
Abstract
Nondeclarative learning refers to abilities characterized by a lack of awareness of what has been learned and an independence from medial temporal lobe structures that support conscious memories of facts and events. Neuroimaging approaches have been used extensively in two domains of nondeclarative learning—priming and skill learning—to investigate the neural substrates supporting performance. Recent neuroimaging studies have attempted to understand what is being learned in different tasks in order to inform psychological theories of nondeclarative memory. For example, priming may be considered a form of perceptual learning or a form of stimulus–response learning, and correlations between performance and activation patterns in different regions may suggest the nature of the brain changes that support behavior. The attainment of expertise in a skill has been characterized as greater efficiency of processing in the same neural structures that support novice performance or, alternatively, as the recruitment of additional regions. Current research suggests that, within the domains of priming and skill learning, there is much heterogeneity in the underlying brain representations and psychological theories will need to account for these variations.
Several decades ago it was discovered that patients with profound amnesia, who are almost completely unable to learn about new facts and events, are nonetheless able to demonstrate learning in other situations (see Squire & Zola, 1997, for a review). For example, such patients demonstrate normal levels of priming—in which a previous encounter with a stimulus results in improved performance on tasks in which the stimulus is encountered again. Also, these patients can show normal acquisition of skills. Priming and skill learning share the property that subjects are generally not able to verbalize or “declare” what has been learned (e.g., it is difficult to describe in words the skill of riding a bicycle) and are thus considered examples of “nondeclarative” learning. In nondeclarative-learning tasks, subjects are not necessarily aware of what they have learned—or indeed, that learning has even taken place. Subjects can exhibit priming even if they do not remember their initial encounter with the primed stimulus. Findings from amnesic patients indicate that nondeclarative learning does not depend on the brain structures, including the hippocampus, that are damaged in amnesia. Neuropsychological studies initially focused on the capabilities of nondeclarative memory, but neuroimaging studies have recently provided insights into the nature of the neural representations supporting different forms of nondeclarative learning.
Although all forms of nondeclarative learning appear to be distinguished by their independence from structures in the brain's medial temporal lobe, there are substantial differences between them. For example, priming can occur after a single exposure to a stimulus, while skill learning is usually gradual and incremental. Not surprisingly, the neural structures supporting priming and skill learning differ, as do the psychological theories developed to account for these types of learning. In this article we discuss priming and skill learning in turn, describing how recent advances in neuroimaging have informed theories of mechanisms and discussing what issues remain unresolved.
PRIMING
One way to understand priming is to think of it as an increase in fluency associated with repeated stimuli (Jacoby & Dallas, 1981). If a stimulus has been recently processed, it requires fewer cognitive resources to process it when it is encountered again. By this view, a recently encountered word embedded in visual noise is easier to identify than a novel word, on account of the former's greater perceptual fluency.
Initial functional magnetic resonance imaging (fMRI) studies contributed to the idea that priming arises due to alterations in the cognitive representation of a repeated stimulus. These studies showed a decrease in activation in response to repeated stimuli in regions of the brain involved in visual perception, including occipito-temporal regions (see Schacter & Buckner, 1998, for a review). These decreases in activation have been referred to as neural priming, implying that fluency leading to behavioral priming may be the result of reduced neural activity required to effectively process a repeated stimulus. According to one view, neural priming reflects a sharpening of perceptual representations (Wiggs & Martin, 1998). Those neurons that encode critical features of the stimulus for the task at hand (e.g., identifying a stimulus as quickly as possible) would continue to fire across repetitions, while those neurons encoding elements of the stimulus that are not useful would drop out with repetition. It is unclear how the relevant versus irrelevant aspects of the representation would be identified, but the net result would be a decrease in the number of neurons firing. This more efficient neural representation would result in a faster behavioral response and a decrease in activation associated with repetition. By this view, behavioral and neural priming reflect perceptual learning: Experience gives rise to a new perceptual representation.
The case for a perceptual locus of priming is supported by neuroimaging data reporting neural priming in areas involved in perception. However, the link between neural and behavioral priming is speculative. The development of event-related fMRI has given us the ability to tie performance on individual trials to activation levels. This allows us to correlate the amount of behavioral priming with the amount of neural priming in a given region within an individual subject. Recent studies using this approach have not found compelling evidence for correlations between neural priming in visual regions and behavioral priming.
For example, in a study by Dobbins, Schnyer, Verfaellie, and Schacter (2004), subjects initially made decisions about whether pictured objects (e.g., a pencil or a stroller) were bigger than a shoebox. In the next phase, new items along with repeated items were shown, but the subjects now had to decide whether pictured objects were smaller than a shoebox. The researchers found a sharp reduction in priming when subjects were required to switch their response for repeated items. This finding suggests that behavioral priming can be supported by an enhancement of the link between a stimulus and a specific response. While neural priming occurred in perceptual regions during the task, only neural priming in the posterior prefrontal cortex correlated with behavioral priming across the response manipulation. This finding suggests that changes in the prefrontal cortex, and not in visual areas, supported behavioral facilitation. Similar results were obtained by Maccotta & Buckner (2004), who found that activity reductions in prefrontal regions were correlated with behavioral priming when subjects judged whether stimuli were living or nonliving. These correlative findings suggest that, in some cases, priming is more akin to rapid learning of stimulus–response habits than it is to perceptual learning.
The relationship between behavioral and neural priming in prefrontal regions has been reported using other methods, in tasks in which a decision is made about a stimulus with a fixed number of alternatives (e.g., yes or no; Schacter, Wig, & Stevens, 2007). Using magnetoencephalography, Dale et al. (2000) found less activity for repeated words than for novel words in ventromedial temporal and prefrontal areas, and little difference in perceptual regions. In this study, subjects made decisions about whether items were greater or less than 1 foot in length. Furthermore, a causal link between the prefrontal cortex and behavioral priming was demonstrated in a study using transcranial magnetic stimulation (TMS) to disrupt regions showing neural priming in a decision task, as shown by fMRI. The TMS technique uses a rapidly changing magnetic field applied on the surface of the scalp to temporarily disrupt neural activity in a specific cortical region. TMS treatment in prefrontal regions that exhibited neural priming impaired behavioral priming, whereas targeting visual regions associated with neural priming had no effect (Wig, Grafton, Demos, & Kelley, 2005).
In light of this neuroscientific evidence, cognitive theories of priming emphasizing response learning are increasingly plausible. Correlations of trial-by-trial performance with brain-activation levels and TMS findings of causal links between priming and prefrontal activations have to some extent shifted the focus of priming research to anterior regions of the brain and away from the perceptual cortex. However, these neuroimaging results may be the result of the type of priming paradigms that are well-adapted to fMRI research. This includes tasks in which subjects make yes/no decisions about stimuli using a button box. Priming can also occur in tasks in which responses are not so constrained, thus they may not rely on stimulus–response facilitation and instead may depend more on neural priming in perceptual regions. For example, picture naming is facilitated for previously presented pictures, and subjects are likely to complete word stems (e.g. mot___) with previously presented words. Indeed, patients with lateral occipital lobe damage exhibit selective deficits in these forms of priming, suggesting that perceptual processing was disrupted (Keane, Gabrieli, Mapstone, Johnson, & Corkin, 1995). There are likely multiple neural priming mechanisms, and more work is needed to show how changes at different neural levels relate to performance facilitation. In particular, a recent report of priming lasting 17 years raises the question of whether the neural priming mechanisms discovered using neuroimaging could support such enduring behavior (Mitchell, 2006).
SKILL LEARNING
Whereas priming is a rapid form of learning, skill learning is distinguished by being gradual. Real-world skills, such as mastering a golf swing or a chess opening, may take months or years to perfect. The fact that skill learning develops slowly can pose practical problems in laboratory settings, as subjects must consent to multiple sessions practicing skills that have little direct benefit to them. On the other hand, fMRI is well suited to assess changes in patterns of activation over the course of practice. Through comparison of activation patterns during novice performance with those associated with expertise, a clearer picture has emerged of the neural dynamics of skill learning.
Psychological theories of skill learning have emphasized the development of automaticity. With extensive practice, many skills can be performed automatically, in that performance does not noticeably decrease if another task is performed concurrently. For example, extensive practice allows ordinary people to carry on a conversation while walking, whereas only expert dancers could converse as easily while performing a tango. Recent cognitive neuroscience studies have been influenced by the idea that automaticity is an important hallmark of expertise. Although the overall level of performance may not change as skill automaticity develops, changes in the underlying neural representation have been revealed through neuroimaging. Performing a perceptuomotor skill generally activates a wide network of regions, and the development of automaticity is associated with a decrease in activation in this network (Kelly & Garavan, 2005). For example, Wu et al. found that practicing a sequential-finger-tapping task to the level of automaticity resulted in decreased activation in a number of regions, including areas in the parietal and premotor cortices and the cerebellum (Wu, Kansaku, & Hallet, 2004). No areas showing increased activation level with the development of automaticity were found, suggesting that the development of automaticity reflects increased neural efficiency, not a qualitative change. Similar results were obtained by Poldrack et al. (2005), who found decreased activation in the caudate nucleus with the development of automatic performance on a serial-reaction-time task. In this task, subjects press keys corresponding to target locations that appear serially in one of four locations. Skilled performance appears to involve rapidly shifting attention to each successive location and selection of the appropriate motor program to press the corresponding key, whereas in the sequential-finger-tapping task expert performance is associated with smooth execution of an internalized motor program of the appropriate sequence of finger movements. This difference between tasks may be the reason that Poldrack et al. did not see decreased activation in premotor regions with automaticity; any changes in the ability to execute the response in the serial-reaction-time task may have occurred rapidly during the first scanning session.
While these studies compared novice to automatic performance, a more detailed picture of skill learning was obtained by examining activation at three levels of training: during novice performance, after extensive training but before skill learning had become automatic, and after overtraining had led to automatic performance (Puttemans, Wenderoth, & Swinnen, 2005). As seen previously, improved performance was associated with decreased activation in premotor regions both during initial learning and during the development of automaticity. In contrast with other studies, increases in activity with learning were seen in some areas, suggesting that improved performance was associated with recruitment of these regions. For example, activity in the putamen and anterior cerebellum increased with learning, and this increase was maintained as the skill became automatic. The reason for this difference from other studies may be that subjects in Puttemans et al. practiced a bimanual coordination task in which they rotated their wrists at different rates. While selection and execution of motor responses may become more efficient with training, bimanual coordination, which is undoubtedly a major component of real-world skills, may continue to require a high level of neural activity even when performance is automatic (see Oliveria & Ivry, 2008, this issue). While neuroimaging data have generally supported the idea that skill learning reflects increasing efficiency, the particular structures involved in learning even simple motor skills differ depending on the task. It may be interesting to analyze the cognitive components of those skills in which neural regions are recruited as expertise develops.
A pattern of activation that decreases in some structures during learning and increases in others may reflect a switching of the cognitive processes that support performance at different levels of expertise. For example, Floyer-Lea & Matthews reported decreases in cortical activity on a visuomotor control task early in training, with the later development of automaticity associated with increases in activation in subcortical regions including the anterior cerebellum and putamen (Floyer-Lea & Matthews, 2004). In this study, subjects controlled a moving target by exerting force on a pressure plate. The shift from cortical deactivation to subcortical activation may have reflected a shift from performance relying on executive control and hypothesis testing (i.e., about the relationship between force and the target position) to performance relying on procedural knowledge of the relationship. Neuroimaging, coupled with probe tests of the content of learning, could allow the identification of neural markers of the shift between declarative and procedural memory.
The identification of neural markers for declarative versus procedural learning can be used to determine the contributions of declarative and nondeclarative knowledge to cognitive-skill tasks. This may be particularly important, as most cognitive skills are not purely declarative or procedural, with performance influenced by both types of knowledge depending on the circumstances. For example, we (Foerde, Knowlton, & Poldrack, 2006) showed that, in a probabilistic-classification task, subjects appeared to perform based on declarative memory unless they had learned the task while distracted by another concurrent task. Although the overall level of performance was similar under both learning conditions, correlations between performance and fMRI activations differed. If subjects had learned under dual-task conditions, performance was correlated with activity in the striatum, whereas performance on items learned under single-task conditions was correlated with activity in the medial temporal lobe (see Fig. 1). Thus, different neural systems appear to support behavior, depending on the training conditions. Here, neuroimaging results helped resolve contradictory findings about whether probabilistic classification is dependent on the medial temporal lobe or on the striatum. The ability to correlate performance with activation level can probe how subjects are doing a task and can thus provide insights that would not be apparent from looking at performance levels alone.
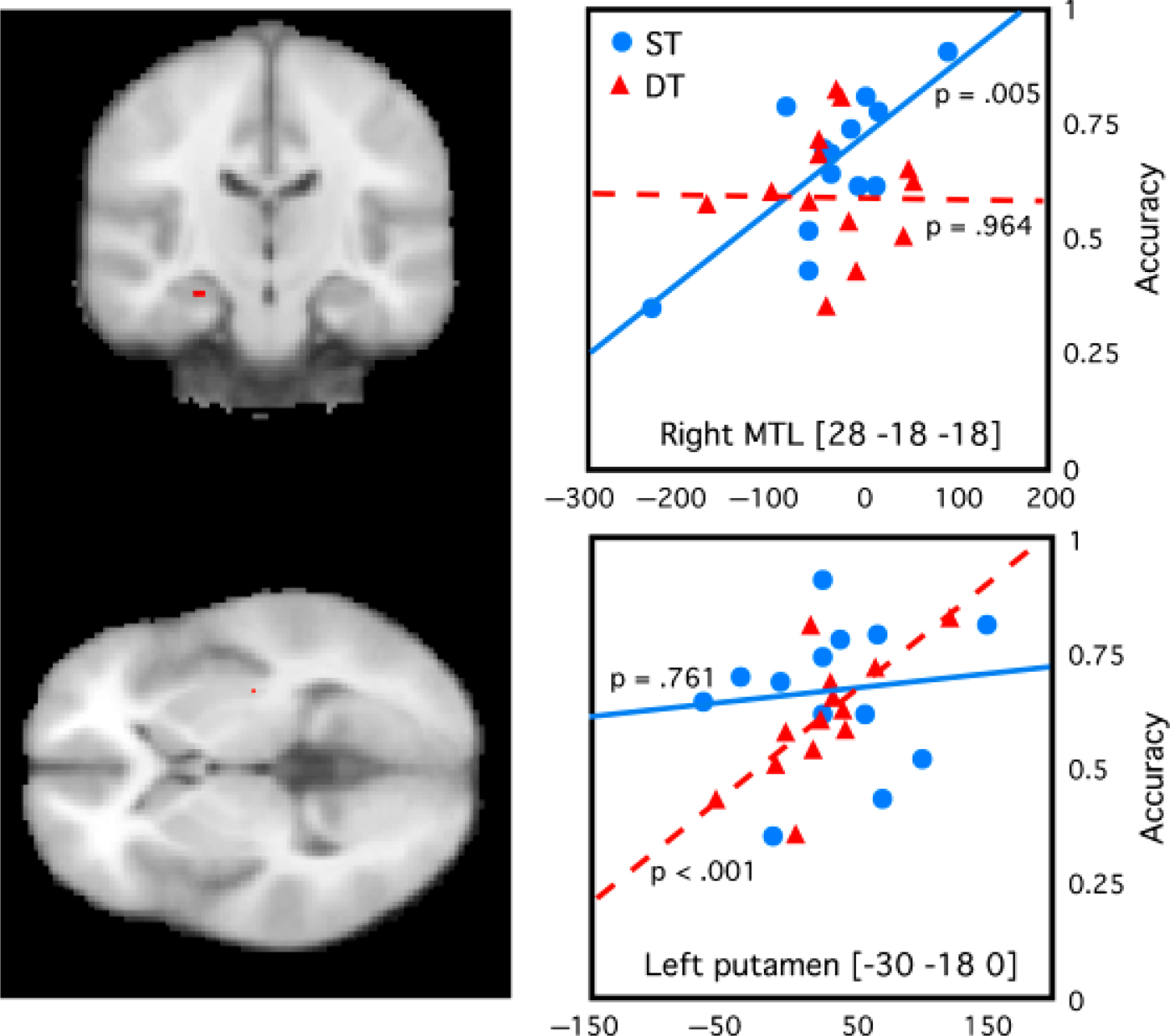
Brain activity associated with a probabilistic-classification task when learned alone (single-task, ST, conditions) or along with another task (dual-task, DT, conditions). Activity in a hippocampal region (activity shown as the red spot in the top left panel) was significantly correlated with performance (accuracy) when the task was learned under ST conditions but not when it was learned under DT conditions (top right panel). Activity in a region in the putamen (activity shown as the red spot in the bottom left panel) was correlated with performance of the task learned under DT conditions but not ST conditions (bottom right panel) The x-axes represent signal change in arbitrary units. Montreal Neurological Institute (MNI) coordinates of the regions of interest are shown on the graphs. (The brain images shown are left–right reversed according to radiological convention.) Data from Foerde, Knowlton, & Poldrack, (2006).
CONCLUSION
Despite constraints in the types of tasks that have been used in neuroimaging studies, there is nevertheless a huge literature on brain activation associated with nondeclarative learning. While these studies have generally supported the predominant view that nondeclarative learning does not rely on medial temporal lobe structures, they have also helped shape theories of how nondeclarative knowledge is represented, by asking whether priming is a perceptual or associational change and whether expertise reflects a quantitative or a qualitative difference in brain activity from the novice level. In addition to informing these theoretical issues, these studies have helped reveal the remarkable breadth and diversity of learning-related changes that occur in the human brain.