
Abstract
Outcomes of actions, in the form of rewards and punishments, are known to shape behavior. For example, an action followed by reward will be more readily elicited on subsequent encounters with the same stimuli and context—a phenomenon known as the law of effect. These consequences of rewards (and punishments) are important because they reinforce adaptive behaviors at the expense of competing ones, thus increasing fitness of the organism in its environment. However, it is unknown whether similar influences regulate covert mental processes, such as visual selective attention. Visual selective attention allows privileged processing of task-relevant information, while inhibiting distracting contextual elements. Using variable monetary rewards as arbitrary feedback on performance, we tested whether acts of attentional selection, and in particular the resulting aftereffects, can be modulated by their consequences. Results show that the efficacy of visual selective attention can be sensibly adjusted by external feedback. Specifically, although lingering inhibition of distractors is robust after highly rewarded selections, it is eliminated after poorly rewarded selections. This powerful feature of visual selective attention provides attentive processes with both flexibility and self-regulation properties.
Overt behaviors are strongly modulated by the more or less desirable consequences that they have led to in the past. For example, an action repetitively followed by a reward will be more readily elicited on subsequent encounters with the same stimuli and context—a phenomenon known as the law of effect (Thorndike, 1911). The effects of rewards (and punishments) are important because they favor the production of adaptive behaviors at the expense of competing ones, thus increasing fitness of the organism in its environment. However, it is unknown whether similar influences regulate covert mental processes, such as visual selective attention. Visual selective attention allows privileged processing of task-relevant information, while inhibiting distracting contextual elements (Allport, 1989; Duncan, 1993; Egeth & Yantis, 1997; Pashler, 1998).
Here we present data from two related experiments in which we investigated the influence of reward on visual selective attention by means of two negative-priming (NP) paradigms (DeSchepper & Treisman, 1996; May, Kane, & Hasher, 1995; Strayer & Grison, 1999; Tipper, 1985, 2001). NP paradigms typically entail the sequential presentation of a prime display, followed shortly afterward by a probe display, with both requiring a perceptual judgment. The prime consists of a task-relevant target stimulus, together with a distracting stimulus that is potentially interfering and therefore should be ignored. Thus, visual selective attention must intervene to favor processing of the target and to suppress processing of the distractor (Reynolds & Chelazzi, 2004; Tipper, 2001). Usually (DeSchepper & Treisman, 1996; Strayer & Grison, 1999; Tipper, 1985), but not always (Fanini, Nobre, & Chelazzi, in press; Lowe, 1979), the subsequent probe also contains a relevant target, along with a potentially interfering distractor. NP, in the form of lower accuracy and longer reaction times (RTs), occurs when the target in the probe display matches the ignored distractor in the prime display. Although it remains hotly debated whether NP reflects primarily lingering inhibition of a distractor's internal representation or some other cognitive operation, on balance there appear to be solid grounds to believe that inhibitory attention mechanisms play a crucial role in generating NP, perhaps in combination with other perceptual or mnemonic mechanisms (May et al., 1995; Neill & Valdes, 1992; Neill, Valdes, Terry, & Gorfein, 1992; Tipper, 2001; Tipper & Milliken, 1996). In contrast to NP, positive priming (PP) within the same context refers to facilitated performance when the target in the probe display has also served as the target in the previous prime display (Rabbitt & Vyas, 1973; Tipper, 1985).
In the two experiments reported here, participants were presented with prime-probe sequences in which primes always contained a target together with a distractor, in order to engage visual selective attention. Participants were informed that their speed and accuracy of response would be monitored on-line throughout the experimental session, and that each correct response to a prime display (but not to a probe display) would receive a variable monetary reward based on their performance level (high reward for optimal performance, low reward for suboptimal performance). Errors and omissions were also signaled but earned no reward. Subjects were strongly encouraged to respond as quickly and as accurately as possible to both primes and probes, because this would maximize the possible monetary gain. Contrary to this deceptive instruction, however, reward values were determined a priori and could be high (θ0.10) or low (θ0.01) with the same probability (50%) for all conditions.
The effects of the variable reward on performance were assessed by analyzing responses to the probes. Our prediction was that NP (and perhaps also PP) effects would be influenced by the amount of reward delivered after correct responses to the primes. We hypothesized that the lingering effects of inhibitory attention mechanisms, or NP, would be stronger following highly rewarded responses (i.e., following selections regarded as optimal by the subject) and weaker following poorly rewarded responses (i.e., following selections regarded as suboptimal by the subject). In both experiments, there were three priming conditions: (a) attended repetition, AR, in which the probe target had also served as the target in the immediately preceding prime; (b) ignored repetition, IR, in which the probe target had been the distractor in the immediately preceding prime; and (c) control, in which the probe target was different from both the target and the distractor in the immediately preceding prime.
GENERAL METHOD
Participants
A different group of 20 subjects participated in each experiment (Experiment 1: 9 females, mean age = 24; Experiment 2: 13 females, mean age = 24). Written consent was obtained from each of them. Most of the participants were students at the University of Verona, Italy; they were all naive as to the purposes of the experiment and, when queried later, did not express any intuition or suspicion regarding the actual schedule of reward delivery.
Apparatus
Stimuli were presented on a 15-in. computer monitor in a quiet and dimly lit room. The viewing distance was held constant at 57 cm by using an adjustable chin rest. Both experiments were created and run with E-Prime software (Psychology Software Tools, Inc., Pittsburgh, PA), on an IBM-compatible computer.
Procedure
Each prime-probe sequence started with an instruction cue lasting 400 ms. Six hundred milliseconds after offset of this cue, a prime display was presented. This display remained visible for 3 s or until the subject's response. Correct responses were followed by a reward, which could be high (θ0.10) or low (θ0.01), and the amount gained was indicated on the monitor for 600 ms. Four hundred milliseconds later, a probe display was presented for 3 s or until the subject's response (Figs. 1a and 1b). After an intertrial interval of 600 ms, a new sequence started automatically. Both experiments comprised four 1-hr sessions, for a total of 1,536 and 1,440 prime-probe sequences, respectively. In both experiments, the actual sum of money awarded to each subject at the end of the four sessions was chosen randomly between θ72 and θ82 to ensure that all subjects would trust our assertions that gains were proportional to individual performance level and would not spread suspicions among fellow students who might participate later.
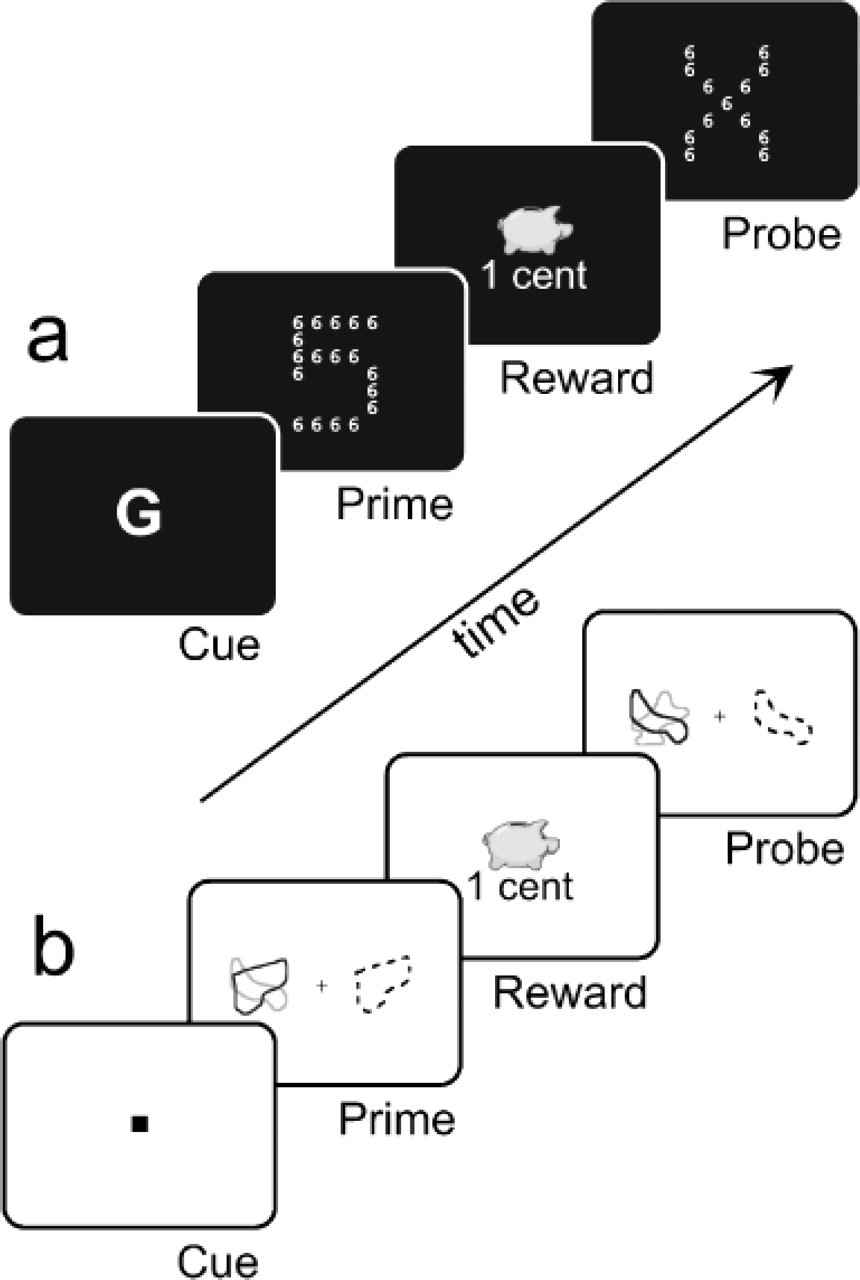
Examples of the prime-probe sequences used in Experiment 1 (a) and Experiment 2 (b). In Experiment 1, primes were global numbers composed of local numbers and were congruent (consisting of the same numerical information at the two levels) in 50% of cases. A letter (G or L) shown prior to each prime indicated whether the global or local level was relevant in the forthcoming display. No cue was delivered before probes, because they contained number information at only one level; they consisted of global Xs made of local numbers or global numbers made of local Xs. In Experiment 2, in both the prime and the probe displays, two overlapping shapes, one red (shown here in black) and one green (shown here in gray), appeared on the left of fixation, while a black shape (shown here as a dashed contour) appeared on the opposite side. A green or red central square cued the color that was relevant for the task in each sequence. Subjects performed a same/different judgment between the shape displayed in the relevant color and the black shape. In both experiments, correct responses to the primes were followed by a reward, which could be high or low, and the amount gained was indicated on the monitor.
EXPERIMENT 1
Stimuli and Task
In Experiment 1, for both prime and probe displays, participants were required to identify the number presented at either the global or the local level in a Navon-like, hierarchical pattern (Navon, 1977; Fig. 1a). The stimuli used were 24 numerical configurations (2°× 2.6°) made up of local items (0.2°× 0.32°) placed within a 5 × 7 matrix. Primes were global numbers composed of local numbers and in 50% of cases were congruent, consisting of the same numerical information at the two levels. Probes instead were global Xs made of local numbers or global numbers made of local Xs. The task was to indicate for each display whether the number shown at a given level was 5, 6, 7, or 8; responses were made by pressing one of four possible response keys with the index or middle finger of either hand. A letter, G or L, appeared prior to each prime to cue the level to be attended (global or local, respectively) in the forthcoming prime. No cue was delivered prior to probe displays because one level was always neutral in these stimuli.
Results
Statistical analyses (analyses of variance and t tests) were performed on RTs for trials with correct responses and on error rates. RTs more than 2 standard deviations from the mean value for each subject and each experimental condition were excluded from analysis. In a preliminary analysis of responses to the prime stimuli, the main factors were relevant level (local or global) and congruency (congruent or incongruent). As in previous studies, results revealed longer RTs (600 ms vs. 533 ms) and higher error rates (8.1% vs. 2.3%) for incongruent than for congruent stimuli, F(1, 19) = 94.12, p < .0001, η p 2= .832, and F(1, 19) = 59.48, p < .0001, η p 2 = .758, respectively. These results reflect Stroop-like response-conflict interference between the competing stimulus levels (Navon, 1977, 1981; Stablum, Ricci, Pavese, & Umiltà, 2001). In addition, overall, RTs were reliably longer to identify a number at the local than at the global level (577 ms vs. 555 ms), F(1, 19) = 5.3, p < .05, η p 2 = .218. This result, also obtained in previous studies, is known as the global-precedence effect (Navon, 1977, 1981; Hoffman, 1980; Kinchla, Solis-Macias, & Hoffman, 1983; Stablum et al., 2001).
The analysis of primary interest was performed on responses to probe displays following incongruent primes (50% of the probe responses), because only incongruent primes must engage active selection and suppression mechanisms. Main factors in the analysis of variance were relevant level (local or global), match between the relevant levels in consecutive prime and probe displays (same or different), reward value (high or low), and priming condition (AR, IR, or control). A significant main effect of relevant level indicated that, again, it took less time to respond to global than to local probes (571 ms vs. 591 ms), F(1, 19) = 17.86, p < .0001, η p 2 = .485. Furthermore, if the relevant level in the probe was different from the relevant level in the prime, a significant increase in RT was observed relative to trials on which the same level had to be identified in the two displays (590 ms vs. 571 ms), F(1, 19) = 60.38, p < .0001, η p 2 = .761. The latter effect may reflect task-switching cost (i.e., the cost incurred when reconfiguration of the task set is required between a prime and the subsequent probe; Allport, Styles, & Hsieh, 1994; Hübner, 2000; Ward, 1982) or some other sequential effect (Schatz & Erlandson, 2003; Shedden, Marsman, Paul, & Nelson, 2003). The main effect of priming condition was significant for both RTs (AR, 543 ms; IR, 598 ms; control, 602 ms), F(1, 19) = 36.28, p < .0001, η p 2 = .656, and error rates (AR, 1.8%; IR, 4.8%; control, 3.8%), F(1, 19) = 16.4, p < .0001, η p 2 = .463. Post hoc contrasts revealed only a robust PP effect (AR vs. control), t(19) = 6.53, p < .0001, for RTs and t(19) = 5.19, p < .0001, for error rates. The main effect of reward value was also significant, showing a general tendency for responses to be faster after low rewards compared with high rewards (high, 586 ms; low, 576 ms), F(1, 19) = 7.43, p < .05, η p 2 = .281.
Most important, a highly significant interaction was obtained between reward value and priming, F(1, 19) = 11.43, p < .0001, η p 2 = .376. Post hoc analyses revealed a reliable NP effect of 11 ms after high reward (IR vs. control), t(19) = 2.1, p < .05, and an opposite (PP-like) effect of 16 ms following low reward, t(19) = 3.08, p < .01. As to authentic PP effects, they were always present in the form of shorter RTs in the AR than the control condition, but they were unaffected by reward value. Crucially, responses in the control condition were not reliably modulated by reward value, t(19) = 0.804, p= .431. Reward interacted significantly also with relevant level, F(1, 19) = 9.35, p < .01, η p 2 = .330, reflecting a reduced global-precedence effect after high versus low rewards (15 ms vs. 25 ms), t(19) = 3.05, p < .01. Finally, there was a significant four-way interaction including all main factors in the analysis, namely, reward value, priming condition, relevant level, and match between the relevant level in prime and probe displays, F(1, 19) = 4.12, p < .05, η p 2 = .178. Preliminary analyses indicated that the joint influence of relevant level in the probe display and match between the relevant level in consecutive prime and probe stimuli was due to the unique influence of the relevant level in the prime. The data presented in Figure 2 were collapsed accordingly. Post hoc contrasts revealed that the modulatory effect of reward on NP was particularly strong when the previous prime had required discrimination at the local level and concurrent suppression of the dominant, global level. In this condition, a standard NP effect of 17 ms was found following high reward, t(19) = 3.162, p < .01, and this effect was replaced by an inverted (PP-like) effect of 26 ms following low reward, t(19) = 3.776, p < .001. In contrast, when the previous prime had required discrimination at the global level and concurrent suppression of the nondominant, local level, priming effects were weaker overall, and they were not reliably modulated by reward value (see Fig. 2).
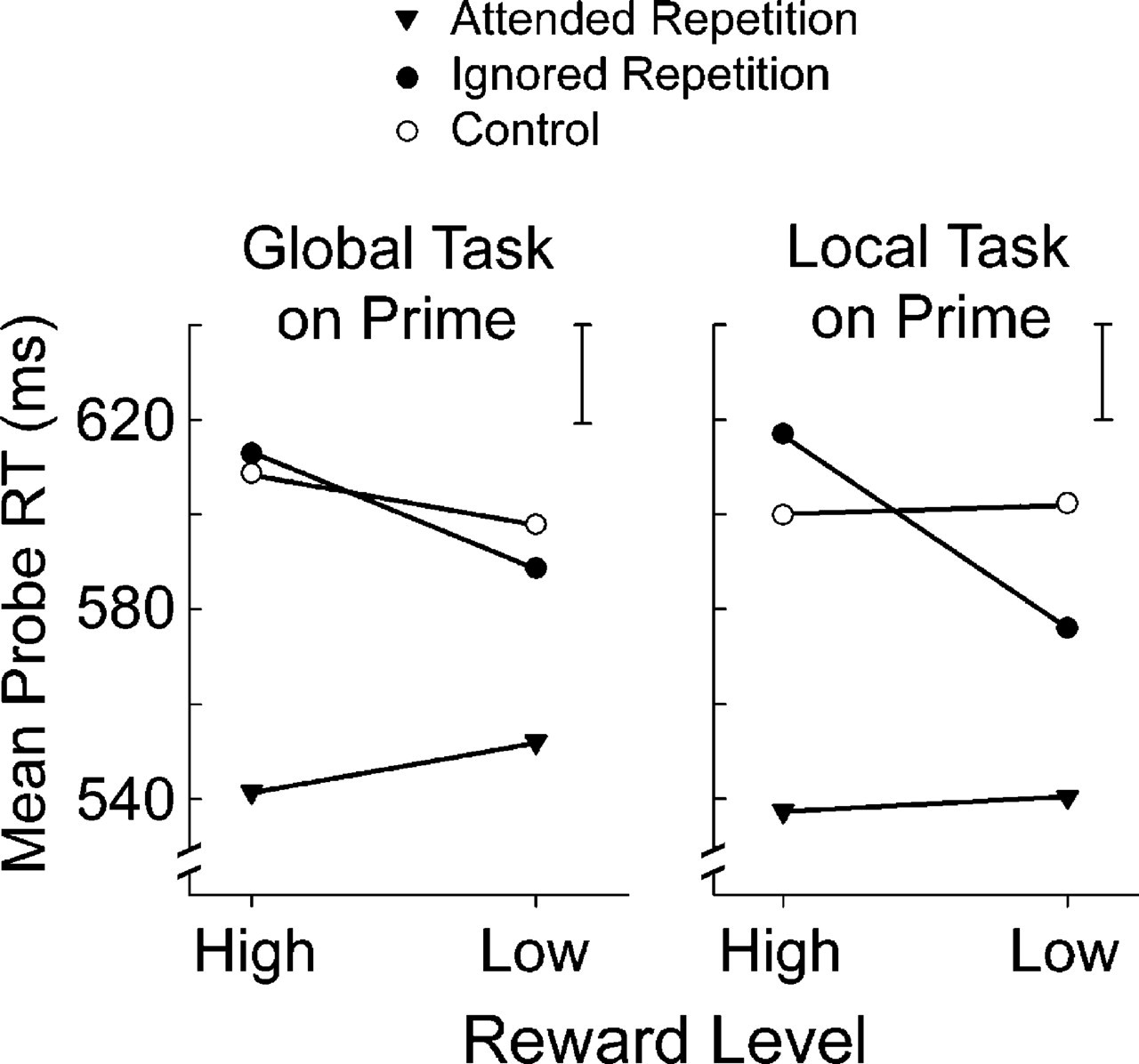
Main results from Experiment 1. Mean reaction time (RT) for probe displays is plotted as a function of priming condition and reward value, for sequences in which the global (left) and local (right) level was relevant in the preceding prime display. The vertical bar at the top right corner of each panel indicates the average standard error of the mean across all points in the plot.
These findings demonstrate that, as we hypothesized, attention mechanisms engaged to select a relevant target and to suppress a concurrent distractor are modulated by the amount of reward delivered to the participant immediately after the act of selection has taken place. In particular, although a robust NP effect was obtained following highly rewarded responses to the prime stimuli, the effect was eliminated (or replaced by a PP-like effect) following poorly rewarded responses to the same stimuli. Note that NP effects and their modulation by reward were particularly evident following prime stimuli requiring selection of the local level and active suppression of the global, dominant level. This observation is in good agreement with the reactive-inhibition view of NP phenomena (Tipper, 2001).
To confirm and extend our findings with entirely new stimuli and participants, and especially with a new paradigm, we set out to perform a second experiment. Experiment 2 again involved the sequential presentation of a prime and a probe. However, unlike in Experiment 1, prime and probe displays both contained a target along with a distractor. Participants performed a same/different judgment between one of two differently colored shapes presented on the left of fixation and a single shape presented on the right (DeSchepper & Treisman, 1996; Strayer & Grison, 1999; see Fig. 1b).
EXPERIMENT 2
Stimuli and Task
The stimuli used in Experiment 2 were 16 outlined nonsense shapes (2°× 2°), selected from the set used by Strayer and Grison (1999). In each trial, two overlapping shapes, one red and one green, appeared on the left side of the monitor, at 3° of eccentricity, while a single black shape was displayed on the opposite side (Fig. 1b). Subjects performed a same/different judgment between one of the shapes on the left and the black shape on the right, indicating their response by pressing one of two response keys with the index or middle finger of the right hand. A green or red square (0.5°× 0.5°) was displayed centrally at the beginning of each prime-probe sequence, signaling which of the two overlapping shapes presented on the left was to be attended during the forthcoming sequence. The color cue was always valid for both the prime and probe displays that followed.
Results
Because all primes were incongruent (the shape in the irrelevant color was always different from the shape in the relevant color), only responses to probes were analyzed: RTs of correct responses (RTs within 2 standard deviations from the mean value for each subject and each experimental condition) and error rates. Main factors in the analysis were priming condition (AR, IR, or control), response type (“same” or “different”), and reward value (high or low). Results are shown in Figure 3.
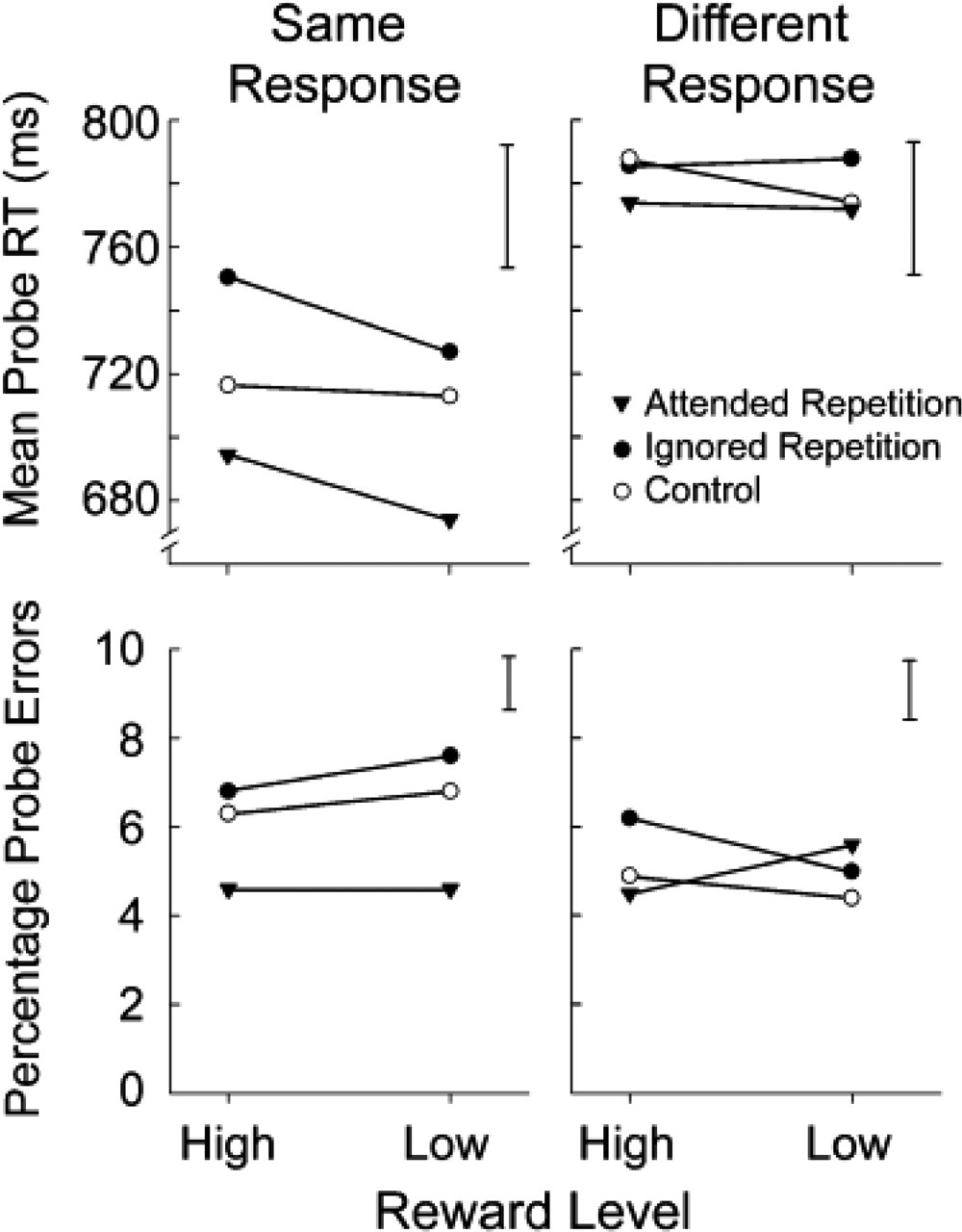
Main results from Experiment 2. Mean reaction times (RTs; top panels) and error rates (bottom panels) for probe displays are plotted as a function of priming condition and reward value. The graphs on the left present results from trials on which the correct response was “same,” and the graphs on the right present results from trials on which the correct response was “different.” The vertical bar at the top right corner of each panel indicates the average standard error of the mean across all points in the plot.
The effect of priming was highly significant (AR, 728 ms; IR, 763 ms; control, 748 ms), F(1, 19) = 18.22, p < .0001, η p 2 = .489. Both the AR and the IR conditions differed reliably from the control condition, t(19) = 3.8, p < .001, for PP and t(19) = 4.42, p < .001, for NP. The effect of response type was also significant, with “same” judgments yielding faster RTs than “different” judgments (712 ms vs. 780 ms), F(1, 19) = 49.88, p < .0001, η p 2 = .724. Again, reward value was significant, with overall faster responses following low rewards than high rewards (741 ms vs. 751 ms), F(1, 19) = 5.52, p < .05, η p 2 = .225. In addition, a significant interaction between response type and reward value, F(1, 19) = 4.96, p < .05, η p 2 = .207, showed that the speeding up of RT after low rewards occurred only for “same” judgments (high reward, 720 ms; low reward, 705 ms), t(19) = 3.29, p < .01. Response type interacted significantly also with priming condition, F(1, 19) = 15.66, p < .0001, η p 2 = .452: Both NP and PP effects were reliably stronger for probes requiring a “same” response than for probes requiring a “different” response—NP effect: 24 ms versus 6 ms, t(19) = 3.29, p < .001; PP effect: 30 ms versus 8 ms, t(19) = 3.2, p < .001.
Finally, and crucially for our purposes, a significant three-way interaction including all main factors in the analysis was found, F(1, 19) = 3.81, p < .05, η p 2 = .167. Post hoc contrasts demonstrated that reward value interacted significantly with priming condition only for “same” judgments to probes, but not for “different” judgments. This result was expected given the previous observation that priming effects were overall much stronger for “same” than for “different” judgments. The results are consistent with those of Experiment 1 in that NP was robust and significant when the previous prime was followed by high reward (IR vs. control, 34 ms), t(19) = 3.89, p < .001, but weak and nonreliable when the previous prime was followed by low reward (13 ms, n.s.; see Fig. 3). Unlike in the previous experiment, PP effects were also modulated by reward value. Although they were significant overall, they increased in magnitude after low rewards (high reward, 22 ms; low reward, 39 ms), t(19) = 2.61, p < .05. Crucially, as in the previous experiment, reward value had no significant influence on RTs in the control condition, t(19) = 1.418, p= .171.
A significant three-way interaction was also found in the analysis of error rates, F(1, 19) = 4.09, p < .05, η p 2 = .177, which showed a reliable effect of reward value on priming for “different” judgments only. Once again, only highly rewarded prime responses were followed by a significant NP effect (1.25% increase in error rate), t(19) = 2.44, p < .05 (Fig. 3).
GENERAL DISCUSSION
The present results demonstrate for the first time a powerful influence of reward on the working of visual selective attention, in particular, on the strength or duration of the lingering effects produced by the intervention of inhibitory attention mechanisms. In both experiments, NP was consistent following highly rewarded selections, but was eliminated following poorly rewarded selections. In Experiment 1, NP was actually replaced by a paradoxical, PP-like effect following poorly rewarded responses. The results suggest that following a poorly rewarded selection, the inhibition applied to the distractor is lifted quickly, as if the attentive system were rapidly reset by feedback information (low reward) indicating relatively unsuccessful performance. The present findings suggest that mechanisms of visual selective attention are adaptively adjusted to optimize interaction of the individual with the environment. Such a feature may be fundamental in providing attentive processes with both flexibility and self-regulation properties.
Unlike the strong and consistent reward-dependent modulation of NP, PP was less consistently affected by reward value, in line with the notion that PP effects primarily index automatic consequences of stimulus repetition (Schacter & Buckner, 1998). More generally, the effects exerted by reward value revealed a remarkable degree of specificity. In both experiments, reward value per se had no reliable effects on the speed and accuracy of responses to the immediately subsequent probe displays in the control condition. The latter finding is especially important because it allows us to distinguish the highly specific reward-dependent modulation of NP demonstrated in the present study from other behavioral effects reported in the literature, such as the effects elicited by false-feedback manipulations (e.g., de Bruijn, Mars, & Hulstijn, 2004; Miltner, Braun, & Coles, 1997).
In our paradigms, reward value was completely decoupled from actual behavioral performance, as high and low rewards were each delivered arbitrarily on 50% of the trials according to a schedule that was independent of the subjects' proficiency in the task. This ensured that the reported effects of reward value were not contaminated by factors relating to the perceptual and motor processes involved in task execution.
It has been suggested that successful identification of a target may be sufficient to trigger an endogenous reward signal, acting as a positive reinforcement and allowing perceptual learning (Holroyd & Coles, 2002; Schultz, 2000; Seitz & Watanabe, 2003). Analogously, when no other feedback is available, the correct selection of a target, and concurrent filtering of distractors, may initiate a form of self-reward. It is within this framework that low rewards, although delivered after correct responses, may interfere with the active suppression of distractors by reducing the strength or duration of the inhibitory mark applied during attentive selection.
In the present study, modulatory effects of reward affected NP occurring with prime-probe delays in the order of a few seconds. However, prior work has demonstrated that, under ad hoc conditions, NP can span much longer intervals between prime and probe episodes, up to several weeks (DeSchepper & Treisman, 1996). Therefore, it will be particularly important to assess whether an influence of reward value similar to the one reported here can also take place in NP paradigms with longer prime-probe intervals.
Footnotes
Acknowledgements
This work was supported by grants to L.C. from the Human Frontier Science Program (HFSP) and the Italian Government (MIUR).