that had been exposed to Dex before birth, their cells were less sensitive to cortisol feedback. This insensitivity was interpreted as being due to more frequent exposure to higher cortisol levels circulating in their bloodstream.
NEUROBEHAVIORAL CONSEQUENCES
Beyond just the lasting effects on the body, prenatal conditions can influence behavioral and brain functioning (Wadhwa, Sandman, & Garite 2001). Several domains appear particularly sensitive to prenatal challenge and/or abnormal early rearing (van den Hove et al., 2005). Processes underlying attention and emotion regulation are affected in the infant monkey. It is also interesting that manual dexterity—the ability to plan and execute fine motor acts—and spatial navigation also seem quite susceptible to disturbance. This finding is consistent with the common observation that clumsiness and problems with coordinating complex movement patterns are frequently associated with neurodevelopmental disorders. Behavioral deficits of this type suggest neural impairments extending beyond just the hippocampus and limbic structures and likely implicate striatal and cerebellar circuitry. Maturation of neurotransmitter pathways associated with emotionality and motivation—including the norepinephrine, dopamine, and serotonin neurons connecting the midbrain to the forebrain—are also very responsive to early life events. Prenatal and rearing influences on dopaminergic activity may help to explain why some individuals are more prone to using addictive psychoactive drugs that act primarily on this neural substrate.
CONCLUSIONS AND IMPLICATIONS
In sum, we have come to appreciate that the womb does not entirely buffer the fetus from the emotional state and experiences of the mother. Events occurring in the outside world can be transmitted by alterations in placental functioning (Fig. 3). In fact, normal fetal development seems to require some priming and an ability to respond in an adaptive manner to placental and uterine cues. The baby is an active participant in the success and progress of this gestational duet. Even during the first days after conception, as the embryo embeds in the uterus, it starts to interact with and alter the surrounding milieu. Once the placenta is formed, many aspects of maternal and fetal physiology are then coordinated throughout pregnancy. This responsiveness permits more flexibility than would be possible if just following a rigid genetic prescription. However, the same adaptability can become the source of problems under some circumstances. Our research has documented the many types of events that are salient and potent enough to disturb fetal development. Some effects were long lasting and resulted in a different adult phenotype at maturity. It seems reasonable to describe many of these influences as a “reprogramming” that altered the developmental plan. However, from a semantic point of view, it is important to make a distinction between fetal programming and teratology, which is the pathological effect exerted by poisonous drugs and toxic chemicals.
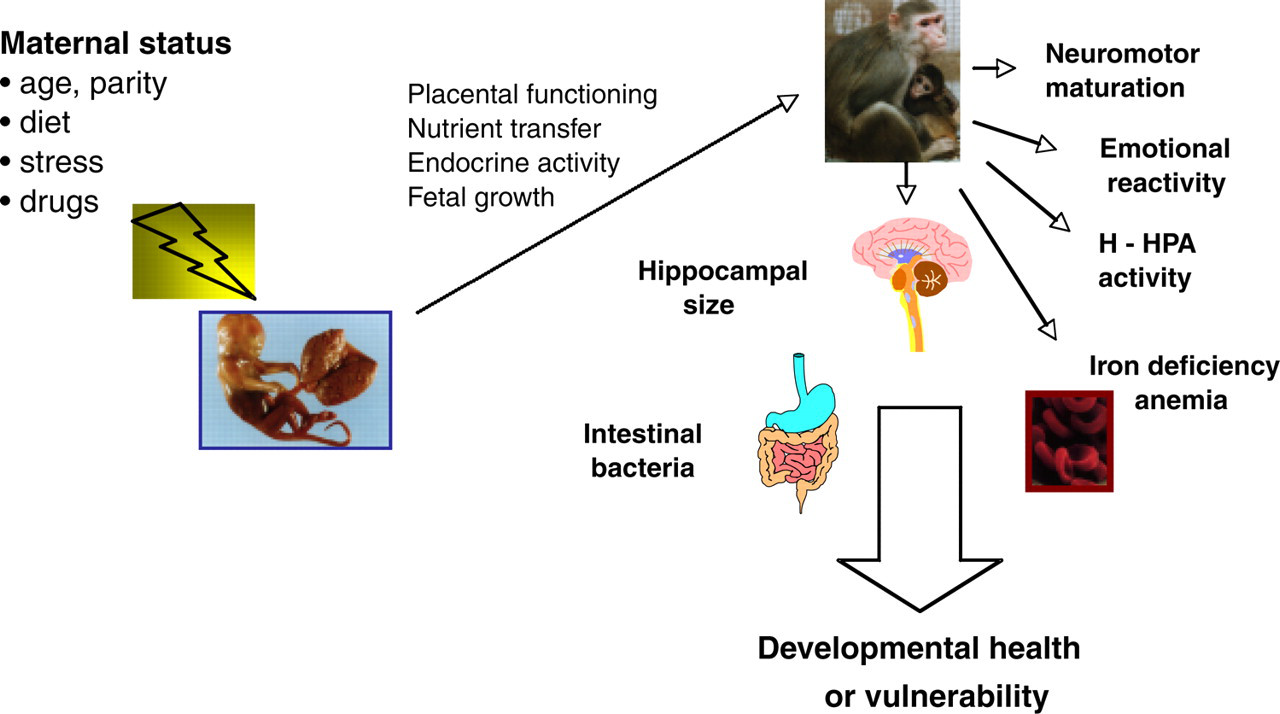
Pathways through which maternal status (diet, stress, etc.) can have lasting effects on the behavior and physiology of infants. The mechanisms and processes include effects on placental transfer of nutrients and hormones, which can impact fetal growth. In this way, prenatal conditions and the health of the mother lay the foundation for resilience or create a vulnerability to illness if the offspring is challenged postnatally.
While the responsiveness of the fetus is easiest to illustrate with examples of abnormality, it should be emphasized that developing babies are remarkably resilient. Certainly, miscarriage, prematurity, and delivery complications can and do occur when challenges to pregnancy become extreme. But in our studies, all of the infant monkeys were viable and would not be readily distinguished from normal animals by a casual observer. Most deviations remained within the normal range and were within the tolerable realm of variation. The vulnerabilities were often unmasked only when the monkey was subjected to a second challenge postnatally. Thus, the developmental ramifications are best described as requiring a second hit or provocation to become manifest (i.e, as a diathesis).
We can return, therefore, to our starting point—the importance of the postnatal world for guiding and ensuring normal development. Societal and economic polices that foster a benevolent rearing environment are still the most enlightened stance, because they serve to maximize the health and behavioral potential of all infants regardless of any differences or deficits evident at birth (Keating & Hertzman, 1999).
FUTURE DIRECTIONS AND COMPELLING QUESTIONS
Many basic-science and translational issues still need to be resolved. More research needs to be conducted on the mediating pathways, beyond just excessive maternal cortisol, including those factors that affect placental functioning. Fetuses change across gestation; it is important to know which systems are most affected early versus late. Impaired infants also react differently to the rearing environment, as exemplified by the fewer protective gut bacteria we found in stressed infants. Understanding variation in responsiveness is germane for designing optimal therapeutic treatments to facilitate recovery in all babies, including those that may react differentially to treatments. While high-quality prenatal care and prevention are the most prudent course for maximizing the likelihood of a healthy baby at delivery, there are many opportunities to investigate creative and compassionate interventions for children with special needs.
Footnotes
Acknowledgements
The primate research was supported by grants from the National Institute of Mental Health, the National Institute of Allergy and Infectious Diseases, and the National Institute of Child Health and Human Development (MH59106, AI067518, HD39386). We gratefully appreciate the help from the staff and students who made these studies possible.