
Abstract
Learning and other common psychological processes presumably evolved because they contribute to reproductive fitness, but reproductive outcomes are rarely measured in psychology experiments. We examined the effects of Pavlovian conditioning on reproductive fitness in a sperm-competition situation. Typically, two males mating with the same female in immediate succession sire similar numbers of offspring. In a study with domesticated quail (Coturnix japonica), we increased paternity success by presenting a Pavlovian signal that permitted one of two competing males to predict copulatory opportunity. Using microsatellite-based DNA fingerprinting, we found that signaled males sired 72% of the offspring when competing with control males, and this effect was independent of copulation order. In the absence of Pavlovian conditioning, rates of fertilization were not significantly different for two males that copulated with the same female. These findings demonstrate that Pavlovian conditioning contributes to reproductive fitness and suggest that individual past experience can bias genetic transmission and the evolutionary changes that result from sexual competition.
Given the prevalence of learning in the animal kingdom, one may assume that learning occurs because it is evolutionarily adaptive (Pappini, 2002). However, with few exceptions (Davey, 1989), investigators have not spelled out exactly how learning contributes to reproductive fitness. The present study examined whether learning can contribute to reproductive fitness in a particularly challenging situation—when two males compete to fertilize the gametes of a single female. To evaluate the effects of learning in sperm competition, we used one of the most familiar learning paradigms in psychology, Pavlovian conditioning.
Pavlovian conditioning is widely used because it provides a controlled method for studying associative learning at both behavioral and neurobiological levels (Turkhan, 1989). The past 100 years of Pavlovian conditioning have told researchers much about how the conditioned response develops (Pearce & Bouton, 2001), but little about why it develops. Our study focused on the “why” question. From an evolutionary perspective, the primary task of an organism is to pass along its genes to future generations. If conditioned responding evolved because it increases reproductive fitness, then subjects exposed to a conditioned stimulus (CS) should sire more offspring than subjects not exposed to the CS (Adkins-Regan & MacKillop, 2003; Hollis, Pharr, Dumas, Britton, & Field, 1997; Mahometa & Domjan, 2005). In the present study, we went beyond this simple prediction and hypothesized that if two males copulate with the same female and compete to fertilize her gametes, paternity will be biased in favor of the male that received a Pavlovian CS prior to the mating episode. The experiments were conducted with domesticated quail because they copulate readily in captivity, and because research has already provided much information on how Pavlovian conditioning modifies sexual behavior in this species (Domjan, Cusato, & Krause, 2004).
EXPERIMENT 1
In Experiment 1, contextual cues served as the CSs. For the sexual-competition test, each female was mated with 2 males in succession. For the signaled male, the test copulation was preceded by placement in a context that had been paired with sexual reinforcement. For the control male, the test copulation was preceded by placement in a control context. (The order in which the signaled and control males were allowed to copulate with the same female was counterbalanced.) The eggs were then examined for fertilization, and paternity was determined using DNA analysis. A microsatellite-based analysis was used to decrease Type II identification errors (Hanotte, Bruford, & Burke, 1992) and increase exclusion rates over those obtained with known microsatellite markers.
Method
Subjects
Fourteen female and 28 male adult domesticated quail (Coturnix japonica) served in the experiment. They were obtained from a colony maintained at the University of Texas.
Apparatus
Two experimental chambers (90 cm × 66 cm × 115 cm), each made of sealed plywood with wire mesh floors, were used. One had green side walls and a flat bottom and was located near the floor in a noisy room (green context). The other chamber was painted white, had a tilted floor, and was located in an isolated room on a table surface (tilt context). The chambers were divided in half by an opaque barrier that separated the male from the female. At floor level, the barrier had a doorway that could be opened to allow the male and female to get together to copulate.
Procedure
Training.
On each of the first 5 days, all of the birds spent 30 min in the green context and 30 min in the tilt context, for habituation. On each of the next 5 days, the females received one conditioning trial in each context. For each trial, a female was placed on one side of the experimental chamber (CS) for 1 min before the door to the male's compartment was opened, providing a 5-min copulation opportunity (which served as the unconditioned stimulus).
On Days 6 through 10, the males were also placed in the green and tilt contexts during alternate trials, but they were able to copulate in only one of the contexts. On copulation trials, a male was placed in one of the experimental chambers for 1 min and then received access to a female (unconditioned stimulus) for 5 min. After an additional 5 min, the male was returned to its home cage. On noncopulatory control trials, the male was placed in the alternate experimental chamber for 11 min and did not receive access to the female. The assignment of the green and tilt chambers as the copulation and control contexts was counterbalanced across subjects.
Copulation partners were varied so that each male-female pairing during conditioning and testing involved new partners. Following training, females were given a 14-day clean-out period without access to males, to ensure voiding of all sperm prior to testing.
Testing.
During the test trial, each female copulated with 2 males in succession. One of the males received access to the female in his signal or copulation context; the other male received access to the female in his control context. Each context exposure lasted 1 min, followed by access to the female for 5 min. Whether the signaled or the control male was permitted to copulate first was counterbalanced across subjects. The copulatory episodes were recorded with a camcorder.
Egg Collection.
Eggs were collected for 10 days starting 2 days after the test copulations. They were incubated at 99.5 °F for 5 days and were examined for embryonic development.
Blood DNA Isolation.
Blood (100 μl) was taken from all males 1 month before the experiment, suspended in lysis buffer, and stored at 4 °C. DNA was isolated using standard Qiagen (Genomic DNA Extraction Kit; Qiagen, Valencia, CA) protocol. Briefly, a 200-μl lysis-blood solution and an 800-μl phosphate-buffer saline were combined, and the resulting mixture was rinsed three times with a lysis-nanopure H20 rinse (Barnstead International, Dubuque, IA). The addition of protease, to complete the digestion, was followed by buffer washes and elution per the manufacturer's protocol. Finally, the DNA product was dissolved in Tris, ethylenediamine tetraacetic acid buffer (pH = 8.0) and stored at −20 °C.
Embryonic-Tissue DNA Isolation.
DNA fingerprinting was performed on the 39 fertilized eggs that were obtained. Embryonic tissue was removed, immediately frozen on dry ice, and stored at −20 °C. DNA was isolated using standard Qiagen (DNeasy Tissue Extraction Kit; Qiagen, Valencia, CA) protocol. Briefly, 25 mg of embryonic tissue underwent proteinase K digestion, followed by a series of buffer washes (per the manufacturer's protocol) and ethyl alcohol rinses. The final elution was stored at 4 °C. Resuspended blood- and tissue-based DNA was further digested overnight using alu1, a restriction enzyme (New England Biolabs, Ipswich, MA).
Pilot studies revealed eight digest-primer combinations with an exclusion power of 99%. Selected primers were based on specificity, independence, and similarity of annealing temperatures. An M13 sequencing probe was used on forward primers. Standard polymerase chain reaction (PCR) protocol was carried out with 1 μl of digested DNA (at a concentration of 100 ng/μl), 1 μl of M13 (forward) primer (at 1 μM), 1 μl of reverse primer (at 1.5 mM), 1 μl of 6-carboxyfluorescein fluorescent marker (at 1.5 mM), and 6 μl of PCR Supermix (Qiagen, Valencia, CA). Using an MJ Thermal Cycler PTC-200 (MJ Research, Reno, NV), we conducted 29 cycles at an initial annealing temperature of 95 °C and a resting temperature of 70 °C. PCR product was diluted (1:4) with nanopure H20 (Barnstead International, Dubuque, IA) and mixed with 8.5 μl of formalin and 0.5 μl of 6-carboxy-X-rhodamin, a passive reference dye for signal normalization, before being submitted for genetic analysis and mapping (GeneMapper software; Applied Biosystems, Foster City, CA).
Paternity Assignment.
Paternity assignment was based on similarity of markers between offspring and adult male quail, as determined by single blind coding.
Results and Discussion
Copulation in quail begins with the male grabbing the back of the female's head, mounting on her back, and making cloacal contact to transfer sperm. No significant differences in the frequency of grabs, mounts, or cloacal contacts were found between males copulating in the tilt and green chambers (all Fs < 1). Similarly, no significant differences between the signal and control contexts were observed in these behaviors, Fs(1, 27) = 3.09 (grabs), 3.11 (mounts), and 0.25 (cloacal contacts), ps > .05. However, males placed in the context that was predictive of access to a female were quicker to make cloacal contact with the female (M = 3.7 s, SE = 7.0 s) than were males placed in the control context (M = 19.1 s, SE = 3.6 s) prior to the test copulation, t(26) = 5.99, p rep = .99, η2 = .76.
Of the 78 eggs laid by the test females, 39 eggs were fertilized. Genetic analysis indicated that 28 of these (72%) were fertilized by the signaled males, and 11 were fertilized by the control males. Ten of the 14 females in the experiment produced more eggs fertilized by the signaled male than by the control male (see Fig. 1; Wilcoxon signed-ranks test, T = 13.5, p < .05). These effects were independent of the order in which the 2 males copulated with the female. Of the 39 fertilized eggs, 20 were sired by the 1st male and 19 were sired by the 2nd male.
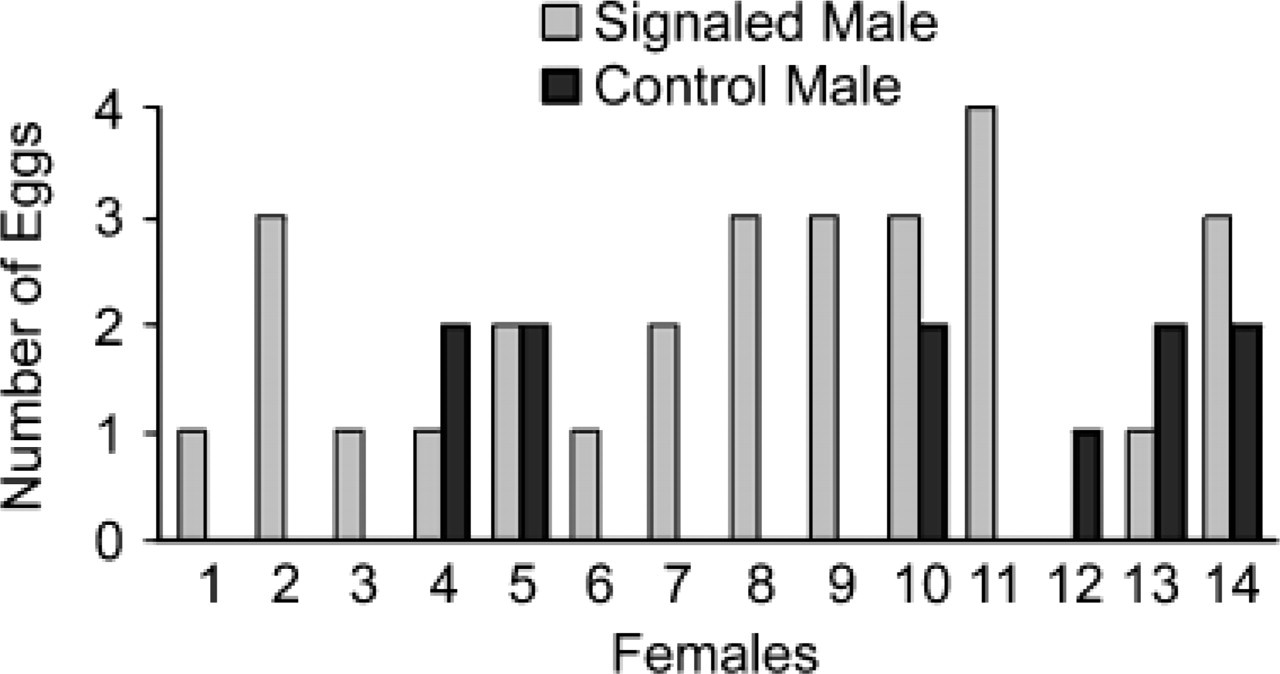
Number of eggs fertilized by the signaled versus control male, after the 2 males copulated with the same female in close succession (Experiment 1). Copulation order of the signaled and control males was counterbalanced.
The present findings show that when 2 males copulated with the same female in succession, the male that received a Pavlovian CS signaling copulatory opportunity fertilized more of the female's eggs. Thus, Pavlovian conditioning increased reproductive fitness in the context of sperm competition.
EXPERIMENT 2
Typically, when 2 males mate in quick succession with the same female, they transfer similar quantities of sperm and share equally in paternity of the offspring (Birkhead & M⊘ller, 1992). Experiment 2 was conducted to confirm this finding under the testing conditions of Experiment 1.
Method
Fourteen female and 28 male adult domesticated quail (Coturnix japonica) served in the experiment. Additional females were used to standardize sperm stores at testing, as described in the next paragraph. The test chambers were the same as in Experiment 1.
All males were placed into each context for 15 min each day over a 4-day period, for a total of eight habituation sessions. Two days prior to the copulation test, the males received a receptive nonexperimental female in their home cage and were permitted to copulate for 5 min to bring their sperm stores to the same level for the test trial. During the test trial, 2 males were permitted to copulate with the same female. One of the males was placed into a test chamber for 1 min and then given access to the female for 5 min. The female was then removed and placed into a holding chamber for 15 min before being placed into the second test chamber with the 2nd male for 5 min. Given the history of the subjects, neither of the test chambers served as a sexual conditioned stimulus. Fertilization rates and paternity were determined as in Experiment 1.
Results and Discussion
Latency to copulate did not differ between the 1st and 2nd males copulating with a given female (M = 25.25 s, SE = 5.5 s, and M = 26.8 s, SE = 7.2 s). More important, no differences in paternity rates were found between the 2 males copulating with a given female. For the 31 fertilized eggs that were obtained, the 1st male sired 17 offspring and the 2nd male sired 14. Among the 14 females that participated in Experiment 2, 9 produced more offspring from the 1st male than from the 2nd, but this outcome was not significantly different from chance (Wilcoxon signed-ranks test, T = 5.4, p > .10).
Although the fertilization rates showed that the 1st male had a slight advantage over the 2nd in both experiments, a chi-square test indicated that the distribution of fertilization rates did not differ as a function of experiment or order of copulation, χ2(1, N = 70) = 0.003, n.s. These results confirm that the testing conditions we used yielded the common finding that the 1st and 2nd male who copulate with the same female are equally likely to sire an offspring if Pavlovian signaling is not a factor.
GENERAL DISCUSSION
The present findings show that Pavlovian conditioning can act directly to increase reproductive fitness in a particularly challenging situation involving sperm competition. When two males copulated with the same female, paternity was favored for the male whose mating episode was signaled by a Pavlovian CS. Furthermore, this paternity advantage was obtained after only five conditioning trials. Thus, extensive training was not required to obtain the effect. These findings suggest that Pavlovian conditioning evolved because it increases reproductive fitness.
Pavlovian conditioning can improve an organism's effectiveness in dealing with many of the biological challenges of living, including digestion (Woods & Ramsay, 2000; Woods & Seeley, 2002), food selection (Rozin & Kalat, 1971), maternal nursing and infant suckling (Blass, 1990), aggression and territorial defense (Hollis, 1990), exposure to pain (Fanselow & Baackes, 1982; Illich & Grau, 1991), and exposure to drugs (Siegel, Baptista, Kim, McDonald, & Weise-Kelly, 2000). These areas of functioning contribute to reproductive fitness indirectly by allowing organisms to live longer and healthier lives. Prior research has also shown that Pavlovian conditioning can make a direct contribution to reproductive fitness by increasing rates of fertilization and numbers of offspring produced in copulatory interactions between a single male and a single female (Adkins-Regan & MacKillop, 2003; Hollis et al., 1997; Mahometa & Domjan, 2005).
The present study focused on competition for paternity when two males copulated with the same female. The standard definition of sperm competition suggests that sperm from more than one male battle each other to fertilize the egg of a single female during a reproductive cycle (Birkhead & M⊘ller, 1992; Birkhead & Pizzari, 2002). However, sperm competition at that level is unlikely in most avian species because an egg is available for fertilization only during a relatively brief period before the hard shell forms (18 min in Japanese quail). To overcome short fertilization windows, avian anatomy provides sperm storage tubules in which viable sperm can be stored for several days (Shugart, 1988). The stored sperm then diffuse out of the tubules randomly.
As we observed in Experiment 2, when two males inseminate the same female without Pavlovian conditioning, the paternity rates for the two males are similar because similar quantities of sperm are deposited in the storage tubules. Factors that enable a male to deposit more sperm will provide that male with a reproductive advantage.
Sperm output in some species, including humans, is influenced by the threat of sperm competition, as determined by the number of surrounding males or time away from the female (Pound, 2002; Shackelford, 2003). Studies with isolated male-female pairs of quail have shown that males can also increase total sperm transfer by increasing the frequency and efficiency of copulations (Domjan, Mahometa, & Mills, 2003; Mahometa & Domjan, 2005) or by learning to predict copulation opportunities (Adkins-Regan & MacKillop, 2003; Mahometa & Domjan, 2005).
The effect obtained in Experiment 1 was mediated entirely by the ability to predict copulatory opportunity. The males in that experiment all had similar histories of Pavlovian conditioning and had equivalent practice copulating with female quail before testing. They differed only in whether or not they were exposed to a Pavlovian signal during the sexual-competition test. Given the slow pace of spermatogenesis, it is unlikely that the Pavlovian signal increased sperm production. Rather, the Pavlovian signal probably increased sperm release. Domjan, Blesbois, and Williams (1998) previously showed that a Pavlovian contextual CS increases semen release, without changing sperm quality or concentration.
The present demonstration of increased paternity induced by Pavlovian conditioning shows that learning and individual experience can bias genetic transmission and the evolutionary changes that result from sexual competition. Given these findings, it is reasonable to begin to examine not only how evolution has shaped learning mechanisms, but also how learning can shape the trajectory of evolutionary change.
Footnotes
Acknowledgements
We wish to thank Cecil Harkey for technical assistance. This research was supported by National Institute of Mental Health Grants MH 39940 (to M.D.), T32 MH18837 (to D.C.), and MH 41770 (to D.C.).