
Abstract
Interactions between genes and the environment are a critical feature of development. Insights into the dynamic interplay between these factors have come from laboratory studies exploring experience-dependent changes in gene function, which illustrate the importance of environmental factors in determining activity of the genome. These studies have implications for our understanding of the origins of individual differences in behavior and may provide new ways of thinking about the transmission of traits across generations. Here we will highlight how these new findings illustrate the importance of putting genes in context.
Historically, the question of the origins of individual differences in personality, aptitudes, and even physical features has led to debates over nature versus nurture. However, it is becoming increasingly clear that creating a division between genes and environment limits our understanding of the complex biological processes through which individual differences are achieved. The reality that the interaction between genes and environment is a critical feature of development is emerging as a central theme in laboratory studies and longitudinal analyses in human populations. However, appreciating the existence of this interaction is simply the first step in broadening our theoretical approach to the study of behavior. To move forward, we must ask “What do genes do?” and “How do genes and environments interact?” Recent studies combining molecular biology with the study of behavior may provide insight into these issues and perhaps even call into question our current understanding of mechanisms involved in the transmission of traits across generations. Here we will highlight these new findings and illustrate the importance of putting genes in context.
LABORATORY AND LONGITUDINAL APPROACHES TO GENE–ENVIRONMENT INTERACTIONS
Though recent advances in our ability to detect genetic variations have led to rapid progress in the study of gene-by-environment (G × E) effects, clues that G × E was critical in considering the origins of behavior have been available for a long time. In 1958, Cooper and Zubek published a report in which rats selectively bred to be either “maze-dull” or “maze-bright” were reared after weaning in either “enriched” environments containing increased sensory stimuli or “impoverished” environments containing limited sensory stimuli (Cooper & Zubek, 1958). In the rats reared under standard conditions, stable and heritable group differences in cognitive ability were observed in adulthood. However, maze-dull animals reared in an enriched environment showed a significant improvement in learning ability, and maze-bright animals reared under impoverished conditions showed a significant decline in performance. This study provides evidence that, even when considering a genetically derived characteristic, our prediction of behavior must incorporate knowledge of the environmental context of development.
A more recent example of G × E comes from the Dunedin longitudinal study (Caspi et al., 2003), which explored the roles of variation in a gene that alters serotonin levels and exposure to stressful life events across a 20-year period in determining risk of depression. Levels of serotonin within neural circuits are altered by the number of serotonin transporter proteins, and in humans there are genetic variations that lead to either high or low levels of the serotonin transporter. The serotonin system has been implicated in variations in mood, and this system is the target of most pharmacological interventions in the treatment of depression. Among individuals within the Dunedin study, risk of depression was predicted by the interaction of serotonin transporter genotype and the number of stressful life events experienced. Thus, no differences in risk of depression emerged as a function of genotype when the number of stressful life events was low. However, when an individual had experienced a high frequency of stressful events, genotype effects were observed, with individuals possessing the low-serotonin-transporter-level gene variant being at greater risk of depression. Though certain genetic variations can lead to risk or resilience to psychological disorder (see Kim-Cohen & Gold, 2009, this issue), this “potential” may not be observed unless variation in the environment is considered.
CONTEXTUAL DETERMINANTS OF GENE FUNCTION
Empirical findings from G × E studies raise an important question: “If the effects of genetic variation can vary depending on characteristics of the environment, then what are environments doing to genes to alter their impact?” To address this question, we must first address the following question: “What do genes do?” Historically, gene was a term used to describe a unit of heritable material. Since the discovery of DNA, the study of genetics has come to mean the study of DNA, with gene defined as a particular sequence of DNA. Due to the complex nature of DNA, it is perhaps easier to employ an analogy that conveys the basic notions of gene function. Think of an individual's DNA as books in a library that have been ordered and arranged very precisely by a meticulous librarian. These books contain a wealth of knowledge and the potential to inspire whoever should choose to read them. Asking what DNA does is like asking what a book in this library does. Books sit on a shelf waiting to be read. Once read, the information in those books can have limitless consequences. Likewise, DNA sits in our cells and waits to be read by an enzyme called RNA polymerase, leading to the production of messenger RNA (mRNA)—a process referred to as transcription (Fig. 1a). The mRNA transcript is a copy of the DNA sequence that can further be “translated” into protein. The reading, or expression, of DNA can, like the books in our library, have limitless consequences. However, without the active process that triggers such expression, this potential may never be realized. Importantly, it is the environment around the DNA that contains those critical factors that make it possible to read the DNA (Fig. 1b; also see Cole, 2009, this issue, for extended discussion of the regulation of gene expression).
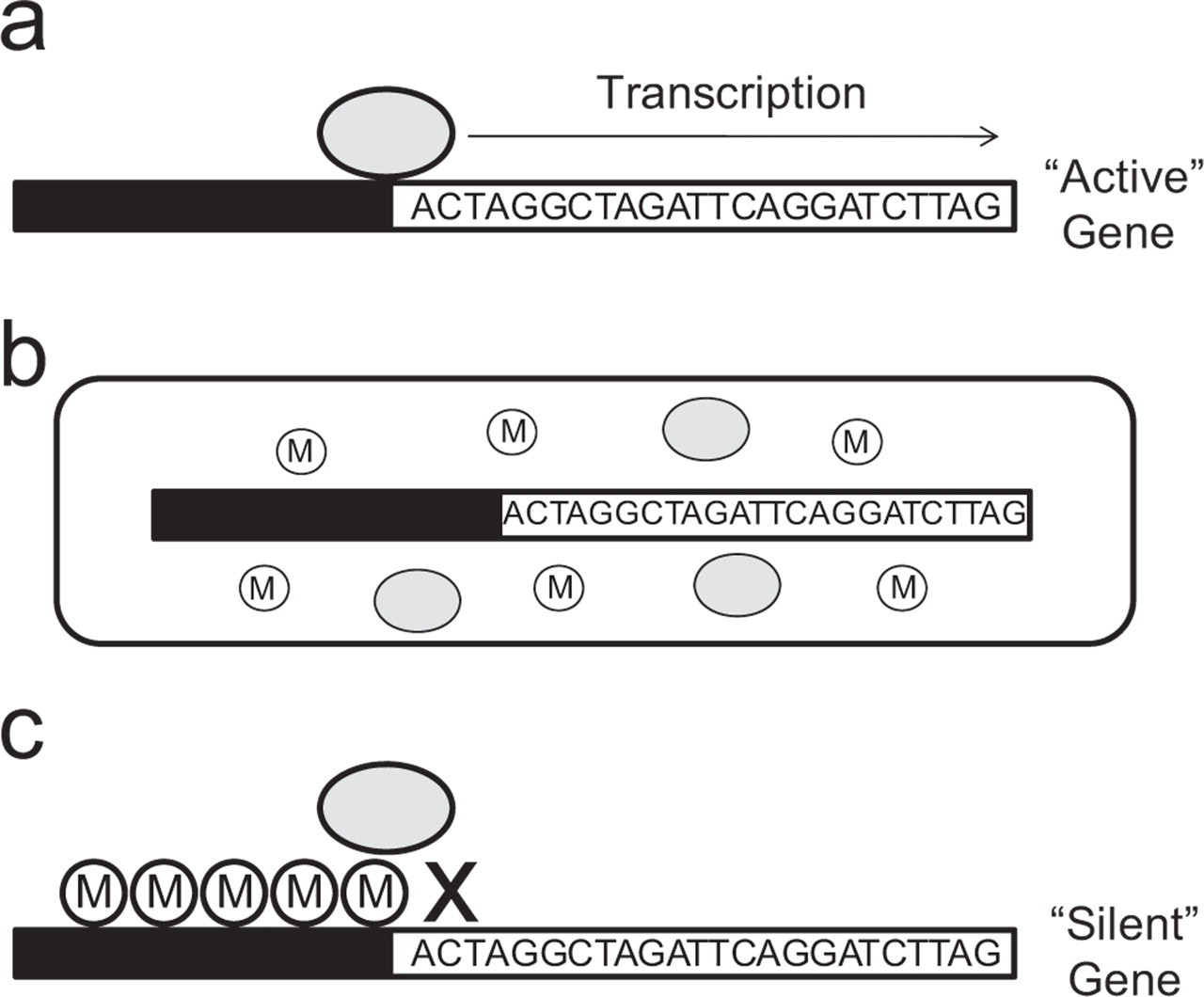
Illustration of the epigenetic control of gene expression and the environmental context of DNA. As shown in the top panel (A), genes consist of a sequence of DNA consisting of “C,” “T,” “A,” and “G” nucleotides preceded by a promotor region of DNA (the black bar). The promoter region responds to factors that control the likelihood of transcription (reading of the DNA). In order for transcription to occur, enzymes that “read” the DNA (the gray oval) must bind to the promotor region of the gene. When this occurs, the gene is “active” and can alter the function of the cell. The environmental context of the gene, shown in the middle panel (B), includes factors that increase gene activity (i.e., enzymes that read the DNA, shown as gray ovals) and factors that decrease gene activity (i.e., methyl groups, illustrated as circles labeled “M”); these factors will determine the likelihood that a gene will be expressed. When a methyl chemical group attaches to the promotor region, as shown in the bottom panel (C), the enzymes that transcribe DNA are blocked and the gene becomes “silent”; this is referred to as DNA methylation.
The control of gene expression is ultimately determined by how accessible the sequence of DNA is to factors within the cell that are involved in transcription. Influences that determine the expression of DNA without altering the sequence of DNA are referred to as epigenetic, meaning “in addition to genetic.” One particular epigenetic mechanism that may have consequences for long-term changes in gene activity is DNA methylation (Fig. 1c). DNA can become modified through the addition of a methyl chemical group to particular sites within the gene sequence. DNA methylation typically reduces the accessibility of DNA and can lead to “silencing” of the gene (Razin, 1998). In the library analogy, one can think of multiple factors that will influence the likelihood a book will or will not be read. Even books containing very valuable information may sit undisturbed and unread, gradually collecting dust. This may be particularly true if the book is hard to get to. It may be located on a shelf that is particularly difficult to reach or blocked by some piece of furniture. DNA methylation reduces the likelihood of transcription much in the same way that shifting furniture in a library can reduce the likelihood that a book will be read. The gene is there, but sits unread, collecting dust.
ENVIRONMENTAL INFLUENCES ON GENE ACTIVITY
A recent breakthrough in our understanding of gene–environment interplay comes from studies exploring the epigenetic processes that are altered by an individual's experiences during development. Based primarily on studies in rodents, these paradigms address the question raised by G × E research: “What are environments doing to genes to alter their impact?” In rodents, variations in maternal care lead to individual differences in the expression of genes that alter the stress response. Low levels of glucocorticoid receptors (GR) within the hippocampus, a brain region critical for learning and memory, result in a prolonged response to stress. Analysis of DNA methylation within the regulatory region of the GR gene indicates that low levels of maternal care are associated with elevated levels of DNA methylation, which epigenetically silence this gene (Weaver et al., 2004). Moreover, the epigenetic status of the GR gene can be targeted pharmacologically in adulthood. Treatment with a drug that promotes increases in accessibility of DNA results in decreased GR methylation and a dramatic shift in the phenotype of adult offspring who received low levels of maternal care (Weaver et al., 2004). Conversely, when adult offspring who experienced high levels of care are treated with a drug that increases the availability of methyl groups within the brain, they become indistinguishable from offspring who received low levels of maternal care (Weaver et al., 2005). These dynamic alterations in DNA methylation in adulthood have also been observed in studies of learning and memory (Miller & Sweatt, 2007). The experience of learning is associated with rapid changes in methylation of genes within the hippocampus, and if DNA methylation is inhibited there will be impairment in memory for the experience. These studies illustrate the role of epigenetic mechanisms in shaping the activity of the genome in response to environmental cues and demonstrate the plasticity that is possible through shifts in DNA methylation.
The prenatal period is characterized by rapid changes in brain development and is thus a sensitive time during which the quality of the environment can exert sustained effects on functioning. In rodents, exposure to chronic variable stress during the first trimester is associated with increased methylation of the regulatory region of the GR gene (Mueller & Bale, 2008). This effect could potentially be mediated by (a) stress-induced decreases in postnatal maternal behavior (Champagne & Meaney, 2006), (b) alterations to gene expression in the placenta (Mueller & Bale, 2008) that may restrict access of the fetus to maternal resources, or (c) a direct influence of maternal stress hormone on fetal gene expression. Modification to the fetal “epigenome” can also be achieved through variations in maternal diet during pregnancy. A striking example of this phenomenon comes from work with a mouse model in which a mutation of the Agouti gene leads to alterations in coat color and metabolism. The severity of the effects of this mutation depends on the level of DNA methylation of the Agouti gene; high levels of DNA methylation will epigenetically silence this mutation and induce a “pseudoagouti” mouse that is comparable in phenotype to a mouse without the mutation. When pregnant female mice with the Agouti mutation are placed on a diet that is rich in methyl groups, the methylation status of this gene is altered such that offspring develop a pseudoagouti phenotype (Dolinoy, 2008). Thus, experience-dependent change in the epigenetic status of genes is not limited to the postnatal period.
IMPLICATIONS OF GENE–ENVIRONMENT INTERPLAY FOR PSYCHOLOGICAL FUNCTIONING
The molecular processes described in laboratory studies may also be critical in understanding the origins of individual differences in humans. Analyses of DNA methylation in cells extracted from fetal cord blood suggest that antenatal maternal depression and anxiety during the third trimester can lead to increased levels of DNA methylation of the GR gene promotor region, having consequences for the stress response of infants at 3 years of age (Oberlander et al., 2008). These effects emerge even in the absence of depression-induced decreases in postnatal mother–infant interactions. The stability of DNA methylation also permits analysis of the epigenetic status of genes in postmortem brain tissue, which can be correlated to life experiences and psychological functioning. In a recent study, DNA methylation of ribosomal genes in hippocampal tissue of suicide victims with a history of abuse and neglect was compared to that of controls. Elevated levels of methylation were detected in ribosomal RNA genes among suicide victims (McGowan et al., 2008), and this effect was found to be specific to the hippocampus. Ribosomes are critical for the production of proteins and thus serve as a critical link between the expression of genes and the level of protein created.
Studies of monozygotic (MZ) twins also provide important insights into epigenetic effects in humans. Comparison of the gene expression of 3-year-old and 50-year-old MZ twins indicates a higher level of discordance in patterns of gene expression among older twins that is associated with increasing differences in DNA methylation in older compared to younger twins (Fraga et al., 2005). Though it is unknown whether concordance in young twins is due to germ-line (the cells that transmit genetic material across generations) or prenatal factors and whether the emerging discordance is random or driven by specific environmental events, there is evidence that epigenetic variation in MZ twins may account for differential risk of mental illness. Analysis of methylation patterns within the catechol-O-methyltransferase (COMT) gene in tissue samples from 5-year-old MZ twins indicates varying degrees of discordance, with some MZ twin pairs showing a high degree of discordance and others being very similar in epigenetic status (Mill et al., 2006). COMT is an enzyme involved in the inactivation of neurotransmitters such as dopamine and norepinephrine, and disruptions in these neurotransmitter systems have been implicated in many forms of psychopathology. The divergence in methylation of the COMT gene within these twin pairs may predict differential risk of neurodevelopmental disorder in later life. Incorporating epigenetic analysis into twin studies represents a novel approach to the study of the origins of individual differences.
TRANSMISSION OF TRAITS ACROSS GENERATIONS: RETHINKING INHERITANCE
In addition to shaping developmental trajectories within an individual's life span, DNA methylation may also have implications for the transmission of traits from one generation to the next. There are two distinct pathways through which this transmission can occur: (a) the behavioral transmission of traits through experience-dependent changes in the methylation of genes, and (b) environmental effects that change DNA methylation in germ cells and are thus transmitted through the germ line of subsequent generations. An example of the first pathway comes from studies of the transmission of maternal care across generations. Variations in maternal care in rodents have been demonstrated to alter the epigenetic status of hypothalamic estrogen receptors of female offspring (Champagne et al., 2006). These receptors are critical in regulating maternal behavior and coordinate the sensitivity of females to hormonal cues. Experience of low levels of maternal care in infancy is associated with increased estrogen receptor promotor methylation, decreased receptor expression, and subsequent decreases in the adult maternal behavior of these offspring. Thus, there is a behavioral transmission of individual differences in maternal care across generations. Interestingly, the quality of environmental conditions experienced by these females at later periods in development can alter this transgenerational inheritance. Prolonged social isolation from peers and prenatal stress can lead to reductions in maternal care that are passed on to subsequent generations (Champagne & Meaney, 2006, 2007). These studies, which are conducted in rodents that have limited genetic variability, suggest that similarities in traits between parental and offspring generations involve far more than the inheritance of genes.
Though epigenetic characteristics of DNA are dynamic in response to environmental cues, these modifications are also stable and heritable. Thus, both genetic and epigenetic factors are transmitted down cell lineages with consequences for the activity of genes within these lineages. However, when considering the question of inheritance at the level of an individual, we must know whether epigenetic patterns within the germ line are correlated to those patterns found within the developing organism. In rodents, prenatal exposure to endocrine disruptors lead to abnormal methylation patterns in sperm cells that are observed several generations beyond the point of initial exposure (Anway, Cupp, Uzumcu, & Skinner, 2005). This germ-line epigenetic inheritance of environmentally induced effects provides further support for the notion that the transmission of traits across generations is not limited in scope to the inheritance of DNA.
CONCLUSION
Just as a library is more than a collection of books, the genome is more than just DNA. The challenge for the field of epigenetics is to determine the origins of the “uniqueness” of each individual's library by exploring the relationship between genetic and epigenetic variation. Though there are many basic questions to be addressed regarding the pathways whereby specific experiences target particular genes, this field of research certainly has promise in uncovering the nature of experience-dependent changes in development both within and across generations. Advances in tools available to study these effects in humans will be critically important in further exploring the role of epigenetics within the broad field of psychological science.