
Abstract
Conventional views suggest that exaggerated biological reactivity to stress is a harmful vestige of an evolutionary past in which threats to survival were more prevalent and severe. Recent evidence, however, indicates that effects of high reactivity on behavior and health are bivalent rather than univalent in character, exerting both risk-augmenting and risk-protective effects depending on the context. These observations suggest that heightened stress reactivity may reflect increased biological sensitivity to context, with potential for negative health effects under conditions of adversity and for positive effects under conditions of support. From an evolutionary perspective, the developmental plasticity of the stress-response systems, along with their structured, context-dependent effects, suggests that variation in these systems has been adaptively patterned to increase the capacity of children to match their stress-response profiles to anticipated developmental environments. Taken together, these theoretical perspectives generate a novel hypothesis: that there is a curvilinear, U-shaped relation between early exposures to adversity and the development of stress-reactive profiles, with high-reactivity phenotypes disproportionately emerging within both highly stressful and highly protected early social environments.
Developmental psychologists frequently consider the effects of life experience on development but rarely consider how these effects have been structured by natural selection. Despite this oversight, the burgeoning field of evolutionary-developmental biology has exciting and profound implications for the study of human development (see especially West-Eberhard, 2003). Over the last two decades, theory and research in the field has come to acknowledge that, in most species, single “best” strategies for survival and reproduction are unlikely to evolve. This is because the optimal strategy varies as a function of the physical, economic, and socioemotional parameters of one's specific environment (Crawford & Anderson, 1989), and thus a strategy that promotes success in some environmental contexts may lead to failure in others. Selection pressures therefore tend to favor adaptive phenotypic plasticity, the capacity of a single genotype to support a range of phenotypes in response to particular ecological conditions that recurrently influenced fitness during a species' evolutionary history. Importantly, the development of alternative phenotypes is a nonrandom process; that is, it is the outcome of structured transactions between genes and environments that were shaped by natural selection to increase the capacity and tendency of individuals to track their developmental environments and adjust their phenotypes accordingly.
We have recently proposed a developmental model of adaptive phenotypic plasticity in the human stress-response systems (see Boyce & Ellis, 2005). The model articulates the precepts and rationale for a new claim about the nature of relations between early life experience and stress reactivity, a claim that we have also explored empirically (Ellis, Essex, & Boyce, 2005). We contend that heightened stress reactivity may reflect not simply exaggerated arousal under challenge but, rather, a form of enhanced, neurobiologically mediated sensitivity to context, or biological sensitivity to context (BSC).
The logic of our argument can be summarized in the following way. Biological reactivity to psychological stressors comprises a complex, integrated system of central neural and peripheral neuroendocrine responses designed to prepare the organism for challenge or threat. Developmental experience plays a role, along with heritable variation, in calibrating the response dynamics of this system. Individual differences in such stress reactivity are thought to underlie broad variability in associations between stress and illness and to reflect constitutional variation in susceptibility to stressful challenge. Highly reactive phenotypes, in which affected individuals mount vigorous or sustained autonomic, adrenocortical (cortisol), or other biological responses to stressors, have been viewed as an atavistic health risk factor, a legacy of physiological responses more commensurate with the perils of prehistoric human environments. Often overlooked in such accounts is a body of anomalous observations revealing oppositional, counter-regulatory processes within the stress-response circuitry itself and, even more compellingly, bidirectional effects of reactivity on biomedical and psychiatric outcomes. Highly reactive children sustain disproportionate rates of morbidity when raised in adverse environments but unusually low rates when raised in low-stress, highly supportive settings (Boyce & Ellis, 2005).
Such bidirectional, environment-contingent health effects suggest that BSC is the core, defining feature of highly reactive phenotypes. These observations call into question the presumably unitary pathogenic effects of high reactivity and suggest that its protective effects within specific developmental ecologies might explain the conservation of such phenotypic variation over evolutionary history. BSC reflects sensitivity to both harmful and protective contextual effects. Indeed, the subset of children with highly reactive biological profiles reveals a unique sensitivity or “permeability” to the influence of environmental conditions (Boyce & Ellis, 2005). Further, although a substantial literature documents the capacity of early developmental trauma to predispose individuals toward high biological reactivity, an evolutionary formulation of recent findings suggests a different and novel hypothesis: that the association between early adversity and reactivity is curvilinear in character, with both highly stressful and highly protective environments yielding disproportionate numbers of highly reactive children (Boyce & Ellis, 2005).
THE DANDELION AND THE ORCHID
A Swedish idiomatic expression, maskrosbarn or “dandelion child,” refers to the capacity of some children—not unlike those with low-reactive phenotypes—to survive and even thrive in whatever circumstances they encounter, in much the same way that dandelions seem to prosper irrespective of soil, sun, drought, or rain. Observations of such children have generated, for example, an extensive developmental literature on the phenomenon of resilience, the capacity for positive adaptation despite experiences of significant adversity (e.g., Luthar, 2006; Masten, 2007). A contrasting Swedish neologism, orkidebarn or “orchid child,” might better describe the context-sensitive individual, whose survival and flourishing is intimately tied, like that of the orchid, to the nurturant or neglectful character of the environment. In conditions of neglect, the orchid promptly declines, while in conditions of support and nurture, it is a flower of unusual beauty.
The metaphorical invocation of highly reactive children as orkidebarn is consistent with a growing number of studies revealing that high-reactivity phenotypes under specific environmental conditions may be associated with protective, rather than harmful, effects and generate normative or improved health outcomes. Such bivalent effects of BSC on human and primate morbidities have thematically characterized a series of studies reported by Boyce and colleagues. In examining cardiovascular and immunologic reactivity in two samples of 3- to 5-year-old children, for example, significant interactions with environmental stressors were detected (Fig. 1A) in the prediction of respiratory illness incidence over the ensuing several months (Boyce et al., 1995). Specifically, the noted interactions suggested bidirectional effects of reactivity on illness incidence: Highly reactive children in high-stress families or childcare centers sustained significantly higher rates of respiratory illness than their low-reactive peers did, but equally reactive children in low-stress settings were the healthiest of all children in the samples. By contrast, the respiratory-illness incidence of low-reactivity children was unresponsive to environmental stress levels, showing approximately the same, mid-level illness rates in both low- and high-stress conditions. Similarly significant interactions were found for injury incidence (Boyce, 1996).
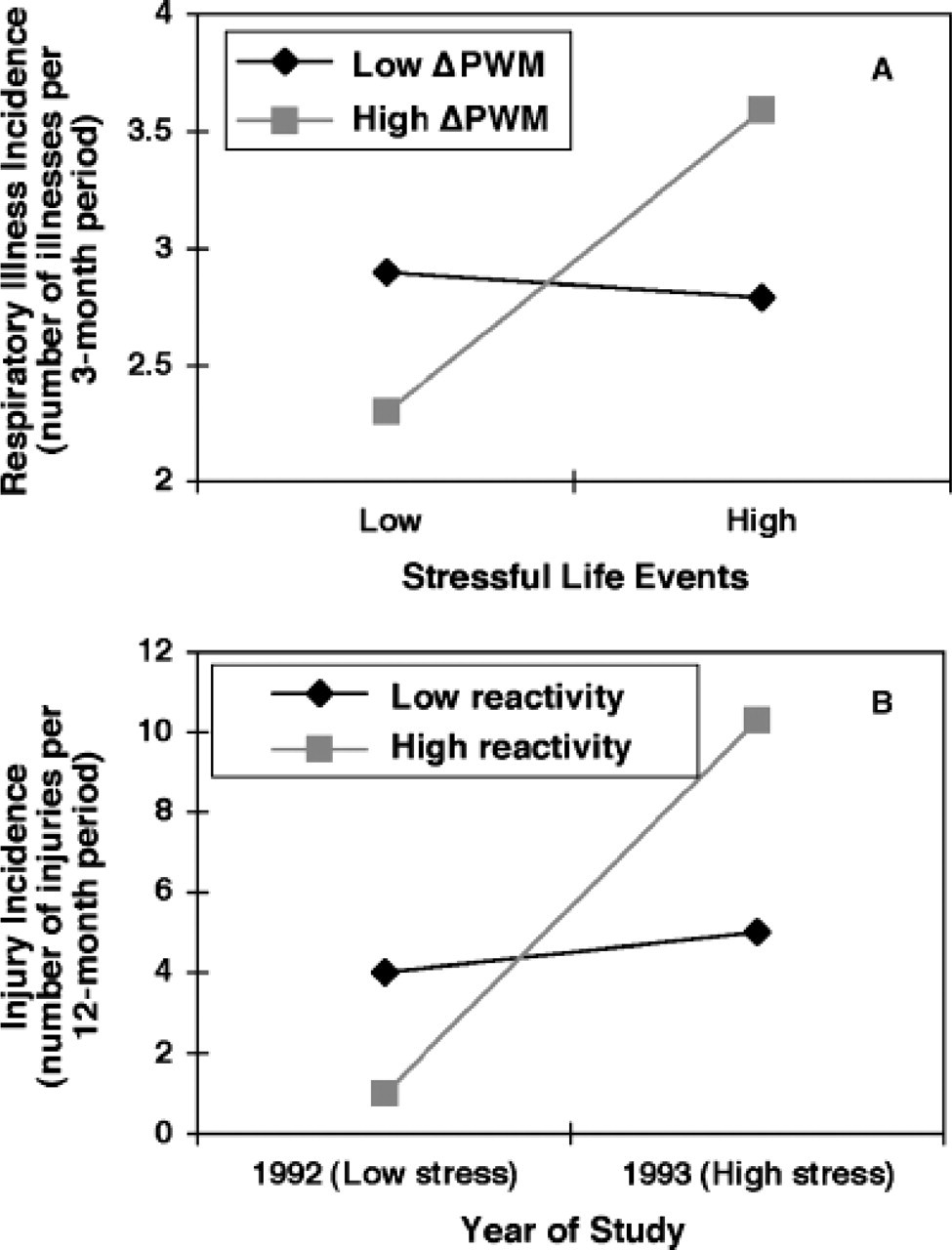
Variation in effects of BSC on physical health across socioecological conditions. The top graph (A) shows a cross-over interaction between immune reactivity (changes in pokeweed mitogen response [ΔPWM]) and family stressful events in prediction of respiratory illness incidence in kindergartners (N = 99; redrawn from Boyce et al., 1995). The bottom graph (B) shows a cross-over interaction between biobehavioral reactivity and confinement stress in prediction of injury rates in a troop of semi-free-ranging rhesus monkeys (N = 36; redrawn from Boyce, O'Neill-Wagner, Price, Haines, & Suomi, 1998).
Even though they were prospective in design, both of these studies were observational in nature and lacked experimental data on the incidence of illnesses or injuries among the same group of highly reactive children in both low- and high-stress conditions. In a subsequent study of semi-free-ranging rhesus macaques, however, such quasiexperimental conditions were satisfied (Boyce, O'Neill-Wagner, Price, Haines, & Suomi, 1998). The troop of macaques, which had been previously assessed for levels of BSC (degree of biobehavioral reactivity to novel or challenging stimuli), lived in a 6-acre wooded habitat in rural Maryland. In 1993, the troop encountered a 6-month period of protective confinement to a small, 1,000-square-foot building, during a construction project on the habitat grounds. The confinement proved highly stressful, and the incidence of violent injuries increased fivefold during the 6-month period. Blinded ascertainment of medically attended injury rates from veterinary records produced evidence for a significant interaction between reactivity status and confinement stress, plotted in Fig. 1B. As with the prior studies of children, low-reactivity individuals showed little effect of the confinement, while those with high reactivity showed dramatically higher rates of violent injuries in the high-stress situation but lower rates in the low-stress condition.
These findings complement research on the bidirectional, context-dependent effects of high intelligence (or high ego development): Whereas highly intelligent, introspective people tend to flourish under relatively benign life conditions, they also tend to be more reactive than others to distress (see Luthar, 2006). Thus, just as high reactivity to stress has generally been considered pathogenic but can be protective in supportive environmental contexts, high intelligence/introspection has generally been thought of as beneficial, but can be harmful if surrounding forces are negative.
DEVELOPMENT OF BSC: AN EVOLUTIONARY APPROACH
Adaptive phenotypic plasticity enables children to match their biological and behavioral systems to the parameters of their early (and predicted future) developmental environments. Given past evidence that early trauma can increase stress reactivity and new evidence that high reactivity can be protective in highly supportive settings, we (Boyce & Ellis, 2005) postulated a curvilinear, U-shaped relation (shown in Fig. 2) between levels of early adversity and the magnitude of biological response dispositions. Specifically, we hypothesized (a) that exposure to acutely stressful childhood environments up-regulates stress reactivity, increasing the capacity and tendency of individuals to detect and respond to environmental dangers and threats; and (b) that exposure to exceptionally supportive childhood environments also up-regulates stress reactivity, increasing susceptibility to social resources and ambient support. Both of these proposed functions of BSC converge with theory and data indicating that temporary, moderate increases in stress hormones and associated neurotransmitters enhance mental activities in localized domains, focus attention, and prime memory storage and thus improve cognitive processes for dealing with environmental opportunities and threats (Flinn, 2006). By contrast, and typically for the large majority of children, (c) exposure to childhood environments that are not extreme in either direction down-regulates stress reactivity, buffering individuals against the chronic stressors encountered in a world that is neither highly threatening nor consistently safe. Although the cellular mechanisms that calibrate such response dispositions are currently unknown in humans, recent work in animal models suggests that epigenetic modifications resulting in differential gene expression may well play an important role (Meaney, Szyf, & Seckl, 2007).
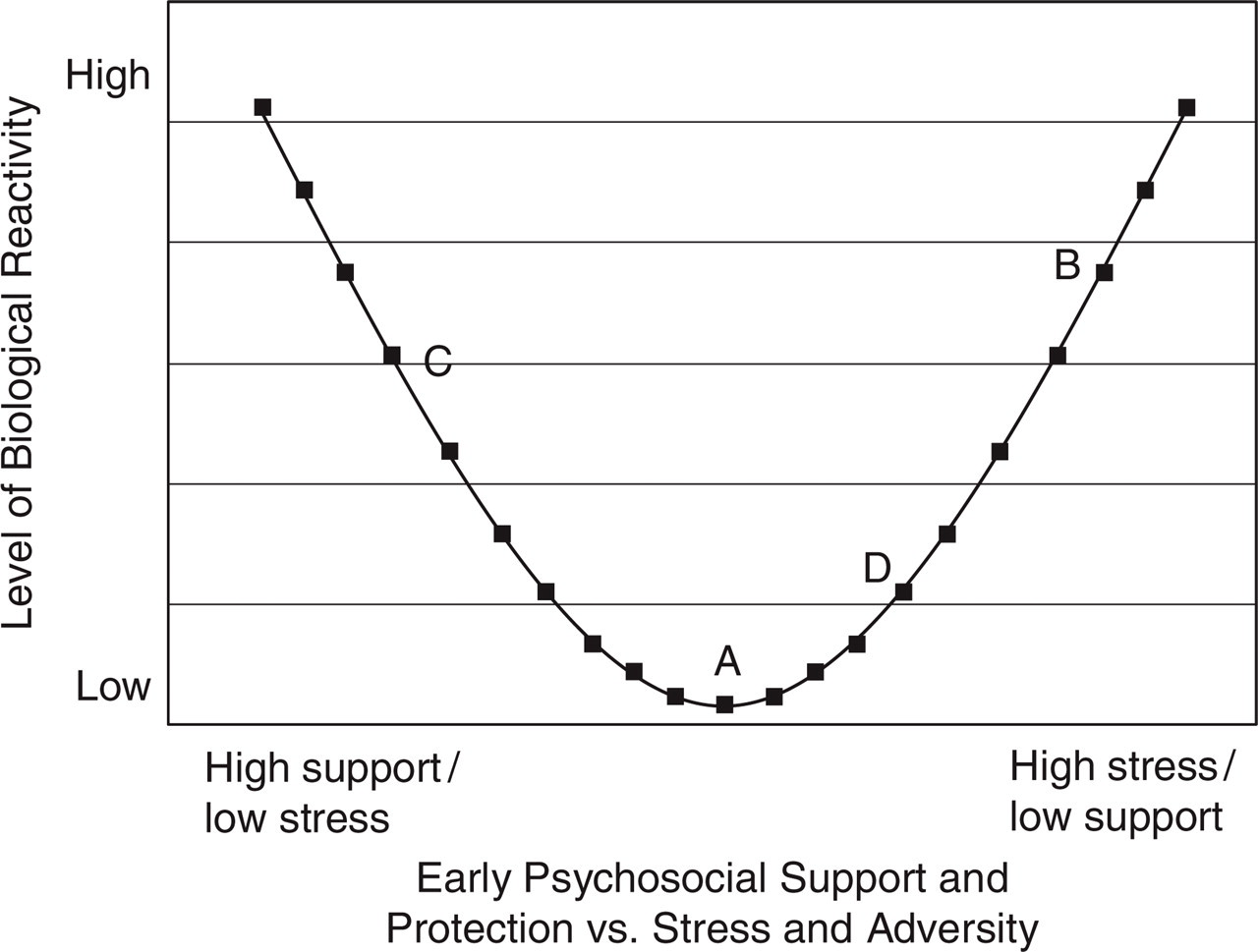
Hypothesized curvilinear relation between biologic reactivity to stress and experiences of support and protection versus stress and adversity in early environments. Comparisons of subjects at points A and B would lead to the conclusion that low support/high stress results in heightened stress reactivity. Comparisons at points C and D, on the other hand, would generate the inference that low support/high stress produces diminished reactivity (adapted from Boyce & Ellis, 2005).
Although the theory predicts up-regulation of stress-response systems in both highly supportive and stressful environments (the U-shaped curve), high stress reactivity may translate into different behavioral phenotypes in supportive versus stressful contexts. Reactive, sensitive children have been found to be more reflective and perhaps more conscious of self and environment; to be more able to delay gratification in pursuit of goals; and to perform better on neuropsychological measures of inhibitory control, executive function, and self-regulation (e.g., Aron & Aron, 1997; Blair, Granger, & Razza, 2005). Up-regulated stress-response systems in children may therefore interact with protective, beneficial developmental environments to produce relatively high levels of cognitive and social competence. Conversely, interactions between high stress reactivity and risky, threatening developmental environments may result in lower thresholds for anticipating threat in ambiguous or unfamiliar situations (e.g., elevated sensitivity to threat cues such as angry faces) and support greater vigilance and wariness in children.
Although the U-shaped curve depicted in Figure 2 specifies environmental sources of variation in BSC, genetic sources of variation and gene–environment interactions are also important and need to be addressed in a comprehensive theory of BSC (see Ellis, Jackson, & Boyce, 2006). Reaction norms are genetically based “bookends” that constrain the range of phenotypes that can develop within varying environmental contexts. Importantly, children differ in the location of these bookends along phenotypic dimensions. For example, children whose reaction norms are located on the upper end of the BSC spectrum have higher starting points for stress reactivity than do children whose reaction norms are on the lower end. These differences can generally be expected to maintain variation in BSC, even if children have equivalent life experiences. In addition, children differ in how widely their bookends are placed. That is, some children have broad reaction norms and display high levels of plasticity in response to developmental experience, whereas others have more narrow reaction norms and display more fixed developmental trajectories (see Belsky, 2005). The current theory should be more successful in accounting for developmental variation in BSC among children with wider reaction norms.
EMPIRICAL EXPLORATIONS OF BSC THEORY
We have initially investigated our curvilinear, U-shaped model of the development of BSC in two studies comprising 249 children and their families (Ellis et al., 2005). In the first study, 3- to 5-year-old children were concurrently assessed on levels of support and adversity in home and preschool environments and on cardiovascular reactivity to laboratory challenges. Because the early environments of these children ranged from exceptionally stable and supportive to moderately stressful, the sample only provided a window into a portion of the proposed U-shaped association between support/adversity and BSC (i.e., the left half of the curve shown in Fig. 2). Within this range, the theory posits that higher stress will be associated with reduced biological responsivity to stressors. In the second study, children were prospectively assessed on family stress in both infancy and preschool and on autonomic and adrenocortical reactivity to laboratory challenges at age 7. This second study sampled the broad range of variation in early childhood environments that is needed to fully explore the curvilinearity hypothesis. Within this range, both highly protected and highly stressful environments should promote heightened BSC.
We found in both studies that a disproportionate number of children in supportive, low-stress environments displayed high autonomic reactivity. Conversely, in the second study a relatively high proportion of children in very stressful environments showed evidence of heightened sympathetic and adrenocortical reactivity. Consistent with our evolutionary-developmental theory, these exploratory analyses also suggested that relations between levels of childhood support/adversity and the magnitude of stress reactivity are curvilinear, with children from moderately stressful environments displaying the lowest reactivity levels in both studies.
CONCLUSIONS AND FUTURE DIRECTIONS
The proposed U-shaped curve can potentially reconcile important contradictions in the existing literature on the origins and consequences of stress reactivity in children. Investigators comparing individuals from points A and B in Figure 2, for example, would conclude, as many researchers in this area have (e.g., De Bellis et al., 1999), that experiences of family and environmental stress are associated with up-regulatory calibrations in biological reactivity systems. Yet studies comparing individuals from points C and D would find, as those reviewed by Gunnar and Vazquez (2001) have, that early stressors are rather associated with down-regulatory changes in salient biological responses. The current theory, which posits two oppositionally distinctive ontogenies for BSC, explains both of these regulatory effects.
A guiding assumption of our work on stress reactivity is that developmental mechanisms have been organized by natural selection to produce enhanced BSC when it is advantageous to the developing person—in both acutely stressful and exceptionally supportive childhood environments. In shaping intervention strategies to prevent developmental psychopathology and other early morbidities, we may do well to consider this conceptualization of individual differences in stress reactivity. Under some circumstances, highly sensitive children may be usefully targeted for interventions involving the provision of ancillary supportive services, while in other circumstances, the needs of highly sensitive children might define the minimum standards of provision for an entire population of children. In still other settings, ascertainment of reactivity status might simply prevent the adoption of a “one size fits all” approach, facilitating the design of strategies and policies specifically tailored to the needs of children with different biological response phenotypes. Whatever the future direct utility of the theory and research described here, it is our hope that this work will advance collective understanding of the sources of individual differences and their implications for the rearing and well-being of children.