
Abstract
Parents of 162 infants ages 6 to 20 months modeled subsets of four of the same set of eight behaviors, each for a maximum of 3 min, and encouraged their infants to imitate. Proportions of infants producing each behavior (a) when it was modeled and (b) during modeling of a different behavior were compared to estimate the age at which infants mimicked each kind of behavior. No reproduction of these motor acts—that is, no mimicry—was observed at 6 months. Mimicry appeared to develop slowly through most of the 2nd year, emerging at different ages for different behaviors. The findings suggest that newborns' behavioral matching may not be continuous with mimicry later in infancy. Imitation is probably not one behavioral competency with one underlying mechanism. It is more likely a category of different ways of combining and using different types of knowledge, some of which develop across the first 2 years of life.
This article presents a cross-sectional study of the imitative behavior of 162 infants ranging in age from 6 to 20 months. In this study, each infant saw a small number of different behaviors repeated by a parent, each for as long as 3 min, and was encouraged to perform the same actions. The primary measure was the proportion of infants at each age level who matched each behavior. At issue are the questions of when and how the capacity to imitate develops.
Imitation is currently a subject of interest to researchers in a wide range of fields, including neuroscience, cognitive science, animal behavior, and robotics, as well as developmental science. Understanding the origins of the imitative capacity, how it works, and how it changes over time would have broad theoretical implications for each of these fields, and possibly practical value for researchers attempting to model or reproduce the capacity to imitate in nonhuman devices.
Despite its wide interest and relevance, the development of imitation has not been the subject of a great deal of research. This may be because it is widely accepted among developmentalists that the ability to imitate is innate, and that much of the underlying mechanism is inherited. The basis for this belief is a series of reports, beginning with one by Meltzoff and Moore (1977), that newborn human infants will imitate the behavior of an adult model (see Anisfeld, 1996, and Butterworth, 1999, for reviews). Neonates have been reported to imitate mouth opening, tongue protruding, pouting, sequential finger movements, and head turning (Butterworth, 1999). The evidence is surprisingly limited and has been challenged on both theoretical and empirical grounds (e.g., Abravanel & DeYong, 1997; Anisfeld, 1996, 2005; Anisfeld et al., 2001; Jones, 1996, 2005, 2006). Nonetheless, the claim that newborn infants can imitate is a foundational idea in many theories, both within and beyond psychology (Hurley & Chater, 2005).
In studies of infants beyond the newborn period, imitation appears largely as deferred imitation, a measure of memory (e.g., Cuevas, Rovee-Collier, & Learmonth, 2006; Meltzoff, 1988). Deferred imitation differs from the kind of imitation claimed for newborns. Want and Harris (2002) offered the term mimicry for the reproduction of another person's specific muscle movements, as in newborns' tongue protruding or mouth opening. In deferred imitation, infants reproduce—after some delay—the effects of another person's actions on an object, but may or may not use the same movements to produce the same effect. Want and Harris suggested the use of Tomasello's (1998) term emulation for actions that reproduce the effect or outcome of another person's behavior.
The present study examined the emergence and subsequent development of infants' ability to reproduce eight motor actions performed by their parents. In four cases, the behaviors modeled for the infants resulted in sounds, so reproduction of those actions might be considered emulation. In the other four cases, however, the specific movement made no sound.
This study is unique in focusing on infants' matching of a standard set of specific motor actions from the middle of the 1st year through most of the 2nd. The developmental course of mimicry beyond the newborn period has been the subject of only a handful of previous studies, and the focus has been on mimicry of preverbal sounds and words during social interactions (e.g., Kokkinaki, 2003; Kokkinaki & Kugiumutzakis, 2000; Masur & Eichorst, 2002; Papousek & Papousek, 1989). Most mimicry of sounds found in the 1st year has been produced not by infants, but by their adult social partners. For example, Kokkinaki and Kugiumutzakis (2000) recorded interactions between 15 parents and their infants every 2 weeks from 2 to 6 months postpartum. Parents matched their infants' vocalizations two to three times in each 10-min session. Infants, however, matched their parents only about once in every 20 to 30 min of interaction. It seems likely that this low rate of sound matching by infants was due to chance. In a study supporting this inference, Siegel, Cooper, Morgan, and Brenneise-Sarshad (1990) examined vocal imitation by infants between 9 and 12 months of age and found “no indications that the children imitated the vocal patterns of their speaking partners” (p. 9).
Pawlby (1977) studied mimicry of all kinds of behavior in 8 infants weekly from about 4 to 10 months of age. As Kokkinaki and Kugiumutzakis (2000) found, parents imitated their infants at quite high rates—on average about seven times in each 10-min session. Infants matched parents' behaviors of all kinds only once or twice in the same interval. Thus, again, infants' behavioral matches were infrequent, and may have been chance occurrences rather than mimicry. Interestingly, infants in this study, like those in the study by Siegel et al. (1990), failed to match their mothers' sounds. Instead, they matched their mothers' actions on objects: that is, they emulated. Masur and Rodemaker (1999), studying mimicry at 10, 13, 17, and 21 months of age, similarly reported that their infant subjects mostly matched actions on objects until the age of 17 months.
One longitudinal case study (Jones & Yoshida, 2006) has tracked the development of mimicry of all sorts in a female infant named Yo. Yo was videotaped weekly from 3 to 16 months of age in 10-min face-to-face interactions with her mother. No objects were used. Initially, the interactions were natural and unconstrained. Later in the study, the mother was encouraged to make direct attempts to get Yo to mimic her behaviors. The resulting developmental sequence was strikingly similar to that observed by Piaget (1945) in his own 3 infants. Specifically, at 3 months, Yo matched nothing—although her mother mimicked Yo a lot. Yo's first behavioral matches were observed in the second half of her 1st year. The actions matched were behaviors (a) that Yo had spontaneously and repeatedly produced at home, on her own; (b) that Yo's parents had imitated; and (c) that produced or were accompanied by sounds. For example, when Yo spontaneously raised both arms, her mother said, “Bonsai!” After several sessions, the mother could say, “Bonsai,” while raising her own arms, and Yo would raise her arms. However, for a long time, just the sight of the mother raising her arms was not by itself a sufficient cue for Yo to raise her arms. Thus, the sight of the behavior did not elicit mimicry, but the word cued behavior that could have been mistaken for mimicry.
In brief, as Piaget described, sounds appeared to be important learned cues for Yo's eventual matching of some of her mother's behaviors. Certainly, Yo matched behaviors cued by sounds before she matched silent behaviors. And Yo matched behaviors she could see herself do long before behaviors she could not see herself do. Yo did not match her mother's tongue protruding until 14 months of age.
The study reported here addresses three questions raised by the observations of Yo's slow acquisition of mimicry. First, when do infants in general begin to match other people's behaviors? Because Kokkinaki and Kugiumutzakis (2000) found so little infant behavioral matching in the 1st half-year, the present investigation began with 6-month-olds. Second, what kinds of behaviors do babies match first? Findings from Pawlby (1977), Masur and Rodemaker (1999), and the literature on deferred imitation suggest that babies will emulate before they begin to mimic. However, the literature on newborn imitation puts mimicry first in development (Want & Harris, 2002). Third, is there a special role for sound in initially linking behaviors that infants perform with behaviors that they observe? Infants in previous studies have not mimicked specific sounds. However, Piaget's (1945) writings suggest that behaviors that produce sounds are likely to be matched first, as the sounds become learned cues for infants' production of behaviors that incidentally match the behaviors of a social partner.
METHOD
Subjects
Subjects were 162 infants within 1 week of 6, 8, 10, 12, 14, 16, 18, or 20 months of age, each with his or her primary caregiving parent. There were 21 infants (10 males) each in the 6- and 10-month-old groups, and 20 (10 males) at each of the other age levels. An additional 42 infants began but did not complete the study because of fussing (34), experimenter error (5), or equipment failure (3). Infants were recruited from county birth records and were predominantly European American and middle- to lower-middle-class.
Modeled Behaviors
Each infant saw four behaviors modeled by a parent. There were eight behaviors in all—two in each of four categories. Each infant saw one of the two behaviors from each category, with the behaviors presented in one of eight predetermined combinations and randomized orders. The four categories were formed by crossing two criteria, each with two values: (a) whether or not the behavior produced sound and (b) whether or not the infant's own behavior would be visible to him or her (see Table 1). The two visible behaviors that made sounds were hitting the surface of a table or high-chair tray with the fingers of one open hand (Tap Table) and bringing the palms of the two hands smartly together (Clap Hands). Visible behaviors that made no sounds were Bye-Bye, flexing and straightening all four fingers of one hand with the palm facing the partner, and Sequential Finger Movements (SFM—Meltzoff & Moore, 1977), flexing the fingers of a raised hand in sequence from little to index finger. Behaviors that made sound but were not visible were AAh, a single /a/ vowel sound, and Eh-Eh, paired /e/ sounds. Behaviors that made no sound and were not visible were raising one arm to the side to touch the head (Hand on Head) and protruding the tongue beyond the inside line of the lower lip (Tongue Protrusion—Meltzoff & Moore, 1983). Each behavior was chosen in part because it could be performed by infants across the entire age range studied. Note that two of the eight behaviors—SFM (Meltzoff & Moore, 1977) and Tongue Protrusion (e.g., Meltzoff & Moore, 1977, 1983)—are reportedly imitated by newborn infants.
Description of and Results for Each of the Eight Modeled Behaviors
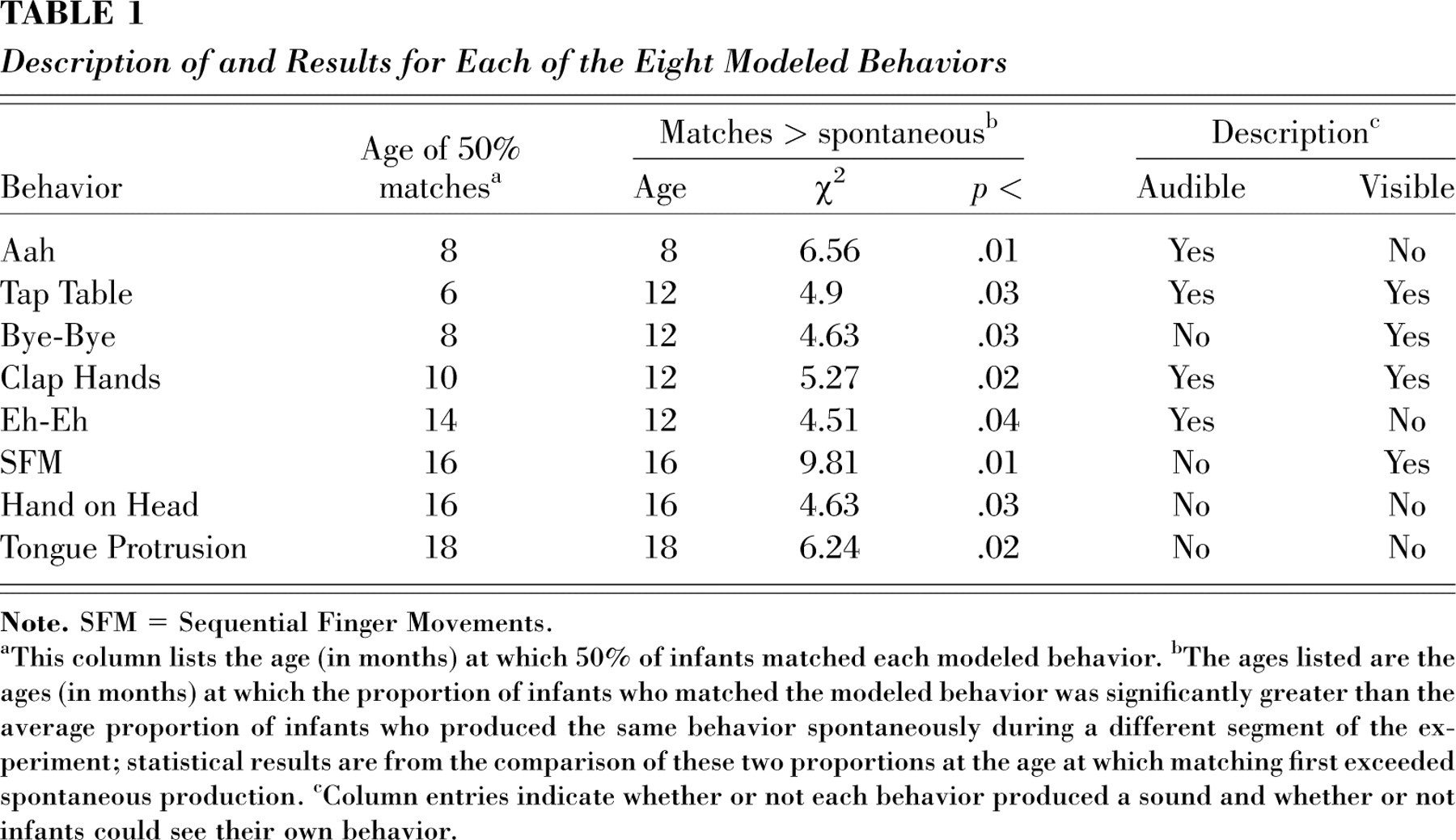
aThis column lists the age (in months) at which 50% of infants matched each modeled behavior.
bThe ages listed are the ages (in months) at which the proportion of infants who matched the modeled behavior was significantly greater than the average proportion of infants who produced the same behavior spontaneously during a different segment of the experiment; statistical results are from the comparison of these two proportions at the age at which matching first exceeded spontaneous production.
cColumn entries indicate whether or not each behavior produced a sound and whether or not infants could see their own behavior.
Eight separate DVDs were recorded. On each DVD, an experimenter provided a moment-to-moment guide for modeling a different set of four behaviors in a random sequence. On each of four separate tracks, the experimenter first demonstrated the behavior to be modeled; then said, “Begin”; and then counted out the rhythm for presentation of that behavior with an extended forefinger. Pauses in modeling were signaled with upraised palms. The experimenter counted out 10-s intervals of modeling and 10-s intervals of rest in alternation for a total of 3 min for each behavior. Alternating periods of behavioral modeling and rest are a common feature of experiments on newborn imitation, and according to Meltzoff and Moore (1983), most imitation occurs when the model rests. Behaviors were timed to occur either once (Aah, Eh-Eh, Bye-Bye, SFM, Hand on Head, Tongue Protrusion) or twice (Tap Table, Clap Hands) per second, followed by a 1-s pause.
Because parents glanced at the video demonstration of behaviors from time to time, they did not maintain constant eye contact with their infants. Models in experiments on newborn imitation have typically fixed their gaze on a clock (Meltzoff & Moore, 1983), both to time movements and to avoid reinforcing the infant for imitating. In the present study, parents interacted with their infants more naturally. They typically glanced at the demonstration video repeatedly but very briefly during each modeling segment after they had seen the form and rhythm of the model's first actions. Parents also were free to encourage their infants to mimic, making comments (e.g., “You do it. Do this.”) that did not label the specific behavior being modeled.
Setting
Data were collected in a small (3.08 m × 2.4 m) laboratory room with two remote-controlled wall-mounted cameras and a one-way observational window. The room contained a high chair, two armless chairs, and a video monitor. Parent and infant were seated opposite one another, and the video monitor was located behind and above the infant's head. A black screen on the floor behind the infant's chair rose to the infant's head level, blocking the monitor for older infants who attempted to follow their parent's gaze. Parents had a single “earbud” to listen to the video model's instructions. Six-month-old infants and some 8-month-olds sat on an experimenter's lap. In general, infants 8 to 16 months of age sat in the high chair with the tray in front. Infants 18 and 20 months of age generally sat at a small table.
The two digital cameras provided separate close-up views of the face and upper body of the infant and parent. Images of parents occupied about one sixth of the screen in one corner; images of infants occupied the rest of the screen.
Procedure
After the infant was seated opposite the parent, the first segment of the DVD for that infant was played. The experimenter observed the parent and infant through a one-way mirrored window. If the infant produced an unambiguous instance of the behavior being modeled by the parent, the experimenter reentered the room, stopped that segment of the DVD, and either began to play the next segment with the next behavior to be modeled or announced a break. Although the initial plan was to measure the frequency of infants' behavioral matching across entire 3-min segments, pilot testing indicated that few of the younger subjects would complete the experiment unless the total time was reduced in this way. Breaks were frequently given between modeling segments, but never during a segment, and often included brief intervals of snacking.
Coding
Coders could not be unaware of the study's purpose, but were taught that the study was exploratory and that all potential outcomes were equally interesting. Eight coders recorded whether or not each infant had performed any of the eight behaviors during the modeling of the four behaviors that infant saw. One primary coder coded behaviors of all infants at a given age level, and a second reliability coder coded a minimum of 5 (25%) of the same tapes. Reliabilities, calculated as agreements/agreements + disagreements, ranged from .94 to 1.0; kappas ranged from .72 to 1.0, and averaged .89.
RESULTS AND DISCUSSION
The proportion of infants at each age level who produced each behavior while it was being modeled was determined directly by the coding. In addition, each behavior had some probability of occurring spontaneously with or without a model. This probability was estimated by calculating, for each behavior at each age level, the average probability that an infant who did not see the target behavior modeled produced that behavior spontaneously during the parent's modeling of one of the other actions. For example, twelve 6-month-old infants saw a parent model Tap Table. Of those, 8 matched this behavior during the modeling segment (p matching = .67). Another nine 6-month-old infants did not see Tap Table modeled. Each of those infants had four opportunities to produce Tap Table during the modeling of some other behavior. These 9 infants produced Tap Table during 24 of those 36 modeling periods. Thus, the average probability that a 6-month-old who had not seen a model of Tap Table spontaneously produced this behavior during any modeling segment (p spontaneous) was also .67.
Figure 1 shows p matching and p spontaneous at each age level for each of the eight modeled behaviors. The age at which p matching was significantly greater than p spontaneous is shown in Table 1, and also in Figure 1 as the point in the “Modeling” curve at which the broken line becomes solid.
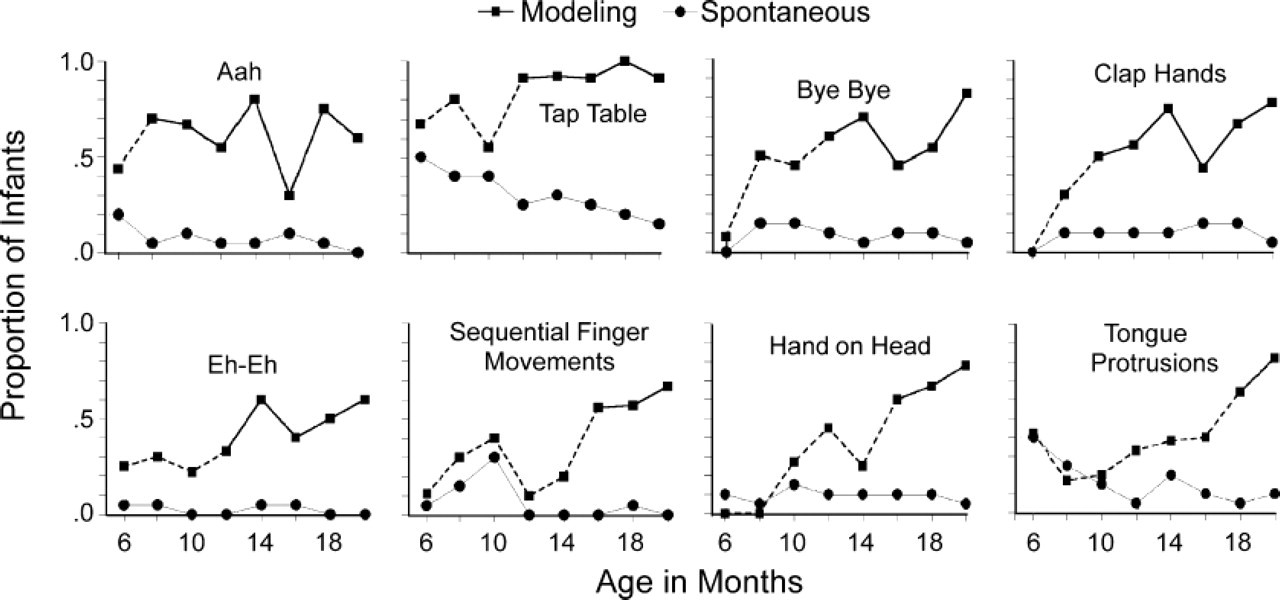
The proportions of infants at eight age levels who matched each of eight behaviors. Different infants saw different sets of four behaviors modeled by a parent. The two curves for each behavior show (a) the proportion of infants producing the behavior while parents modeled the same behavior and (b) the proportion of infants spontaneously producing the behavior (without ever seeing the behavior modeled) while parents modeled one of the other seven behaviors. In the former curves, the change from a dotted to a solid line indicates the age at which production during modeling first significantly exceeded spontaneous production.
Two general questions are addressed by the data in Figure 1: First, how common is voluntary behavioral matching in infants from 6 to 20 months of age? Second, which types of behaviors are matched earliest, and which are matched later in the age range?
Overall, there was little behavioral matching—even with much encouragement—until around 12 months of age. There was no evidence of any voluntary behavioral matching in 6-month-old infants. Four of the behaviors—Clap Hands, Bye-Bye, SFM, and Hand on Head—were virtually never observed at this age, either when modeled or in any other segment of the experiment. Two-thirds of 6-month-olds matched Tap Table, and about 40% matched Tongue Protrusion, but both behaviors were just as likely to occur while a different behavior was modeled. Tongue protruding is a common response of very young infants to any interesting distal stimulus (Jones, 1996, 2006), and at 6 months may still reflect interest in the parent's actions.
For the remaining two behaviors—the vocalizations Aah and Eh-Eh—6-month-old infants were more likely to match the behavior when it was modeled than to produce the behavior spontaneously, but not significantly so. Thus, it is possible that some small number of 6-month-old infants did mimic either or both of these vocal behaviors, but in general, mimicry could not be detected at this early age.
The failure to find mimicry in 6-month-old infants may seem surprising in view of the numerous reports of imitation in newborn infants. The difference in findings cannot be attributed to a difference in the behaviors modeled, as the current set included two—SFM and Tongue Protrusion—that were taken, along with their coding standards, from the literature on newborn imitation. In the present study, infants did not mimic either of these behaviors until well into the 2nd year.
Figure 1 provides values for two criteria for the age of emergence of each type of behavior: first, the point, already described, at which the behavior was produced by significantly more infants while watching that behavior than while watching another behavior being modeled, and second, the age at which half or more of the infants matched the behavior during the modeling period. Table 1 lists the eight behaviors in order of the emergence of matching as suggested by these two numbers together. The table also reports chi-squares for the age levels at which p matching was significantly greater than p spontaneous.
There were too few different behaviors to allow for standard runs tests. However, as Table 1 shows, by and large, behaviors that made sounds were matched at younger ages than behaviors that did not make sounds, and behaviors that were visible to the infant were matched at younger ages than behaviors that were not visible. The importance of sounds suggests that emulation precedes mimicry in development, or that sound cues provide vital bridges to motor mimicry for infants (Piaget, 1945). The primacy of visible behaviors suggests that early motor mimicry is not based primarily on proprioceptive representations of the infant's own behaviors, as has been suggested by Meltzoff and Moore (1997).
The ordering of the eight behaviors in Table 1 is based on age of first evidence of mimicry. However, the developmental trends of ordered pairs in the table are strikingly similar. Figure 2 shows the curves of successive pairs of behaviors together. The curves from Figure 1 have been smoothed to make the overall developmental trends more visible. Each curve is anchored by the 6-month-old and 20-month-old groups' raw proportions of matching, but each point between shows the average proportion of matching over three successive age levels.
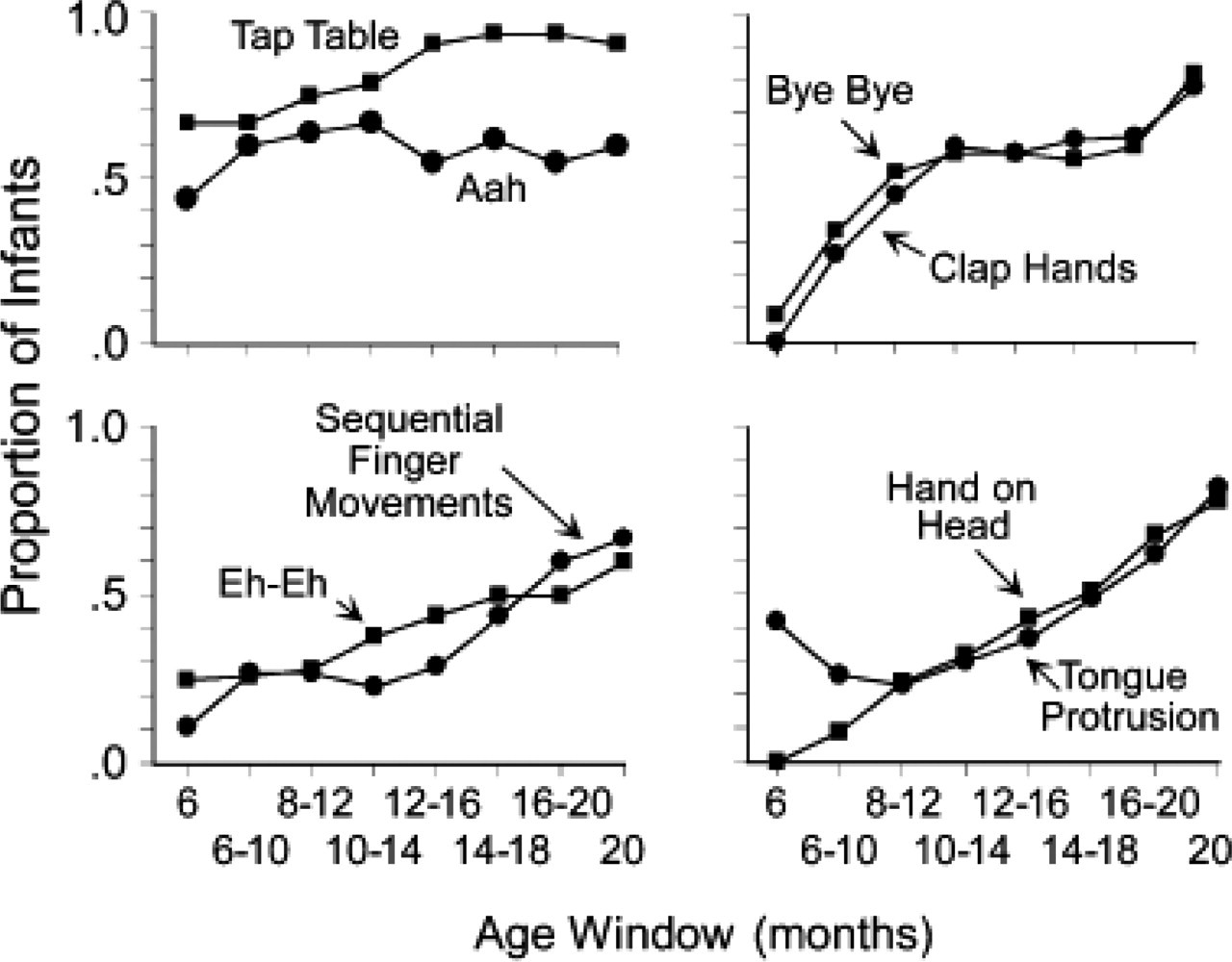
The proportion of infants who matched each modeled behavior, at each of eight age levels. The behaviors are paired according to the order of emergence of mimicry, as shown in Table 1. The curves have been smoothed by averaging the proportions at three successive age levels to determine each point, except for the anchors, which show the raw proportions at ages 6 and 20 months.
The two behaviors that were matched first—roughly between 8 and 12 months of age—are Tap Table and Aah. Both produce sounds that could act as cues for subsequent infant behaviors. Many infants produce these sounds independently, in repetitive sequences of arm flapping or cooing. Piaget (1945) proposed that imitation begins with behaviors like these, repeated by the infant as “primary circular reactions.” With repetition, each iteration of the behavior becomes a cue for the next. “Imitation” starts when parents intrude their own mimicry of the infant into the sequence, and thus cue the infant to continue the repetitions.
The next two behaviors to be mimicked, by about 10 to 12 months of age, were Clap Hands and Bye-Bye. The developmental trends for these two behaviors are almost exactly the same. This may be because both of these behaviors are commonly tutored by parents, who model and reward approximations. Mimicry of these actions may also begin with a parent's imitation of the infant, resulting in paired-associate learning through which the parent's actions and words become cues for the infant's production of the associated behavior.
SFM and Eh-Eh were the next two behaviors to be mimicked, starting in the first few months of the 2nd year. Both require more fine motor control than any of the preceding behaviors. Neither is commonly tutored by parents. Thus, it is likely that infants must independently discover the body parts and movements that produce the sound of Eh-Eh and the sight of SFM. That these discoveries are slow to come is suggested by the finding that only two thirds of 20-month-old infants matched these behaviors.
The last two behaviors to be mimicked, at around 16 to 18 months of age, were Hand on Head and Tongue Protrusion. The developmental trends for these two behaviors are almost identical, suggesting that mimicry of these behaviors requires the same kinds of knowledge. These behaviors are not typically cued by sound, and neither is visible to the infant when he or she produces it. Thus, infants must know their own body and its movements well in order to reproduce either out-of-sight behavior without explicit tutoring.
Overall, the findings from this study indicate that the ability to imitate specific motor acts takes most of the first 2 years to develop. The fact that different kinds of behavior are first mimicked at different ages suggests that imitation is not a single competency that appears all at once and is then applicable to any and all behaviors. Instead, the drawn-out developmental course shown here is likely to reflect changes in infants' knowledge of their own body parts and the behavioral potentials of those parts, and also changes in infants' social knowledge and motivations.
The present findings are at odds with current beliefs that newborn infants imitate the behaviors of other people. However, the data are consistent with previous failures to find convincing evidence of spontaneous imitation during social interactions in infants between 1 or 2 months and 6 months of age (Kokkinaki & Kugiumutzakis, 2000; Pawlby, 1977), and with the failure to find behavioral matching in imitation experiments in 3-month-olds (e.g., Heimann, Nelson, & Schaller, 1989). All of these findings indicate that the behavioral matching found in experiments on newborn imitation is not continuous with the mimicry that begins toward the end of the 1st year.
The discontinuity may exist because early behavioral matching is temporarily masked by developmental events and reemerges later. Thelen and Fisher (1982) showed that the “stepping” observed in newborns is masked when infants' legs become too fat for their muscles to lift, but is still present as supine kicking. Stepping then reemerges with increasing muscle-to-weight ratios at about 8 months. Stepping, however, is a reflex, whereas imitation is a complex voluntary cognitive act.
Alternatively, the discontinuity may exist because the behavioral matching seen in experiments on newborn imitation and the behavioral matching seen in the present study are produced by different mechanisms and not from the same underlying core abilities (e.g., Clearfield & Mix, 1999; Gershkoff-Stowe & Thelen, 2004; Jones, 1996, 2005).
In summary, the present data suggest that imitation is not a single competency, with a single underlying mechanism. Instead, it is likely that imitation is a collection of different ways of combining and using different kinds of knowledge (see Want & Harris, 2002). Mimicry is one form of imitation, and its drawn-out developmental course suggests that it, too, is not the output of a single specialized mechanism, but instead is the emergent product of a large number of component kinds of motor, cognitive, and social knowledge and motivation. Each component is likely to have its own complex developmental course (Thelen & Smith, 1994).
If this is correct, then the origins of imitation (the mechanisms that account for the development of this category of behaviors) and the nature of imitation (the mechanisms producing moment-to-moment mimicry, emulation, and other forms of behavior matching) are almost entirely unknown, and waiting to be described and explained. The present findings suggest that Piaget's (1945) description of the development of imitation (Anisfeld, 2005) and learning accounts of the 1970s (e.g., Parton, 1976) would be good places to take up these issues again.
Footnotes
Acknowledgements
I thank Diana Harwood and Jeanne Heeb for invaluable help in data collection.