
Abstract

Bilateral coordination of movement involves rapid exchange of information between the cerebral hemispheres. A primary structure in this interhemispheric communication is the corpus callosum (CC), which integrates the activities of the hemispheres by transferring information from sensory and higher-level cortical areas between them (Gazzaniga, Bogen, & Sperry, 1962). All CC fibers are present at birth; primary postnatal changes involve myelination of a portion of them, beginning at about 4 months after birth and continuing into young adulthood (e.g., Giedd et al., 1999). In addition, the anterior CC, which mediates interhemispheric influences on motor areas in the frontal lobes, completes a peak growth period at about 6 years of age (e.g., Thompson et al., 2000).
This peak growth period likely reflects rapid myelination of CC fibers that enable the rapid transmission needed for bimanual movements. This proposal is consistent with findings from a study in which participants controlled different components of a pen's movements with separate hands (Jeeves, Silver, & Milne, 1988). Ten- and 20-year-olds performed the task accurately, but 5- and 6-year-olds' performance was much slower and more error-prone, and resembled the performance of acallosal patients (Preilowski, 1972). Jeeves et al. attributed these performance deficits in the younger group to inefficient transfer of motor information between the cerebral hemispheres, as a result of incomplete myelination of the CC. However, this conclusion is tenuous because the task involved a complex and feedback-regulated bimanual skill.
The present study was designed to examine the influence of CC development on bimanual skill using a task that allowed more straightforward inferences regarding interhemispheric communication. The task assesses an effect referred to as bimanual cost. Specifically, adults' reaction times (RTs) to visual stimuli are slower when they use both hands to respond to a centralized stimulus (bimanual responses) than when they use only one hand (unimanual responses; Hughes & Franz, 2007). Our proposal is that when adults respond using both hands, rapid interhemispheric signals are sent from each hemisphere (perhaps to inform the other that a response is about to occur). These signals couple normal bimanual responses, but also activate inhibitory networks in each hemisphere, slowing the responses. In the unimanual control situation, the same interhemispheric signals are transmitted, but only from the responding hemisphere. Thus, the influence of these signals on inhibitory networks is not reflected in movement. Consequently, bimanual responses are slower than unimanual responses (Hughes & Franz, 2007).
We hypothesized that bimanual cost begins to emerge at about 6 years of age, when the peak growth period of the anterior CC is completed. However, bimanual cost could emerge because of development of inhibitory networks alone. To evaluate the latter possibility, we tested people with complete callosal agenesis. Absence of bimanual cost in the acallosal group would implicate the CC and be inconsistent with the view that bimanual cost is due solely to cortical inhibitory networks.
METHOD
Participants were recruited from New Zealand schools and from a nationwide search for individuals with callosal agenesis (acallosal group). The 100 neurologically normal children were recruited according to age (n = 25 in each of four age groups: 4–5 years, 6–7 years, 8–9 years, and 10–11 years). At the time of testing, the 3 female acallosal participants were 11, 17, and 36 years old, respectively; they were diagnosed by magnetic resonance imaging and neurological exam as having complete callosal agenesis. Each acallosal participant showed minor cognitive deficits but no obvious motor problems that would influence her ability to perform our bimanual task. Informed consent was obtained from all participants and children's parents.
The task stimuli were presented on a computer monitor. Each trial began with a 1- × 1-cm fixation point that was displayed for 500 to 1,000 ms. Next, a dot measuring 1.5 cm in diameter appeared in the center of the screen. Participants were instructed to respond to the dot by pressing one or two keys (see the next paragraph) as quickly as possible. The dot remained on the screen for 1,200 ms or until a response was made, whichever came first. The intertrial interval was 1,000 ms. Participants received a verbal warning if their responses occurred prior to stimulus onset or were unusually slow.
Participants used their index fingers to respond. Following 36 practice trials, separate 36-trial blocks tested performance in left-hand unimanual, right-hand unimanual, and bimanual conditions. Each condition was tested in two blocks, for a total of 216 trials per participant. The six test blocks were presented in random order. Participants were highly motivated, as they were told that at the end of testing, they would receive rewards (stickers or gift certificates) for fast performance.
RESULTS AND DISCUSSION
After outlying RTs (less than 100 ms and greater than 700 ms, 5% of the data) were eliminated for all participants, the RTs (see Table 1) for neurologically normal control participants were analyzed with an analysis of variance in which group was a between-subjects factor and condition (unimanual vs. bimanual) and hand were within-subjects factors. RTs declined with age, F(3, 96) = 31.11, p < .001. Bimanual RTs were significantly slower than unimanual RTs, F(1, 96) = 13.72, p rep = .62, and responses of the nondominant hand were slower than responses of the dominant hand, F(1, 96) = 17.07, p rep = .60.
Primary Variables of Interest for Each Age Group and for Acallosal Individuals
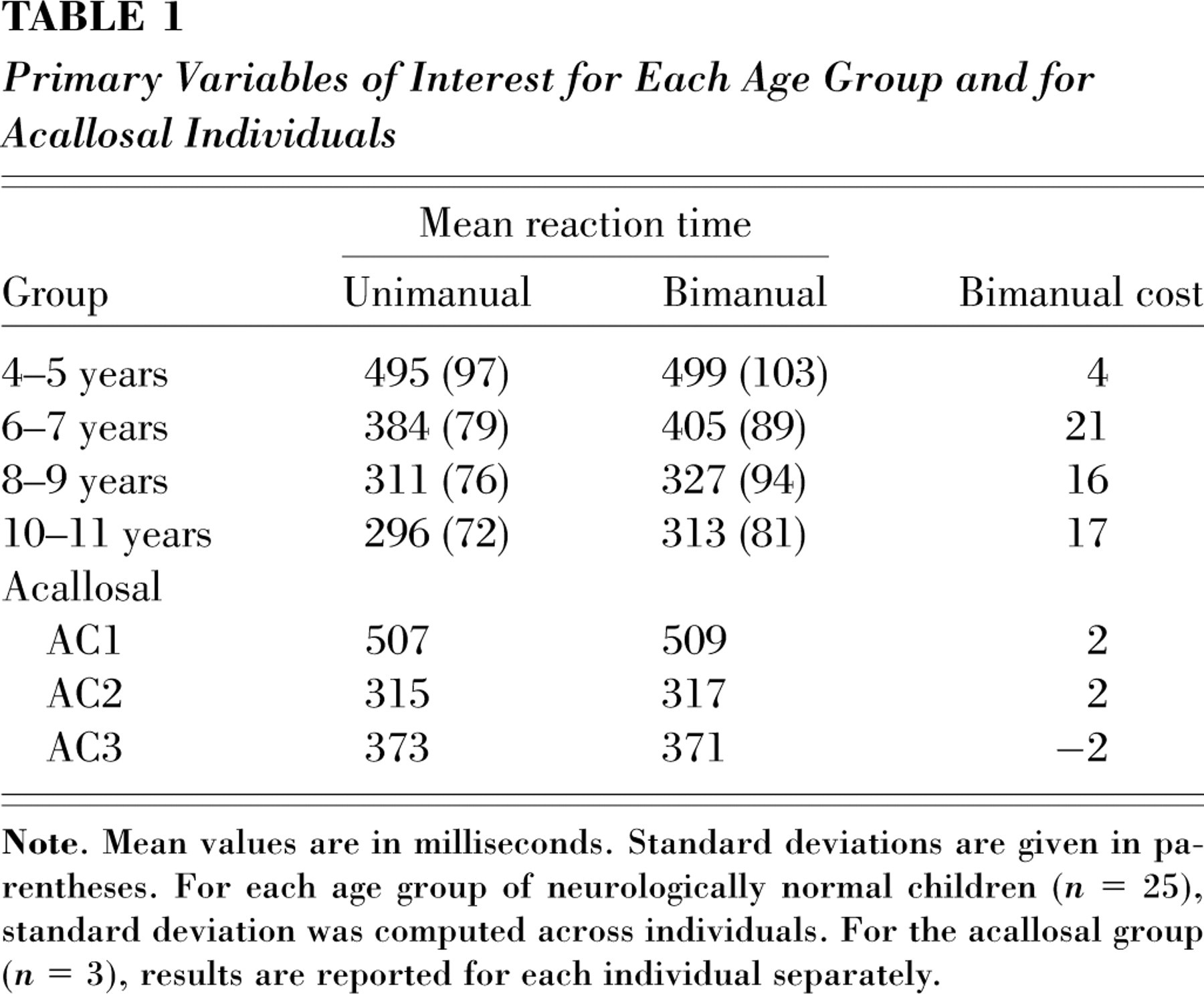
In this group of participants, the absolute value of the RT difference between hands in the bimanual condition (computed across trials and then averaged for each participant to yield a measure of bimanual synchrony) decreased significantly with age, F(3, 96) = 5.29, p = .002; a gradual decrease in this bimanual RT difference is hypothesized to continue into adulthood, because of the continued and prolonged maturation of CC myelination processes (see the introduction). More important, as we hypothesized, bimanual cost emerged at about 6 years of age. The difference between bimanual and unimanual RTs was significant at ages 6 through 11 years, ts ≥ 2.16, p reps = .70–.72, but not at ages 4 to 5 (t < 1).
Mean RT of the acallosal group was similar to the children's mean RT; more important, no acallosal participant exhibited bimanual cost (see Table 1). (Treating trial as a random factor, within-subjects tests on unimanual vs. bimanual trials yielded Fs less than 1.)
In sum, this study demonstrates that bimanual cost, which is thought to reflect a rudimentary form of interhemispheric communication necessary for bimanual movements, emerges in association with completion of a peak growth period in the CC, the primary cortical commissure that mediates this communication. This effect need not be limited to the bimanual task used in this study, given that similar forms of callosally mediated interhemispheric communication underlie many other cognitive and motor tasks.
Footnotes
Acknowledgements
We gratefully acknowledge support from a Todd Foundation Centenary Grant awarded to E.A.F. We thank Harlene Hayne for helpful comments.