
Abstract
It has long been assumed that metacognition—thinking about one's own thoughts—is a uniquely human ability. Yet a decade of research suggests that, like humans, other animals can differentiate between what they know and what they do not know. They opt out of difficult trials; they avoid tests they are unlikely to answer correctly; and they make riskier “bets” when their memories are accurate than they do when their memories are inaccurate. These feats are simultaneously impressive and, by human standards, somewhat limited; new evidence suggests, however, that animals can generalize metacognitive judgments to new contexts and seek more information when they are unsure. Metacognition is intriguing, in part, because of parallels with self-reflection and conscious awareness. Consciousness appears to be consistent with, but not required by, the abilities animals have demonstrated thus far.
Anyone who has tried to remember the name of a long-forgotten friend or well-known public figure is probably familiar with the “tip of the tongue” experience—that frustrating feeling that, even without recalling a word or name, you know that you know it (Schwartz, 2002). Knowing that you know involves metacognition, the ability to think about one's own thoughts, make judgments about one's own memories, and generally (as the name suggests) engage in cognitive processing about one's own cognition.
Metacognition is omnipresent in daily life, though unlike the tip-of-the-tongue experience, it often goes unnoticed. It can be as simple as conveying a feeling of uncertainty. Upon being asked when the last great dinosaur extinction occurred, for example, you might say “65 million years ago” to signal high confidence, “I think about 65 million years ago” to signal some uncertainty, or “I don't know” to signal complete uncertainty. Another example is deciding when to give up on puzzling over an elusive memory. As these examples reflect, most research on human metacognition involves judgments about one's memory (see Dunlosky & Bjork, 2008).
There are two basic categories of metacognition: monitoring and control (Nelson & Narens, 1994). Monitoring involves making judgments about one's memory and/or cognition; control involves using metacognitive judgments to guide one's future behavior. If, for example, one is asked who composed “Rhapsody in Blue,” answering, “I'm not sure” is an example of monitoring; looking up the answer (Gershwin) is an example of control. Metacognition is of special importance when students make decisions about how to study (Kornell & Bjork, 2007); for example, when studying for a test, a student needs to gauge how well he or she has learned a chapter—an example of monitoring—and then decide whether to review the chapter again or turn to something else—an example of control.
METACOGNITIVE MONITORING IN ANIMALS
An animal's life is full of uncertainty—about the source of its next meal, the location of predators, and so on. Watching a bear hesitate before attempting to climb a tree, or a cat waver before jumping a large gap, creates the impression that the animals do not know if they can make it. Not knowing is not a sign of metacognition, however. The question is, does the animal know that it does not know?
The ability to think about one's own thoughts has long been considered a uniquely human ability (Metcalfe & Shimamura, 1994). That assumption was first tested a dozen years ago (see reviews by Smith, Shields, & Washburn, 2003; Smith & Washburn, 2005): Rhesus monkeys (Macaca mulatta) and dolphins (Tursiops truncatus) were presented with a stimulus-discrimination task (e.g., categorizing tones based on frequency) consisting of easy and difficult trials. In addition to standard response options signifying high and low frequency, the animals could also escape (i.e., skip) any trial, thereby forgoing reward but avoiding punishment. Both dolphins and monkeys tended to escape difficult trials, on which they apparently knew they did not know the answer. (Pigeons have not demonstrated metacognition in a similar situation; e.g., Sole, Shettleworth, & Bennett, 2003.)
Categorizing tones, and the ability to make other judgments based on external stimuli, are susceptible to a more mundane explanation than uncertainty monitoring. It might be called the “third response” problem: For example, if an animal learns to respond to the left key for low tones and the right key for high tones, it may learn to respond to the middle (escape) key on medium (difficult) tones—based on the external stimulus (the tone), not an internal cognition (i.e., without metacognition; but for evidence that such judgments are not based purely on external stimuli, see Smith, Beran, Redford, & Washburn, 2006). Metamemory judgments—that is, judgments about internal memory representations instead of external stimuli—are the strongest test of metacognition because they avoid this problem. Rhesus monkeys can escape difficult trials in a metamemory task (see Smith et al., 2003).
Another way to study animal metacognition is to present a stimulus and then take it away, so that it is only available in memory, and then allow the animal to decide whether or not to take a test on that stimulus. Hampton (2001) showed rhesus monkeys a sample picture and then, after a delay, tested their ability to select the sample picture when it was presented among three distractors. On some trials, after viewing the initial picture, the monkeys could choose not to take the test before it began. Memory accuracy was better on self-selected test trials than on mandatory test trials, which implies that the monkeys opted out when they knew they did not know the answer.
PUT YOUR MONEY WHERE YOUR MEMORY IS
My collaborators and I recently measured monkeys' confidence in a new way, by asking them, essentially, to gamble on their memories (Kornell, Son, & Terrace, 2007). After an initial training period (see next paragraph), two male rhesus monkeys were trained in a task in which they were shown six sample pictures sequentially on a touch-sensitive video monitor. After the last sample had been presented, nine pictures were presented simultaneously, one of which had been presented among the previous samples (Fig. 1). The monkeys' task was to touch the sample. Then, before feedback was provided, two “risk” icons were presented, which essentially allowed the monkeys to bet on the response they had just made. A high-risk bet resulted in the gain of three tokens—which could be exchanged for food—if the monkey had selected the correct picture, but a loss of three tokens otherwise. A low-risk bet resulted in a sure gain of one token. We reasoned that a metacognitive monkey would make a high-risk bet when he felt confident about his memory but avoid risk when he was unsure—and that is what happened. The monkeys chose high risk more often on correct trials than they did on incorrect trials.
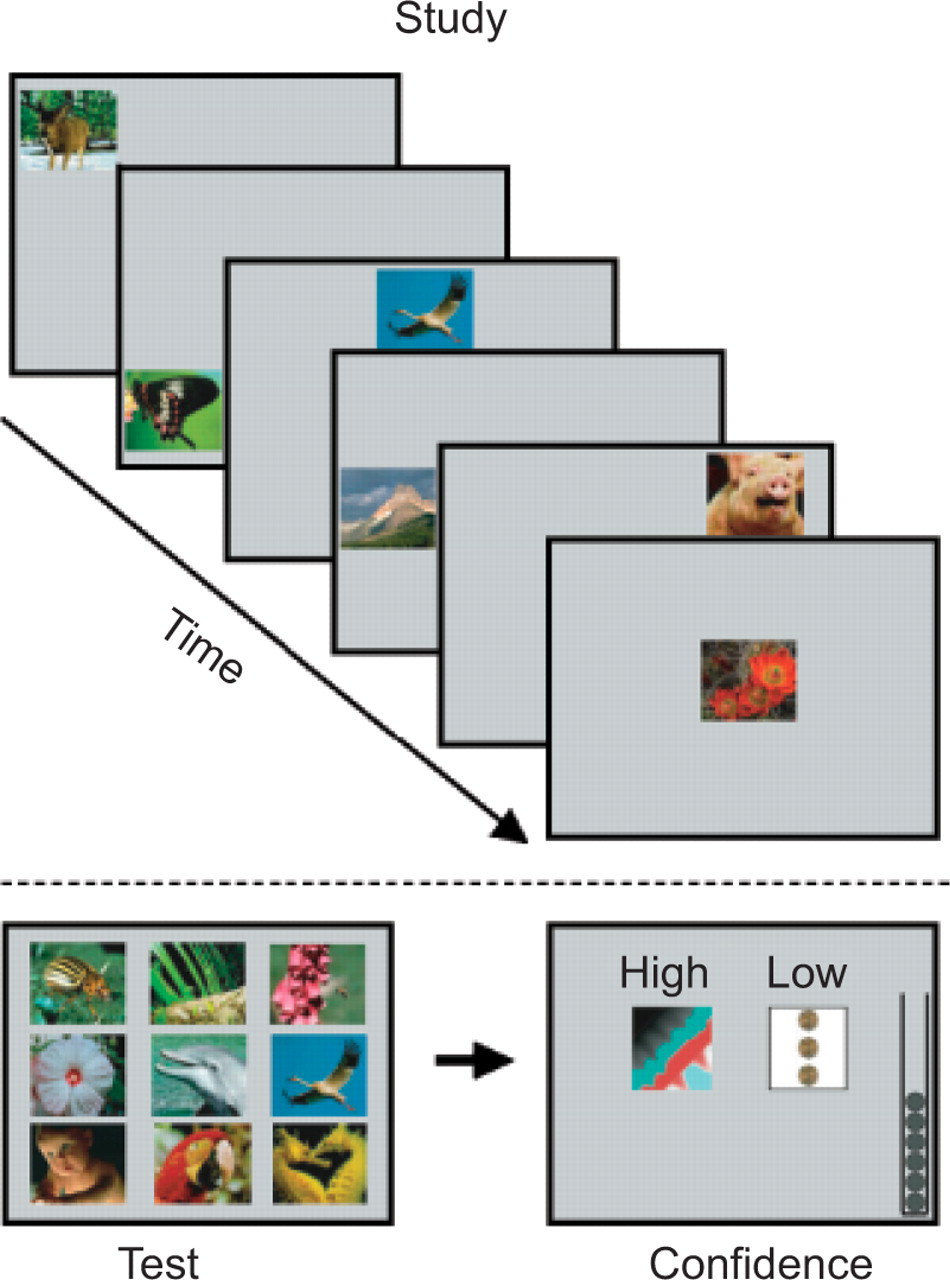
Trial structure of the Kornell, Son, and Terrace (2007) metacognitive monitoring experiment. Six sample pictures were displayed successively, followed by a test during which one of the six samples was presented with eight distractors. The subject's task was to select the sample and then select the appropriate confidence icon. (Figure reprinted from Kornell, Son, & Terrace, 2007, p. 66.)
Our monkeys required extensive initial training to respond metacognitively, whereas humans rarely require any training (at least in the lab, although human children naturally receive extensive metacognitive training in the form of questions like “Are you sure?” and “Do you remember?”). To investigate the necessity of training, the memory task described above (Kornell et al., 2007) was only introduced after we had trained the monkeys to make metacognitive judgments in three discrimination tasks (in which they had to discriminate line length, number of items, and circle size). After this preliminary training, the monkeys were able to respond metacognitively on the first day of the meta-memory task described above. In doing so, they demonstrated a flexible metacognitive ability that was readily generalized to new situations. This flexibility represents a significant advance in the level of metacognitive sophistication animals have expressed.
METACOGNITIVE CONTROL IN ANIMALS
Metacognitive control in humans—that is, using metacognition to control one's behavior—is a theoretically and practically important topic that has been well researched in humans (e.g., Kornell & Bjork, 2007) but not in animals. We investigated metacognitive control by allowing two male rhesus monkeys to request information when they were unsure, just as a person might ask for a hint when solving a riddle (Kornell et al., 2007). The monkeys' job was to learn to press four pictures in a certain fixed arbitrary order (Fig. 2). On some trials the monkeys could press an icon on the right side of the touch screen that produced a “hint”—that is, a blinking border that appeared around the next correct response in the sequence. The price the subjects paid for requesting one or more hints was that correctly completed sequences were rewarded with a banana-flavored pellet, instead of a more appetizing M&M candy. The monkeys completed four sessions on a given set of pictures. Across the four sessions, the monkeys' hint requests steadily decreased as their performance without hints improved. By asking for hints only when they knew they did not know, the monkeys demonstrated metacognitive control. This advance is significant because when the monkeys felt uncertain, they did not simply make an uncertain response, as in previous studies; instead they took steps to rectify their ignorance.
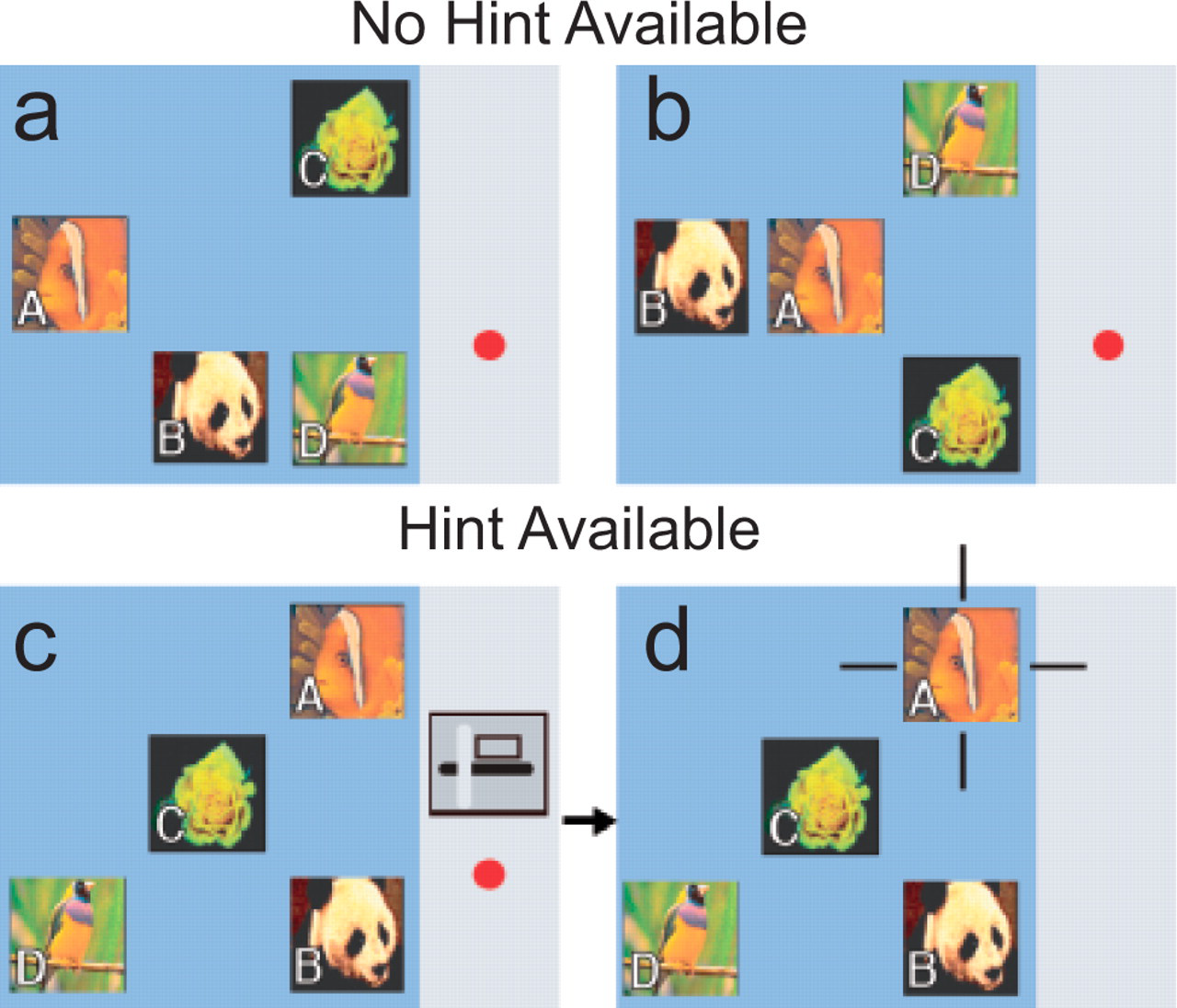
Examples of displays used in Kornell, Son, and Terrace's (2007) metacognitive control experiment. Subjects were rewarded for responding to all items in a particular order (A→B→C→D). As illustrated in (a) and (b), the position of the items varied randomly from trial to trial. On 50% of trials, no hint was available (a and b). On the other 50% of trials, a hint was available, as indicated by an icon that appeared on the right side of the monitor, above a red circle (c and d). Touching the hint icon resulted in the appearance of four blinking lines around the correct item for that position in the sequence. The red circle signaled that the subject could earn an M&M, instead of a less desirable banana pellet, by completing the trial correctly. The first request for a hint eliminated the red circle (d). (Figure reprinted from Kornell, Son, & Terrace, 2007, p. 68.)
Another line of research related to information seeking shows that a chimpanzee (Pan troglodytes), orangutan (Pongo pygmaeus), or 2.5-year-old child allowed to see food being hidden will reach for it directly, but if the hiding occurs out of sight, he or she will do a visual search for the food before reaching into one of the hiding places (Call & Carpenter, 2001). Rhesus monkeys do the same, although unlike apes, monkeys require considerable training (Hampton, Zivin, & Murray, 2004). These results suggest that the need for training may be an important metacognitive difference between apes and other primates (although research on species differences is in short supply). Interestingly, chimpanzees and orangutans also diverge from other primates in mirror self-recognition, a task that, like metacognition, seems related to self-awareness (Gallup, 1982). (There is also some evidence that, unlike rhesus monkeys, orangutans can also escape difficult trials without training; Suda-King, 2008.)
ACTIVE LEARNING AND “DESIRABLE DIFFICULTIES”
Like other primates, humans are fond of hints. The downside to hints, however, is that they can make learners passive. Learning is most effective when learners are active (e.g., solving problems on their own). The hint paradigm described above (Kornell et al., 2007) provided an opportunity to examine the benefits of active learning in monkeys. We (Kornell & Terrace, 2007) trained two rhesus monkeys to touch a set of pictures in a fixed order, as described above. Hints were provided during training for some picture sets (in three hint conditions) but not for others (the no-hint condition). After 3 days of training (either with or without hints), we measured learning on the 4th day by testing the list without hints. Not surprisingly, hints enhanced performance during the first 3 days of training. On day four, however, the pattern reversed: Accuracy in the no-hint condition remained high, while accuracy in the hint conditions dropped to the baseline level that the monkeys had obtained without previous training (Fig. 3). Thus monkeys benefited from being actively engaged in learning. The data fit a classic “desirable difficulty” pattern: The more difficult condition, which impaired performance during training, enhanced long-term learning (Bjork, 1994). People are frequently fooled by desirable difficulties, assuming that activities that enhance training performance also enhance learning; whether other primates suffer from similar metacognitive illusions is unknown.
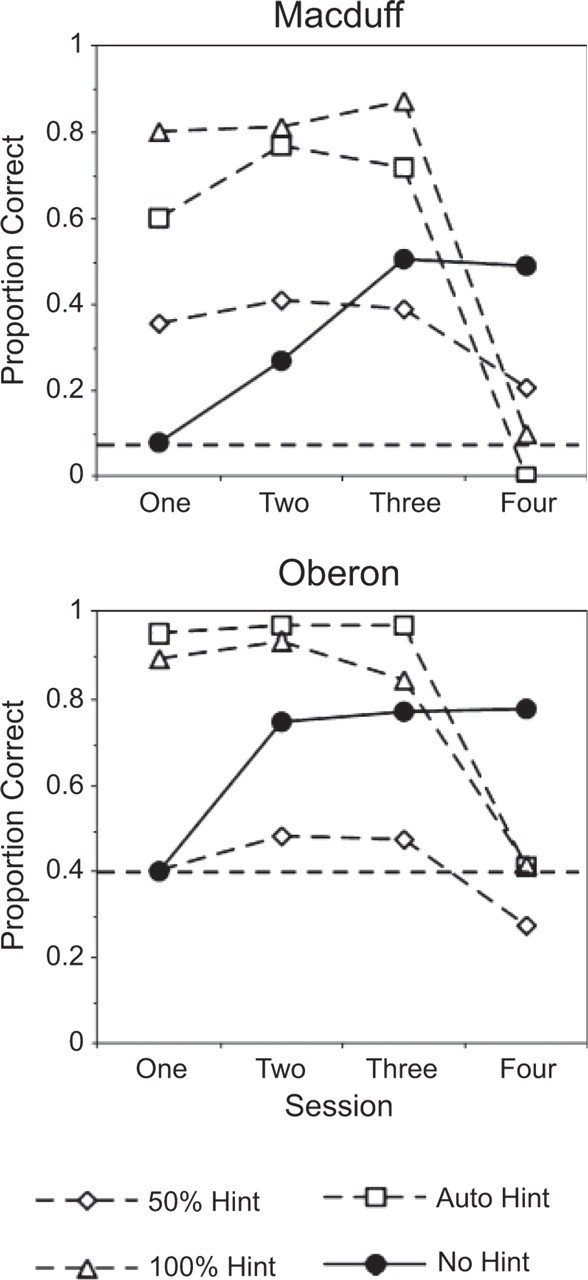
Proportion of trials completed correctly in Kornell and Terrace's (2007) active learning experiment. Data from each of the four training conditions are plotted separately for the two subjects, Macduff and Oberon. No hints were available during Session 4. All trials, including trials on which the hint was available, were included in the analysis. The horizontal dashed lines represent baseline performance on a new list when hints were unavailable. (Figure reprinted from Kornell & Terrace, 2007, p. 684.)
THE BASIS OF METACOGNITIVE JUDGMENTS
Humans do not seem to be able to make metacognitive judgments by turning an inward eye on their memories and somehow analyzing them directly. Instead, metacognitive processes appear to be inferential (Schwartz, Benjamin, & Bjork, 1997). We make inferences based on external cues—for example, when one is asked a question, the question itself is an external cue—and these evoke internal cues, such as how quickly an answer comes to mind or a feeling of familiarity with the question. Internal cues are the primary basis for metacognitive judgments in humans.
Little is known about the processes underlying animals' metacognitive judgments, although they are also presumably inferential. Understanding those processes—a central issue in human metacognition research—will play an important role in moving the field forward. For a window into those processes, researchers might follow human metacognition researchers' lead and examine animals' metacognitive illusions. For example, processing information fluently makes humans feel confident about their memories—even when excessive fluency undermines learning (e.g., when similar images are displayed in direct succession, rather than at spaced intervals, they create a strong sense of fluency but produce relatively little learning; Kornell & Bjork, 2008). Similar illusions on an animal's part would indicate that it, too, relied on fluency.
METACOGNITION AND CONSCIOUSNESS
One of the most intriguing aspects of metacognition is how it relates to consciousness. It seems logical that if people and animals engage in similar metacognitive processes, and people are aware of some of those processes, animals may be similarly aware. As Hampton (2001) put it, in discussing the finding that monkeys can appropriately avoid tests, “Although it is probably impossible to document subjective, conscious properties of memory in nonverbal animals, this result objectively demonstrates an important functional parallel with human conscious memory” (p. 5359). Smith et al. (2003) drew a similar conclusion.
Not all metacognitive processes are necessarily conscious in people, however. If you were asked the name of a stranger passing on the street, you could say, “I don't know” without consciously feeling uncertain or in other ways considering metacognition. Similarly, if asked your own name you could respond without being conscious of a feeling of confidence. Animals might make metacognitive responses unconsciously as well (Son & Kornell, 2005). Animals know what they know and what they do not know; the human equivalent of these abilities is essentially doing one of two things: answering with confidence, or saying “I don't know” (or “I didn't know” or “I won't know”). Many of the metacognitive processes that become conscious in humans—for example, making a shopping list to avoid forgetting an item, or the tip-of-the-tongue state—have yet to be demonstrated in animals. One reason evidence of metacognitive control is important is that seeking a remedy for one's ignorance goes beyond saying “I don't know.”
CONCLUSIONS AND FUTURE DIRECTIONS
For much of the 20th century, psychologists who studied animals focused almost exclusively on observable behavior. Even a dozen years ago, the idea that animals can monitor their own internal mental states would have been met with skepticism. Today the evidence suggests they do just that: Animals can make judgments about their own memories and seek information when they are unsure.
As outlined above, many questions remain: What metacognitive differences exist between species? Do animals hold beliefs about how memory works (e.g., forgetting happens over time, studying results in learning)? Can animals respond metacognitively without training (and if so, which animals, in which tasks)? Can animals enhance their own memories by making adaptive decisions about how to learn? What processes underlie animals' metacognitive judgments? Most importantly, humans make all sorts of metacognitive judgments that range from simple (I'm sure I know my name) to complex (I usually write in the morning, when I'm most alert). Animals have not yet matched the metacognitive sophistication of their human counterparts. Can animals go beyond the mostly simple, binary metacognitive judgments they have demonstrated thus far? Animals' ability to generalize their metacognitive judgments, and to seek information when they are unsure (Kornell et al., 2007), are small steps in the direction of a richer metacognitive understanding.
The most difficult challenge for future research may be finding more convincing evidence of conscious metacognition. In humans, metacognition is frequently a conscious experience. Evidence that an animal that can behave in ways that parallel conscious human metacognition has the potential to make a unique contribution in the search for proof that animals are, in their own ways, conscious.
Footnotes
Acknowledgements
I am indebted to my human primate collaborators, Herbert S. Terrace and Lisa K. Son, and to my nonhuman primate collaborators, Ebbinghaus, Lashley, Macduff, and Oberon. This article benefited from comments by Bennett L. Schwartz, Lisa K. Son, Jim Kornell, and Sam Kornell.