
Abstract
A growing literature suggests that the degree to which distracting information can be ignored depends on the perceptual load of the task, or the extent to which the task exhausts perceptual capacity. However, there is currently no a priori definition of what constitutes high or low perceptual load. We propose that interactions among cells in visual cortex that represent nearby stimuli determine the perceptual load of a task, and that manipulations designed to modulate these competitive spatial interactions should modulate distractor processing. We found that either spatially separating the task-relevant items in a display or placing the target and nontargets in different visual fields increased interference from a distractor that was to be ignored. These data are consistent with the idea that the ability to ignore such distracting information results in part from the need to actively resolve competitive interactions in visual cortex, and is not the consequence of an exhausted capacity per se.
The extent to which distracting information can be ignored has been debated for decades, with some researchers suggesting that ignored information is filtered early in the system so that it is essentially blocked from perception (i.e., early selection; e.g., Broadbent, 1958), and others suggesting that the unattended information is processed to a fairly high level and thus can still influence behavior (i.e., late selection; Deutsch & Deutsch, 1963). A more recent theory, however, proposes that attentional selection can be either early or late, depending on the perceptual demands, or perceptual load, of the relevant task (Lavie, 1995). When the perceptual load of the task is high, few resources are available for processing the distractor, and the distractor is excluded early in processing. However, when the perceptual load of the task is low and consequently does not exhaust perceptual capacity, the remaining resources spill over on to the distractors, and processing proceeds past the early stages, resulting in the need for late selection. Although this theory has received considerable support from behavioral, electrophysiological, and neuroimaging experiments (Beck & Lavie, 2005; Handy, Soltani, & Mangun, 2001; Lavie, 1995; Lavie & Cox, 1997; Rees, Frith, & Lavie, 1997), it remains unsatisfying for two reasons. First, a clear definition of perceptual load has yet to emerge. Second, the concept of an exhausted capacity is difficult to reconcile with mechanisms in the brain. We propose a definition of perceptual load that is neurally plausible and also generates two novel predictions that are supported by the data.
In a number of phenomena, the detectability, discriminability, or neural representation of a target stimulus is diminished by the presence of nearby items (Andriessen & Bouma, 1976; Intriligator & Cavanagh, 2001; Kastner, de Weerd, Desimone, & Ungerleider, 1998; Petrov, Carandini, & McKee, 2005; Polat & Sagi, 1993; Reynolds, Chelazzi, & Desimone, 1999), and many of these phenomena are thought to originate in early to intermediate areas in visual cortex (e.g., striate and extrastriate cortex) as a result of interactions among cells that represent stimuli near one another in space (i.e., local spatial interactions; Bair, Cavanaugh, & Movshon, 2003; Blakemore & Tobin, 1972; DeAngelis, Robson, Ohzawa, & Freeman, 1992; Kastner et al., 1998; Polat, Mizobe, Pettet, Kasamatsu, & Norcia, 1998; Reynolds et al., 1999; Reynolds & Desimone, 2003). For example, stimuli presented simultaneously in the visual field are not processed independently, but instead interact in a mutually suppressive way that suggests a competition for neural representation (Beck & Kastner, 2005; Desimone & Duncan, 1995; Kastner et al., 1998; Reynolds et al., 1999). Moreover, evidence suggests that selective attention can bias this competition in favor of the attended stimulus; that is, competitive interactions among stimuli are resolved when the subject attends to a single item (Kastner et al., 1998; Reynolds et al., 1999).
We propose that these competitive interactions in visual cortex and the resulting biasing mechanisms needed to resolve the competition in favor of the target may be the neural mechanisms underlying perceptual load. In particular, we suggest that the degree to which items within a display compete for representation in visual cortex determines the strength of the biasing mechanisms necessary to resolve the competition; the strength of these biasing mechanisms, in turn, will determine the degree to which unattended information is processed. Tasks that induce strong competitive interactions among potential target items should require a strong top-down bias to overcome the competition and select the target, and can thus be characterized as having a high perceptual load. In contrast, tasks in which minimal competition is induced among the potential target items should require little top-down bias for the target to be selected and can thus be characterized as having a low perceptual load.
Such a characterization of perceptual load fits well with the two most common manipulations of load in the literature, increments in the number of items in a display (i.e., set size) and similarity of target and nontargets. Increments in set size are said to increase the perceptual demands of the task, and have been shown to result in diminished interference from a distractor, as predicted by perceptual-load theory (Lavie & Cox, 1997). However, increments in the number of items in a display also increase the likelihood of competitive interactions in visual cortex (e.g., the more items, the more likely that their representations interact in visual cortex), thus resulting in a greater need for top-down biasing to identify the target.
In the second common manipulation of perceptual load, set size is kept constant, and the similarity of the target and nontarget items is manipulated (Beck & Lavie, 2005; Lavie & Cox, 1997). In the low-load condition, the target and nontargets are perceptually dissimilar, which makes the target salient and easy to detect (e.g., the target is an X or N embedded in a field of Os). Competitive interactions among items would also explain why such displays would be considered low load; competitive interactions among stimuli in visual cortex are reduced when one of the items is made salient (Beck & Kastner, 2005; Reynolds & Desimone, 2003). In the high-load condition, however, the same target is embedded among other letters that share many features with the target (e.g., Y, Z, and H when the targets are X and N). Therefore, the target does not benefit from saliency, and a strong top-down bias is needed to bias the competition in favor of the target.
This conception of perceptual load as determined by competitive interactions in visual cortex not only can account for previous results in the perceptual-load literature, but also suggests two novel and previously untested perceptual manipulations that should modulate distractor processing. First, because these competitive interactions are thought to occur locally, the distance between potential target items should affect the degree to which the interactions occur. Indeed, the distance between stimuli has been shown to modulate competitive interactions in visual cortex (Bles, Schwarzbach, de Weerd, Goebel, & Jasma, 2006; Kastner et al., 2001). Thus, if perceptual load is determined by local interactions among stimuli in visual cortex, as we have argued, then manipulations of display density should modulate the degree to which subjects are able to ignore a distractor. Specifically, as the distance between the target and nontarget stimuli decreases, the bias needed to resolve the competition should increase, resulting in diminished processing of the distractor. We tested this prediction in Experiment 1.
Second, local spatial interactions have primarily been seen in early to intermediate levels of visual cortex (i.e., V1–V4), in which the representation of a visual hemifield (right or left) is confined to the contralateral hemisphere. If perceptual load is determined by local interactions in these areas, then placing multiple items within the same hemifield should result in greater perceptual load and less distractor processing, compared with placing the target item alone within a hemifield, so that it is not subject to the same local interactions. We tested this prediction in Experiment 2.
EXPERIMENT 1
In Experiment 1, we varied display density, keeping set size, target and nontarget similarity, and distance between the target and distractor constant across density conditions. On the basis of what has previously been accepted as a perceptual-load manipulation, there would be no reason to expect that the density of the search display would affect distractor processing. It is reasonable to suspect, however, that finding a target would be more difficult in higher-density displays, so high density might be considered high perceptual load. However, we emphasize that our manipulation was derived in a principled way from our theory regarding local interactions; unlike many previous perceptual-load manipulations, it was not based on an appeal to task difficulty. Such appeals are not wholly satisfying, as they beg the question of why the task is more difficult in the first place.
Method
Sixteen volunteers recruited from the University of Illinois, Urbana-Champaign, participated for course credit or monetary reward. On each trial, a CRT computer monitor displayed an array of four letters arranged in an arc (radius of 3.7° of visual angle) around a distractor centered at fixation (see Fig. 1a). The density of the display was manipulated by either placing small circles (0.2° × 0.2°) between the letters, resulting in a center-to-center distance of 2.57° between adjacent letters (low density), or placing the four letters immediately adjacent to each other, separated by 1.29°, with the circles located at the ends of the arc of letters (high density). Participants were asked to search the array of letters for the target and ignore the distracting letter at fixation. The distractor was either compatible with the target (e.g., Z when the target was Z), incompatible with the target (e.g., Z when the target was X), or neutral with respect to the target (R when the target was either X or Z). Each trial began with a fixation cross that appeared in the center of the screen for 500 ms. The search array was presented for 100 ms immediately after the fixation display. Trials terminated following a response, or after 2 s if there was no response. At a viewing distance of 40 cm (stabilized by a chin rest), each letter in the search array subtended 0.7° × 1° of visual angle, and the distractor, which was fixated, subtended 0.8° × 1.1°.
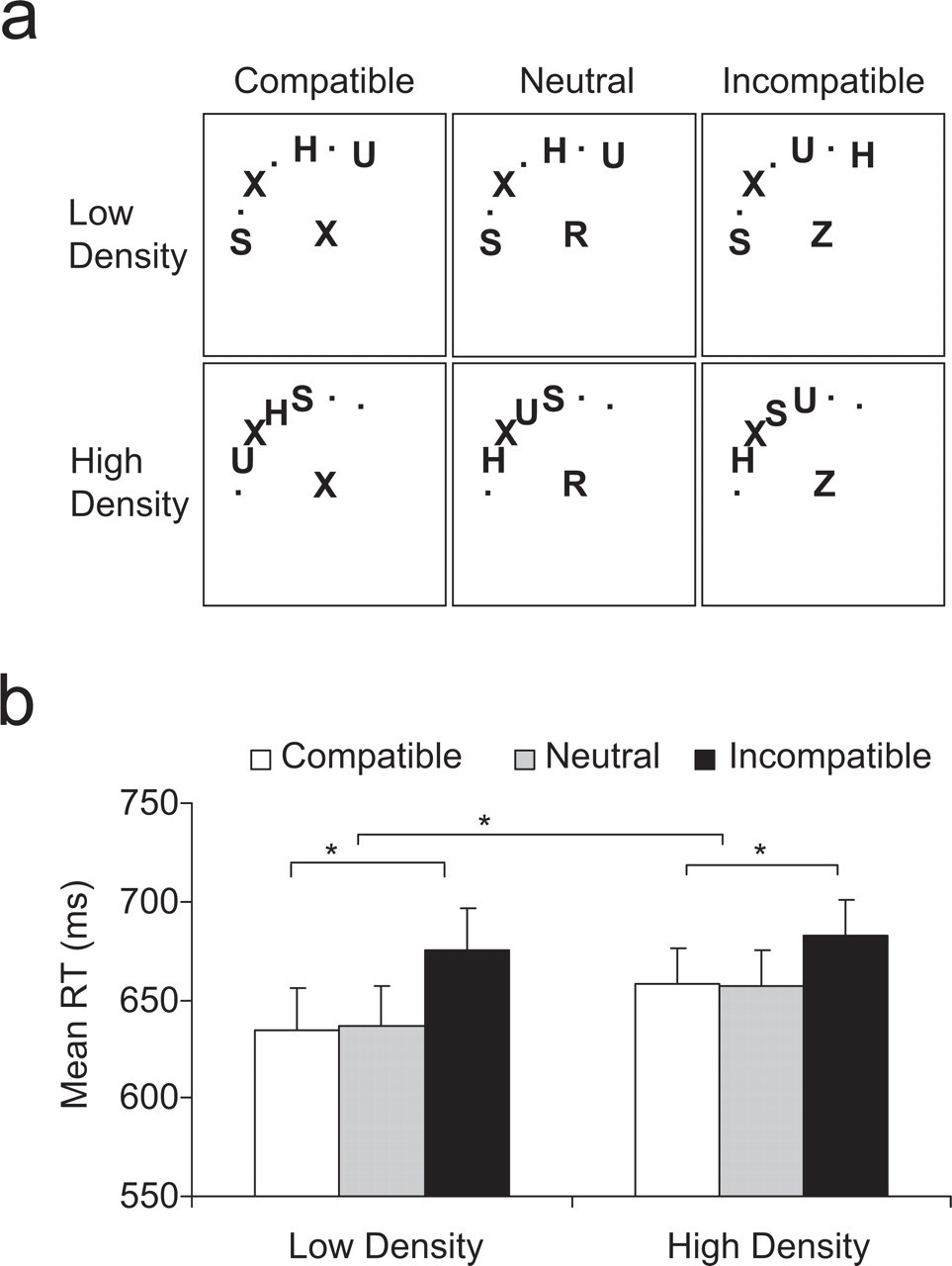
Illustration of the displays used in Experiment 1 (a) and results of the experiment (b). In these sample displays, the stimuli appear in the upper left quadrant, but in the experiment they appeared equally often in all four quadrants. Density of the letters in the search array was either high or low, and the central distractor was compatible with the target (X in the examples shown here), incompatible with the target, or neutral with respect to the target. The graph shows mean reaction times (RTs) for correct responses as a function of density of the search array and compatibility of the distractor. Error bars denote standard errors of the means, and asterisks denote statistically significant comparisons (p < .05).
Targets were always in one of the two inner positions of the array, and they appeared equally often in these two positions. The quadrant in which the search array was presented was determined randomly on each trial; across trials, the array appeared equally often in all four quadrants. Participants completed a practice block of 48 trials and six experimental blocks of 96 trials in each density condition. Within a block, the distractor was compatible with the target on one third of the trials, incompatible with the target on one third of the trials, and neutral with respect to the target on one third of the trials. Low-density and high-density blocks were presented in alternation; half the participants started with a low-density block, and half began with a high-density block. Participants were instructed to indicate which of the target letters (X or Z) was present in each search array by pressing one of two keys on the keyboard. The left-right mapping of the keys to letters was counterbalanced across participants. Participants were told to answer as quickly as possible without sacrificing accuracy. They completed 24 trials for each combination of display density, compatibility, target position, and array location.
Results and Discussion
Mean reaction times (RTs) and error rates were computed for each participant for each combination of compatibility condition (compatible, incompatible, neutral) and density condition (low, high). These RTs and error rates were entered into separate two-way within-subjects analyses of variance. Trials that were not answered correctly were excluded from the RT analyses. RTs more than 2.5 standard deviations above or below the mean of each cell in the experimental design, computed separately for each subject, were considered outliers and were also excluded from the RT analyses. In all, 3% of trials with correct responses were excluded for this reason.
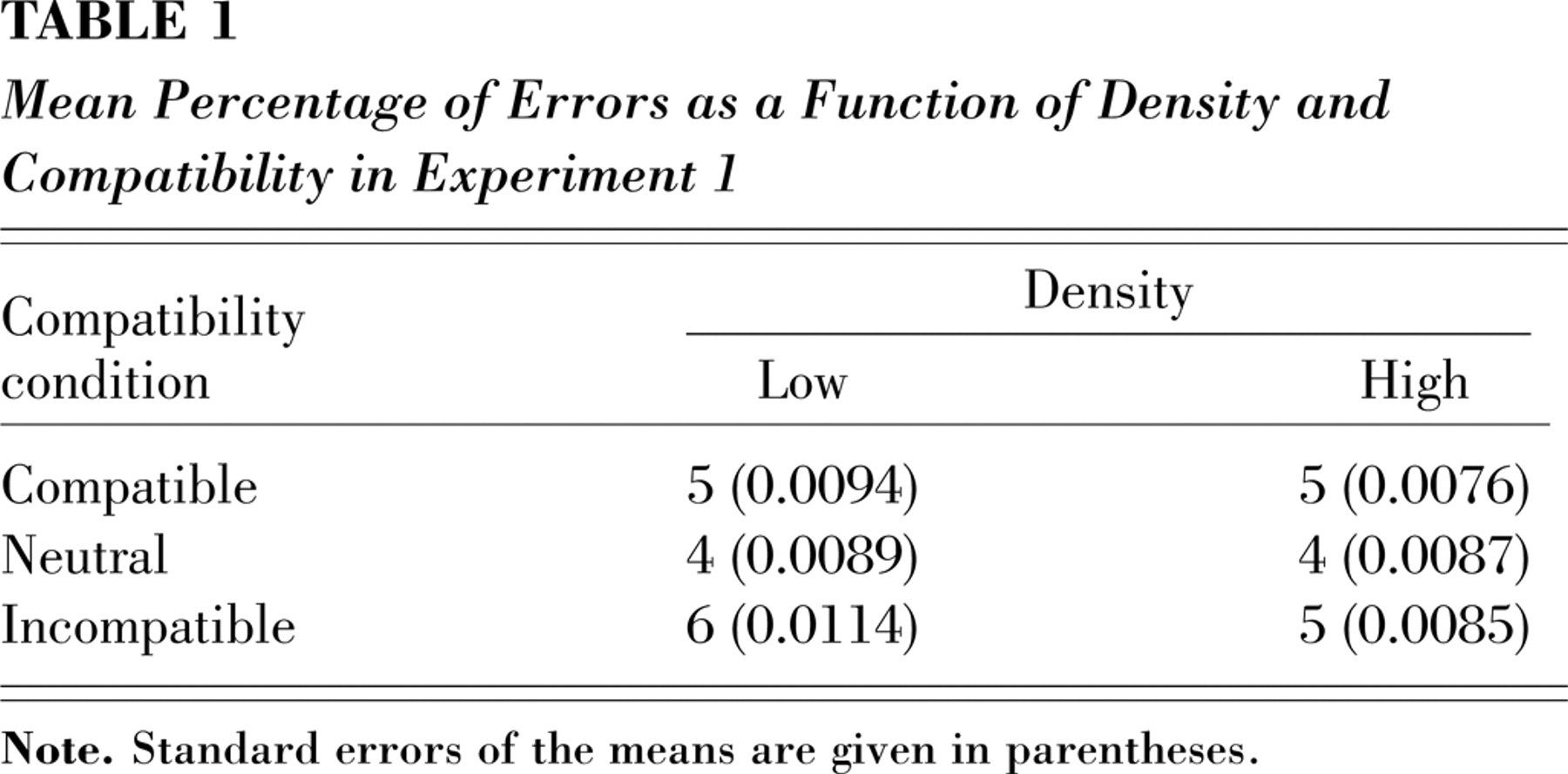
The RT analysis showed a main effect of density, F(1, 15) = 8.48, p < .01, such that RTs to high-density displays were slower than RTs to low-density displays (666 ms and 648 ms, respectively). The degree to which subjects processed a distractor placed at fixation was assessed by comparing RTs on compatible and incompatible trials. There was a main effect of compatibility, F(2, 30) = 28.54, p < .0001, such that mean RTs were significantly slower in the presence of an incompatible distractor (679 ms) than in the presence of a compatible distractor (647 ms), t(15) = 5.09, p < .0001, or a neutral distractor (646 ms), t(15) = 7.55, p < .0001; this result indicates that the distractor was in fact processed despite instructions to ignore it because it was irrelevant to the task. However, this effect of the distractor's compatibility was modulated by the density manipulation, F(2, 30) = 4.32, p < .022, such that the high-density displays produced much smaller distractor effects than the low-density displays (Fig. 1b). The effect of distractor compatibility was significant for both low-density displays (40 ms), t(15) = 5.42, p < .0001 (two-tailed paired t test), and high-density displays (24 ms), t(15) = 4.21, p < .001, but were significantly reduced in the high-density condition, relative to the low-density condition, t(15) = 3.86, p < .002. The error rates also showed a larger compatibility effect for low-density than for high-density displays (see Table 1), but only the main effect of compatibility was significant, F(2, 30) = 6.26, p < .005. Thus, in accordance with our hypothesis that local competitive interactions in visual cortex underlie the effects of perceptual load, more closely spaced letters resulted in less interference from a distractor than did less closely spaced letters.
Mean Percentage of Errors as a Function of Density and Compatibility in Experiment 1
EXPERIMENT 2
In Experiment 2, we manipulated whether or not the nontargets (i.e., potentially competing letters) fell within the same hemifield as the target by placing the peripheral search display (the target and three nontarget letters arranged in a diamond configuration) such that only one of the four items crossed the vertical midline (see Fig. 2a). In half the trials, the target was the item that crossed the midline, and it therefore appeared alone in the hemifield; in the other half of the trials, the target shared a hemifield with two nontarget items.
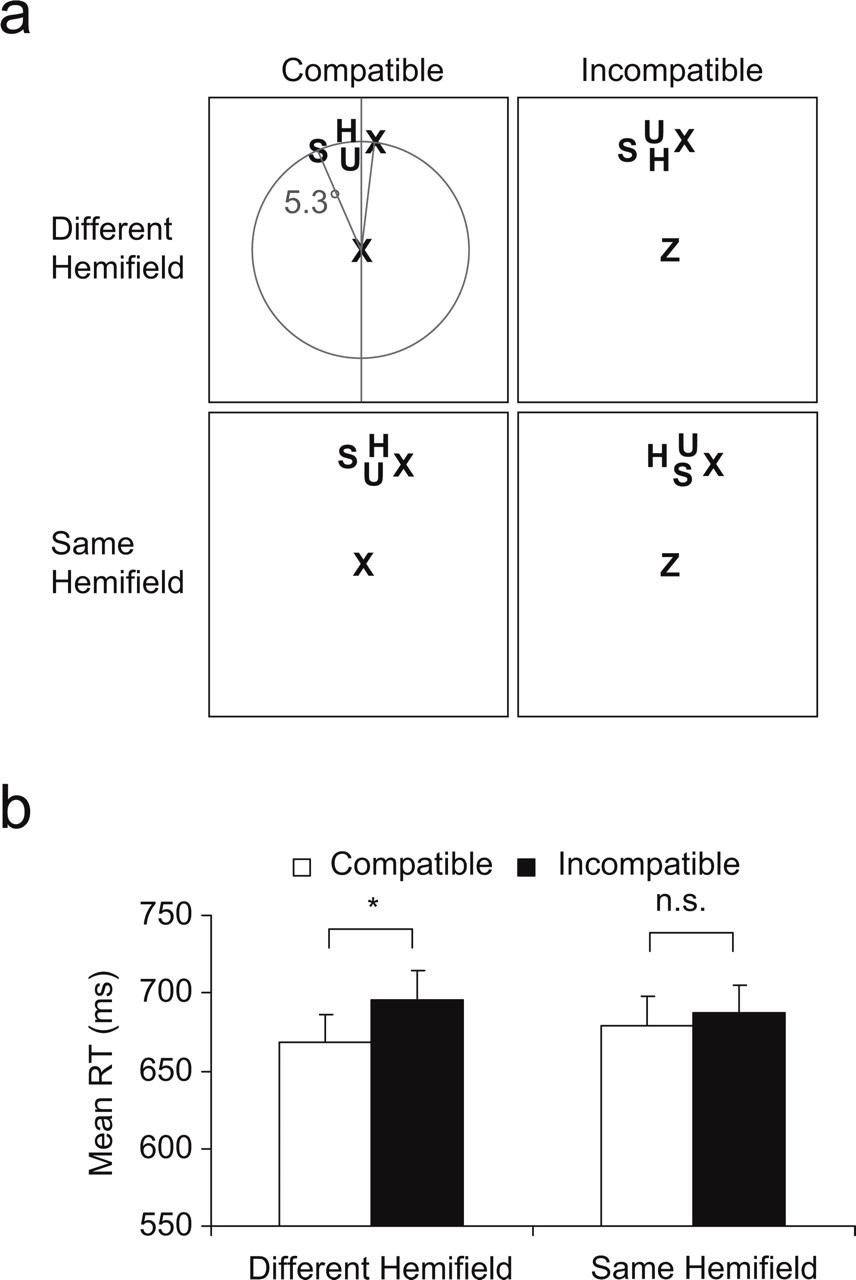
Illustration of the displays used in Experiment 2 (a) and results of the experiment (b). In this experiment, the target appeared in the same hemifield as two nontargets on some trials and was alone in the hemifield on other trials (examples of targets in the upper right hemifield are shown here, but targets appeared equally often in the left hemifield and equally often in the upper and lower visual fields). The circle, lines, and indication of distance in the upper left panel were not shown in the actual experiment. The graph shows mean reaction times (RTs) for correct responses as a function of hemifield condition and compatibility of the distractor. Error bars denote standard errors of the means, and the asterisk denotes a statistically significant comparison (p < .05).
Method
Twenty volunteers recruited from the University of Illinois, Urbana-Champaign, participated in this study for course credit or monetary reward. Their task was to identify a target letter (X or Z) in a peripheral search display arranged in a diamond configuration. The sides of the diamond subtended 1.57° in length, and the two possible target locations (right and left vertices of the diamond) were located 5.3° from fixation (see Fig 2a). The minor and major axes of the diamond subtended 1.40° and 2.80°, respectively. The closest edge of the target was 0.28° or 1.76° from the midline, and nontargets were 0.28°, 0.50°, and 1.76° from the midline. This diamond configuration was mirrored across the vertical meridian both in the upper (see Fig 2a) and in the lower visual fields, resulting in four possible diamond locations. Distractors were equally likely to be compatible or incompatible (neutral trials were not included in this experiment). As in the first experiment, the distractor was located at fixation, and thus represented in both hemifields. Participants completed a practice block of 32 trials and 6 experimental blocks of 64 trials each. There were 24 trials for each combination of diamond location, target location, and compatibility, and these trials were randomly intermixed within blocks. Because local interactions should occur within and not between visual hemifields, we predicted greater competition among target and nontarget letters, and thus reduced distractor processing, when the target and nontargets appeared in the same hemifield than when they appeared in different hemifields.
Results and Discussion
As in Experiment 1, mean RTs and error rates were computed for each participant for each combination of compatibility condition (compatible, incompatible) and hemifield condition (target and nontargets in same hemifield, target and nontargets in different hemifields). These RTs and error rates were submitted to separate two-way within-subjects analyses of variance. Using the same criterion as in Experiment 1, we excluded 2.8% of trials with correct responses because the RTs were outliers.
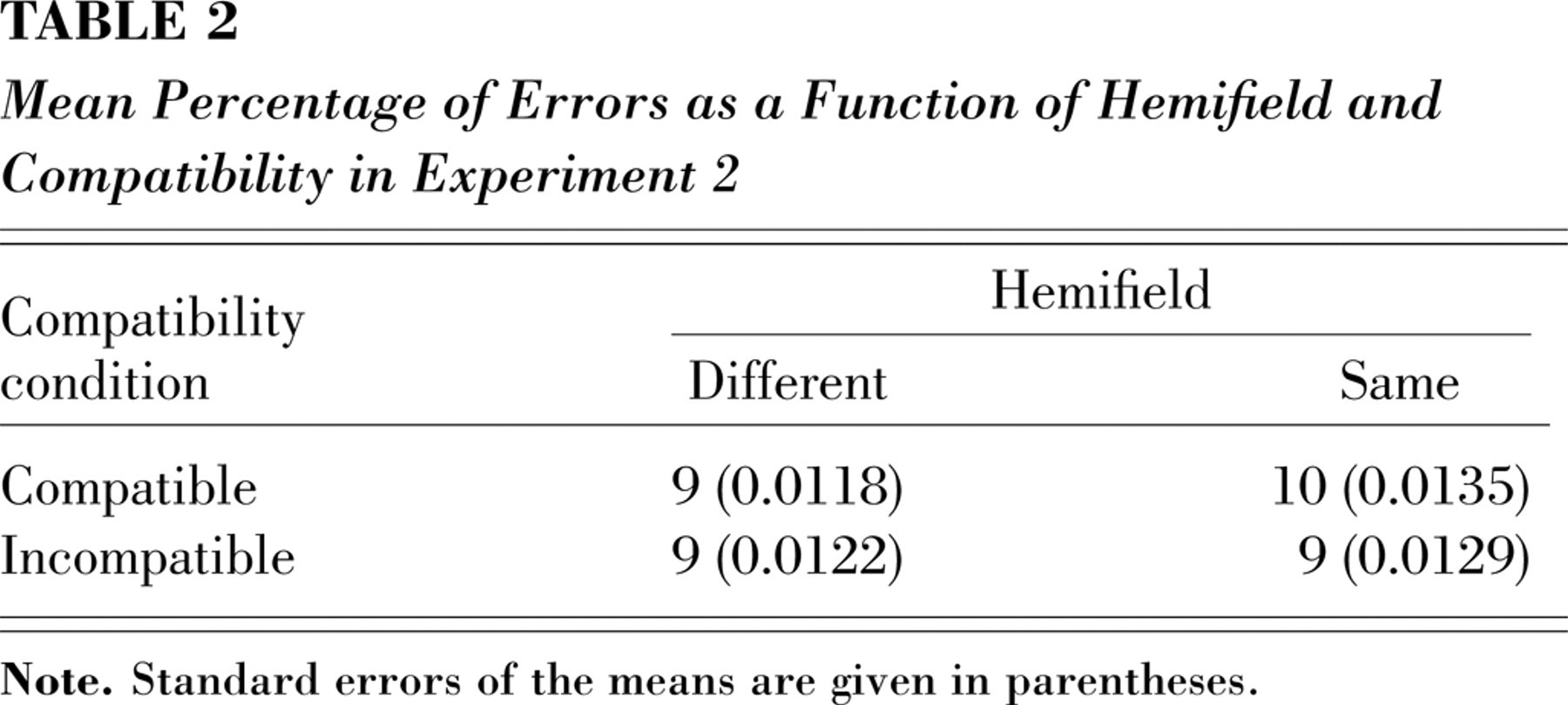
The RT analysis showed a main effect of compatibility, F(1, 19) = 13.27, p < .002; RTs were slower on incompatible trials than on compatible trials (691 ms and 674 ms, respectively). However, as predicted, trials in which the target appeared by itself in a hemifield produced greater distractor effects than trials in which the target appeared in the same hemifield with nontargets, F(1, 19) = 6.97, p < .016 (see Fig 2b; Table 2 presents the error rates). The effect of distractor compatibility was significant when the target was presented alone in the hemifield (27 ms), t(19) = 4.75, p < .0001 (two-tailed paired t test), but not when the target and nontargets appeared in the same hemifield (8 ms; p > .2). There were no significant effects in the analysis of error rates.
Mean Percentage of Errors as a Function of Hemifield and Compatibility in Experiment 2
Because observers are generally unaware of the division between their left and right hemifields, from the subjects' perspective the task was the same throughout the experiment, yet the hemifield manipulation had a dramatic effect on distractor effects. These results further support our proposal that perceptual load is determined by local interactions in early to intermediate levels of visual cortex, because it is in these areas that the right and left visual fields are processed in separate hemispheres.
These results not only support the hypothesis that local spatial interactions among the target and nontargets dictate the extent to which a distractor can be ignored, but also suggest that the attentional bias is determined independently in the two visual cortices. Despite the fact that the target and nontargets formed a single perceptual group, the degree to which the distractor was processed depended on the location of the target within that group. By our theory, a strong bias was produced when the target appeared in the same hemifield as the nontargets, whereas at most a weak bias was produced when the target and nontargets appeared in different hemifields.
Our results are in accordance with evidence of parallel attentional processing by the two hemispheres in split-brain patients (Luck, Hillyard, Mangun, & Gazzaniga, 1989) and in neurologically intact subjects (Alvarez & Cavanagh, 2005; Scalf, Banich, Kramer, Narechania, & Simon, 2007). Moreover, our theory suggests why attentional biases may be determined independently in the two hemispheres, despite the fact that information in the two hemispheres is integrated across visual fields in higher visual areas. If the task is such that the target must be resolved in early to intermediate levels of visual cortex (i.e., local spatial interactions in those areas must be counteracted in order to resolve the target), then different top-down biases may develop to resolve competition in the two visual cortices.
GENERAL DISCUSSION
The data presented here suggest that competition for representation in visual cortex may underlie perceptual load, and thus determine the degree to which unattended information is processed. Stimuli that should produce greater competition in visual cortex resulted in distractor effects akin to those found with high perceptual load. Moreover, these experiments implicate local interactions in early to intermediate levels of visual cortex, in which the representation of the visual fields is spatiotopic (Experiment 1), and in which the right and left visual fields are represented in separate hemispheres (Experiment 2). Higher visual areas, such as inferotemporal cortex, have large receptive fields that encompass both visual hemifields and therefore are less probable candidates for the effects we observed (Gross, Rocha-Miranda, & Bender, 1972).
We should note that some researchers have manipulated perceptual load in ways such that the observed effects cannot be explained by appealing to local competitive interactions. These manipulations did not involve changes to the stimuli, but rather involved changes to the task itself (e.g., discriminate colors vs. discriminate color and orientation conjunctions; Handy et al., 2001; Lavie, 1995). Thus, we are not suggesting that the only way to manipulate the perceptual load of a task is to manipulate local competitive interactions in visual cortex. However, it is possible that the same biasing mechanisms needed to resolve local competitive interactions are involved in binding features, such as orientation and color, into a single object in a conjunction task (Duncan, Humphreys, & Ward, 1997; Schoenfeld et al., 2003). In other words, the reduction in distractor effects found in conjunction tasks relative to feature tasks may reflect the need for stronger top-down biasing in conjunction tasks.
Finally, these data and our theory suggest a neurally plausible description of perceptual resources. Many researchers have taken issue with the concept of “a resource,” as it is not clear what this means in terms of the brain. We suggest that one so-called limited resource that stimuli are competing for is the response of local regions of visual cortex. However, although the concept of “exhausting” (or failing to exhaust) this limited resource provides a convenient description of the data (Lavie, 1995), we are proposing a very different conceptualization of perceptual load based on known effects in visual cortex. In situations of high perceptual load, it is not the case that perceptual resources are exhausted per se; rather, in order to overcome local competitive interactions, a strong bias is necessary to isolate and improve the representation of the target.
Footnotes
Acknowledgements
This research was supported by a fellowship from the Ministerio de Educación y Ciencia of Spain to A.T. (AP 5121-2003) and by a grant from the National Institute of Mental Health to D.M.B. (R03 MH082012 A). We thank Arthur F. Kramer, Daniel Simons, Alejandro Lleras, and James L. Beck for comments on an earlier draft of this manuscript.