
Abstract
Proponents of a geometric module claim that human adults accomplish spatial reorientation in a fundamentally different way than young children and non-human animals do. However, reporting two experiments that used a conflict paradigm, this article shows striking similarities between human adults and young children, as well as nonhuman animals. Specifically, Experiment 1 demonstrates that adults favor geometric information in a small room and rely on features in a larger room, whereas Experiment 2 demonstrates that experience in a larger room produces dominance of features over geometric cues in a small room—the first human case of reliance on features that contradict geometric information. Thus, use of features during reorientation depends on the size of the environment and learning history. These results clearly undermine the modularity claim and the view that feature use during reorientation is purely associative, and we discuss the findings within an adaptive-combination view, according to which a weighting system determines use of feature or geometric cues during reorientation.
In recent years, research on the reorientation abilities of mobile organisms has proceeded at a fast pace, in part because of the debate over the existence of a geometric module. Cheng (1986) made the fascinating discovery that, following disorientation, animals reorient within a fully enclosed rectangular space by using the metric information given by the lengths and angles of the walls that form the shape of the surrounding environment, while largely ignoring feature cues, such as a colored wall or landmarks. This reorientation procedure yielded similar results for rats (Cheng, 1986) and very young children (Hermer & Spelke, 1994, 1996) in that they searched for a target in either the correct corner or the rotationally opposite corner (with respect to the geometry of the enclosure). In both cases, searches for a target were evenly distributed between the geometrically equivalent corners regardless of whether or not the space contained features. In contrast, human adults easily combine geometric and feature cues in order to search the correct corner and are thus highly accurate in the standard reorientation task (Hermer & Spelke, 1994; Hermer-Vazquez, Spelke, & Katsnelson, 1999; Ratliff & Newcombe, 2008).1
Additional evidence, however, has led researchers to question the existence of a geometrically driven reorientation system. When a feature is more distal, stable, and useful than in the original studies, and when geometric spaces are not small, young children (Learmonth, Nadel, & Newcombe, 2002; Learmonth, Newcombe, & Huttenlocher, 2001; Learmonth, Newcombe, Sheridan, & Jones, in press; Newcombe, Jones, Shallcross, & Ratliff, 2006) and various nonhuman animals, including chickens (Vallortigara, Zanforlin, & Pasti, 1990), pigeons (Kelly, Spetch, & Heth, 1998), monkeys (Gouteux, Thinus-Blanc, & Vauclair, 2001), and even fish (Sovrano, Bisazza, & Vallortigara, 2002), show increased use of features during reorientation. Having prior experience using features also increases feature use during reorientation in a small rectangular space, as has been found with wild-caught mountain chickadees (Gray, Bloomfield, Ferrey, Spetch, & Sturdy, 2005), fish that have been raised in circular tanks (Brown, Spetch, & Hurd, 2007), and young children with brief exposure to feature training (Twyman, Friedman, & Spetch, 2007) or experience using features in larger spaces (Learmonth et al., in press).
The adaptive-combination model (Newcombe & Huttenlocher, 2006; Newcombe & Ratliff, 2007) proposes that use of features and geometry depends on the relative weight associated with each cue. These weights are determined by factors such as the reliability and validity of the cue, the salience and strength of encoding of the cue, and prior experience using the cue. For example, landmarks are more likely to be used during navigation when they are more distal (Nadel & Hupbach, 2006). Because objects that are visible from further away tend to be large, immovable objects, their distance from the observer increases their cue validity and salience, giving added weight to features.
Applying this model specifically to the reorientation paradigm, we propose that search behavior is guided by a weighting mechanism combining both geometric and feature information weighted by the variance associated with each source. The weights associated with the cues depend on the strength of each cue at encoding; the relative salience, reliability, and validity of the cues; and which cue has been used most often in the past. A weight is initially established for each kind of information (geometric vs. feature) during encoding, and these weights are then combined following Bayesian rules (e.g., greater unreliability is associated with greater variance; Newcombe & Ratliff, 2007). We propose that when action is required in the space again, either for basic navigation or following disorientation, mobile organisms map the location of cues from their previous encoding onto the current space, making any necessary adjustments given changes in cue salience, cue validity, or prior experience. For instance, if a particular kind of feature changes frequently (e.g., flowers that bloom and die), that feature would not be used in subsequent reorientation efforts.
To account for developmental changes in reorientation, proponents of the geometric module argue that productive use of spatial language overcomes the encapsulation of geometric information (Hermer-Vazquez et al., 1999). In this view, the ability to correctly use terms such as “left” and “right” allows children to flexibly conjoin geometric and feature cues. Indeed, the age at which children acquire this linguistic ability corresponds to the age at which they succeed in reorienting in the small room (around 6 years of age). However, from an adaptive-combination view, older children and adults may be more proficient feature users because they have had more experience using features alone and in conjunction with geometric cues. Such experience would allow them to better understand the circumstances under which features should or should not be relied upon.
Additionally, feature use among nonhuman animals provides evidence against a language-based approach to overcoming modularity. Several nonhuman animals use features during reorientation when tested with a conflict paradigm (chicks: Chiandetti, Regolin, Sovrano, & Vallortigara, 2007; Sovrano & Vallortigara, 2006; Vallortigara, Feruglio, & Sovrano, 2005; pigeons: Kelly et al., 1998; fish: Sovrano, Bisazza, & Vallortigara, 2007). In this procedure, the animal learns a stable spatial layout with both geometric and feature cues, is removed from the environment while the feature is displaced to another location in the room, and then is placed back in the room for the reorientation test. The result is a forced-choice reference memory paradigm in which the animal must decide whether to reorient on the basis of the current feature location or the geometric information. For example, fish were trained to reorient within a rectangular fish tank that had a distinct landmark of one blue wall (Sovrano et al., 2007). The landmark was then moved to an adjacent wall prior to testing, thereby changing the length of the colored wall (either from long to short or from short to long). The displacement of the feature between training and testing put the location of the feature cue in direct conflict with the learned geometric information given by the shape of the room. Findings from this and other conflict studies suggest that geometric information is of greater importance during reorientation in smaller spaces, whereas feature cues are relied upon more in larger spaces.
Evidence from children and nonhuman animals suggests that room size and experience influence feature use during reorientation. However, associative learning, rather than actual reorientation using features, may underlie feature use in reorientation paradigms (e.g., a target object may be linked with a specific hiding box regardless of its placement in a layout; Lee, Shusterman, & Spelke, 2006). In the experiments reported here, we investigated whether there are fundamental differences among the reorientation processes of human adults, children, and nonhuman animals by testing adults using a conflict reorientation procedure. Our findings also bear on the issue of whether associative learning drives feature use in a mature spatial system that is forced to choose between spatial cues. Specifically, we examined the effect of placing geometric and feature cues in conflict when adults reoriented within different-sized environments (Experiments 1 and 2) and when prior experience encouraged reliance on different cues (Experiment 2). In accordance with the adaptive-combination view, we predicted (a) that corners identified by feature cues would be more likely to be chosen in a larger room, where distal landmarks were likely more valid and salient, than in a smaller room (Experiment 1), and (b) that participants would rely on feature cues in the smaller room when prior training in the larger room had demonstrated that features can be a useful guide (Experiment 2). If the conflict procedure yielded analogous results among humans and nonhuman animals (e.g., an effect of room size and prior experience, regardless of feature placement), such parallels would suggest that, across ages and species, feature and geometry use during spatial reorientation depends on a weighting system, rather than an encapsulated module that is overcome only through language or associative learning.
GENERAL METHOD
Temple University undergraduates were tested in either a small all-white rectangular enclosure (4 ft × 6 ft × 6 ft) or a larger all-white enclosure (8 ft × 12 ft × 8 ft). The larger room was 4 times the area of the smaller one, but the geometric information remained constant given that the ratio of long to short wall lengths was the same in the two rooms. Both enclosures were illuminated with symmetrically positioned lights. Identical plastic containers were placed at the four corners to serve as potential hiding places for the target, a pair of keys. Participants listened to white noise through headphones so that they could not use directional sound cues. The landmark feature was a removable piece of brightly colored, patterned fabric (approximately 1.5 ft × 4.5 ft), distinct from the all-white background but flush with the wall.
Four training trials, in which the experimenter hid the target in the same corner each time, established a stable spatial layout. On each trial, participants watched the target being hidden, were blindfolded, and were then disoriented by being slowly turned in circles for at least 10 full rotations with two direction changes. Participants then took off the blindfold, and the experimenter asked, “Where did I hide the keys?” Participants were instructed to point to the corner they would search first. After training, participants were instructed that the test trials would be the same and that “the keys will be in the same place as they were during training, but you will not see me hide them this time.”
Following training, participants were taken out of the room and drew a sketch of the enclosure so we could evaluate their encoding of its geometry and features. While participants waited outside the room, the experimenter went back in to move the feature to an adjacent wall. Participants were then blindfolded and led back into the middle of the room for the test trial. After the disorientation procedure, participants pointed to the corner where they thought the target object was located. No feedback was given, and the experimenter asked which corner they would pick if they needed to make a second choice. Participants were escorted out of the room and given a 1-min break before the next trial. They were then blindfolded and led back into the room for disorientation and the second test trial, with the feature location in the same shifted position as in the first. Following the test trials, participants were debriefed about the purpose of the experiment and asked about any strategies they used.
All searches were coded as at a landmark corner, a geometrically appropriate corner, or an error corner (see Fig. 1a). There was no designated “correct” location because the feature had been moved from its position during training. For each of the two test trials, we calculated scores for landmark and geometric searches: the proportion of searches at the corner where the landmark had been moved during testing and the sum of the proportion of searches at the two geometrically appropriate corners, respectively.
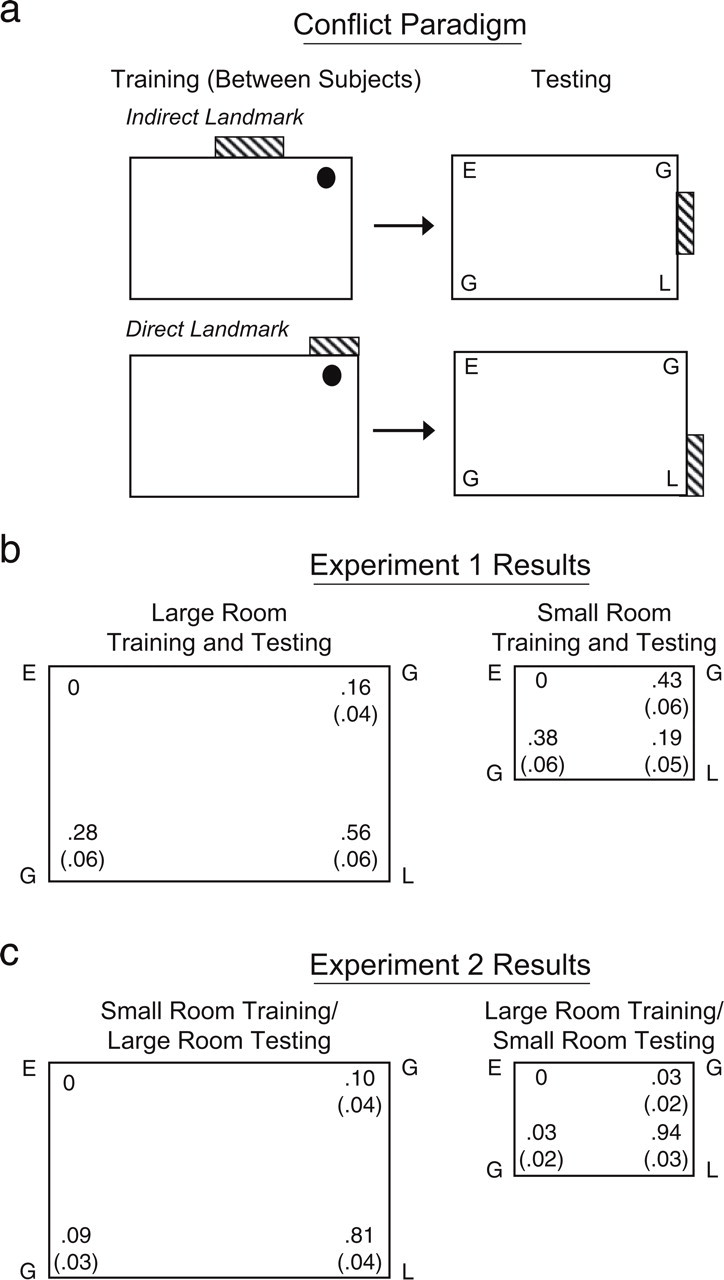
Illustration of the training and testing procedure (a) and experimental results (b and c). During training, the feature location (i.e., whether the landmark directly or indirectly marked the corner where the target was hidden) was manipulated between subjects. During testing, the landmark was moved to an adjacent wall. Participants' search choices were coded as follows: L = landmark corner; G = geometrically appropriate corner; and E = error corner. In Experiment 1 (b), half the participants were trained and tested in the smaller room, and half were trained and tested in the larger room; in Experiment 2 (c), half the participants were trained in the smaller room and tested in the larger room, and half were trained in the larger room and tested in the smaller room. The diagrams indicate the proportion of first searches (with standard errors in parentheses) at each corner. First searches were collapsed across test trials and landmark locations, as there were no main effects of these variables.
In both experiments, the feature either directly or indirectly marked the hiding corner (see Fig. 1a), in a between-subjects design. Facing position following disorientation was manipulated within subjects, and both feature location (direct vs. indirect) and facing position were counterbalanced and matched across conditions (smaller room vs. larger room). Test trial (1 vs. 2) was included as a within-subjects variable in the statistical analyses.
EXPERIMENT 1
In Experiment 1, we tested the prediction that participants reorienting in the small room would choose geometric corners, whereas those in the larger room would choose the corner associated with the feature. The 32 participants in Experiment 1 were ages 18 through 21 (M = 18.73 years); 6 males and 10 females were randomly assigned to the smaller room for training and testing, and 7 males and 9 females were trained and tested in the larger room.
Results
The diagrams drawn by the participants prior to testing suggested that they had accurately encoded the experimental setup with respect to the geometric information (63% of participants drew an obviously nonsquare rectangle, and the remaining 37% drew a rectangle that more closely resembled a square); performance was identical in the two room-size conditions. Participants were also accurate in marking the location of the feature relative to the hiding spot in both the larger room (94% correct) and the smaller room (75%); the difference between the room sizes was not significant, t(30) = 1.46, p = .15. All participants reported noticing that the landmark location had changed from training to testing.
There was no effect of gender, F(1, 24) = 0.79, p = .38; feature location (direct vs. indirect), F(1, 24) = 1.18, p = .29; or trial, F(1, 24) = 2.98, p = .10, on the proportion of landmark first searches, but there was a significant effect of room size, F(1, 24) = 8.19, p < .01.2 Adults in the larger room tended to search first at the landmark corner, whereas those in the smaller room based their initial searches on geometric information; this difference between conditions was significant, t(30) = 3.16, p < .01, d = 1.12 (see Fig. 1b). After choosing a corner, participants were immediately asked where they would search if they had a second choice. In the smaller room, participants were equally likely to base their choice on geometry (M = .53, SE = .05) and to base their choice on the feature (M = .38, SE = .05), t(15) = 0.86, p = .40, but participants in the larger room searched the geometric corners (M = .75, SE = .04) more than the landmark corner (M = .16, SE = .03), t(15) = 4.54, p < .01. Thus, search patterns differed depending on the size of the testing environment. Initially, adults in the larger room picked the landmark corner more often than did those in the smaller room. Given a second choice, adults in the smaller room picked the geometric and landmark corners equally often, whereas those in the larger room searched the two geometric corners.
Discussion
Although adults can easily use both geometric and feature cues to reorient when the two kinds of cues agree, the conflict situation forces them to choose one cue over another. As predicted, adults in Experiment 1 made more geometry-based choices in the smaller room than in the larger room, and made more feature-based choices in the larger room than in the smaller room. The critical finding here is that the choice depended on the size of the enclosure. This effect mirrors the room-size effect found for young children and nonhuman animals. If reorientation is guided by a modular mechanism focused on geometric information, featural cues should not affect reorientation strategy differently within environments of different sizes. However, if navigation occurs via a weighting mechanism, as proposed in the adaptive-combination view, the increased use of features in a larger space in a conflict situation is expected.
Lee et al. (2006) have argued that the successful use of features in previous studies demonstrates a two-step process: geometric reorientation followed by use of a direct association between the landmark and the target. The present data bear on this issue. The feature was not serving as a direct beacon in this case, as there were no significant differences overall in feature use between the indirect- and direct-landmark conditions (see also Learmonth et al., 2001). Additionally, if feature use is only secondary to use of geometry in a two-step process of reorientation, it is puzzling why participants in a forced-choice situation used features preferentially in a larger room and used geometry in a smaller room—one would think that they would use either the first or the second step in the two-step process in a fashion unaffected by room size.
EXPERIMENT 2
In the modular view, spatial language is the bridge to overcoming encapsulation. Brief experiences that demonstrate the usefulness of features as a spatial cue should have no effect on subsequent reorientation according to this view. By contrast, such experiences should modify weightings according to the adaptive-combination view. Experiment 2 tested the prediction that training in a larger room will increase feature use in a smaller room. We replicated Experiment 1 but switched room sizes between training and testing. Hence, half the adults were trained in the larger room and tested using the displaced feature in the smaller room, whereas the other half were trained in the smaller room and tested in the larger room. There were 32 participants between the ages of 18 and 24 (M = 19.14 years); 6 males and 10 females were randomly assigned to training in the smaller room and testing in the larger room, and 6 males and 10 females were trained in the larger room and tested in the smaller room.
Results
Room drawings again suggested accurate encoding of the geometry of the experimental setup (75% of participants drew an obviously nonsquare rectangle, and the remaining 25% drew a rectangle that more closely resembled a square), as well as of the location of the feature relative to the hiding spot (87.5% correct); performance was identical in the two room-size conditions. All participants reported noticing that the size of the enclosure had changed from training to testing, but only about half of the participants reported that they noticed the feature had moved (9 tested in the larger room and 8 tested in the smaller room).
There were no effects of feature location (direct vs. indirect), gender, trial, or room size on either the proportion of landmark searches or the proportion of geometric searches (all Fs < 2.05, ps > .17). Adults trained in the larger room and tested in the smaller room overwhelmingly chose the landmark corner over the geometric corners on their first search, t(15) = 13.33, p < .001, d = 3.05 (see Fig. 1c), whereas in Experiment 1, adults tested in the smaller room used geometry. The adults trained in the smaller room also based their first search on the location of the feature in the larger room in Experiment 2, t(15) = 4.04, p = .001, d = 2.20. The geometric corners were picked as a second choice, both by adults tested in the smaller room (geometric corners: M = .85, SE = .04; landmark corner: M = .03, SE = .02), t(15) = 8.06, p < .001, and by adults tested in the larger room (geometric corners: M = .81, SE = .03; landmark corner: M = .13, SE = .03), t(15) = 6.21, p < .001.
The crucial test of training effects was to determine whether feature use in the smaller testing room was higher for adults trained in the larger room (Experiment 2) than for those trained in the smaller room (Experiment 1). As predicted, adults trained in the larger room and tested in the smaller room in Experiment 2 initially searched the landmark corner significantly more often than did adults trained and tested in the smaller room in Experiment 1 (.94 vs. .19; see Fig. 1), t(30) = 8.49, p < .001, d = 3.00. However, the effect of training was not symmetric. Adults trained in the smaller room and tested in the larger room in Experiment 2 also chose the landmark corner more often than did adults trained and tested in the smaller room in Experiment 1 (.81 vs. .19; see Fig. 1), t(30) = 2.11, p = .02, d = 0.75. Therefore, following training, feature use transferred from the larger to the smaller room, but geometry use did not transfer from the smaller to the larger room.
In addition to training, the change in scale of the room itself (the increase or decrease in area size) may have increased feature use in Experiment 2. To examine the impact of scale changes, we compared feature use within the larger-room testing environment in the two experiments. If training in the larger room was the only factor that influenced feature use in Experiment 2, adults tested in the larger room would not have shown significantly more feature use in Experiment 2 than in Experiment 1. In fact, however, adults tested in the larger room in Experiment 2 used the feature more than did those trained and tested in the larger room in Experiment 1 (.81 vs. .56; see Fig. 1), t(30) = 2.11, p = .04, d = 0.74. Thus, both training and the changes in scale influenced feature use in Experiment 2. Experience in the larger room during training strengthened the salience and validity of the feature cue and increased its use during reorientation in the smaller room, whereas scale changes gave added weight to features during testing in both rooms.
Discussion
This is the first study of human reorientation to demonstrate feature-dominated search behavior when geometric information provided opposing information. The feature guided reorientation during testing in both the small and the larger rooms in this conflict situation. Adults initially searched for the keys according to the location of the landmark, picking a geometric corner only as a second choice, regardless of the size of the enclosure during testing. Further, the adults were not simply relying on the most salient cue, which would have been geometry in the smaller room, but rather were mapping the locations from encoding on the basis of changes in the salience of cues in the testing space. This finding suggests use of a weighted combination of features and geometry, rather than just use of the most salient cue, and also highlights the flexible nature of the adaptive-combination view in its ability to incorporate changes in cue salience from encoding to action.
The room-size effect found in Experiment 1 dissolved once participants were exposed to the larger room, either during training or during testing. It appears that prior experience using features is pivotal in encouraging their use in small environments. If one is given the opportunity, even briefly, to realize that features are useful cues in determining location and orientation in a very small space, where geometric cues are often dominant, this experience will generalize to other similar environments. These results, in addition to those from human children (Learmonth et al., in press; Twyman et al., 2007), suggest that learning history is a strong determinant of the weights given to spatial cues, as proposed in the adaptive-combination view.
Features are weighted more heavily when prior experience has shown them to be useful. However, training did not have the same effect on use of geometric cues to reorient. Exposure to the larger space during testing was apparently sufficient to overcome the prior exposure to a more geometrically dominant environment (the small room). The change of scale of the room itself (either an increase or a decrease in area) between training and testing also influenced adults' use of features over geometry. However, this finding is also evidence against use of a geometric module to guide reorientation in the sense that the change in scale of the rooms led to greater reliance on the feature, even though the relative geometric information (ratio of long to short wall length) remained constant. Use of features appears to be less influenced by change of scale and thus outweighed geometry as the more stable and reliable cue in the two environments at the time of testing. Geometric cues may be more fragile than features when the environment changes in scale and may dominate reorientation only when one is in exactly the same small environment.
GENERAL DISCUSSION
The geometric and featural properties of space contribute to adult reorientation in different ways depending on factors such as the size of the environment and the individual's prior experience. In the present experiments, adults used both geometry and features in order to reorient in fully enclosed rectangular spaces. We applied a conflict paradigm previously used with nonhuman animals to examine the specific role of each kind of cue within different environments. When features were pitted against geometry in Experiment 1, adults chose to reorient by the geometric room shape in a very small enclosure, but used features to guide navigation in a larger space. However, the effects of exposure to geometric and feature cues are not symmetric. In Experiment 2, adults were trained in either the larger or the smaller room and then performed the conflict test in the opposite-sized room. They reoriented strictly by the location of the feature, regardless of room size. For the first time in a reorientation study, humans overwhelmingly chose feature cues that contradicted the information given by geometric cues—an instance of features dominating geometry. Thus, it appears that experience in the larger room increased feature use in a very small space where geometry was typically dominant, whereas training in the small room did not produce a more geometrically driven search pattern in the larger room. These results, along with those from nonhuman animals and children, support the case for an adaptive-combination view emphasizing the importance of the cue itself (validity and salience) and the learning history of the organism.
Taken together, these results coincide with predictions of the adaptive-combination view and suggest that the development of spatial navigation is best accounted for by a flexible approach, in that geometric information does not automatically dominate features during reorientation. In fact, reorientation guided by featural cues is highly malleable given sufficient prior experience using those cues and the opportunity to encode them with low variability and high certainty, as in the case of a distal landmark in a large space. Feature use not only is observed as a result of associative learning (Lee et al., 2006), but also can guide reorientation, as demonstrated by these experiments. The adaptive-combination approach explains why some sources of information are weighted more heavily than others and provides a better account for the data than the geometric-module hypothesis.
Reorientation is a prime example of how humans use distinct types of spatial information to different degrees in order to navigate to remembered locations. To succeed in such navigation, humans must align the location and orientation information from their current surroundings within a previously formed mental representation of the space, or cognitive map (Tolman, 1948). The nature of such representations has long been debated. Some researchers have proposed that representations are egocentric (based on locating objects relative to parts of the body) and that an encapsulated geometric module is used to solve the problem of disorientation (Wang & Spelke, 2002). However, others have argued that both egocentric and allocentric representations perform in parallel, with their use depending on such factors as the degree of self-motion, the size of the environment, and prior experience within the environment (Burgess, 2006). Clearly, the present findings support the latter view.
Footnotes
1It is only when adults perform a concurrent task during reorientation that their search patterns begin to show the limited feature use found among rats and young children, although the degree to which this reduction in feature use is due to interference is debatable in that replication studies in which adults have performed a concurrent verbal task have not produced the chance performance originally found by Hermer-Vazquez et al. (1999) and results vary with the type of secondary task used (Hermer-Vazquez et al., 1999; Hupbach, Hardt, Nadel, & Bohbot, 2007; Ratliff & Newcombe, 2008).
2Participants never chose the error corner on first searches (in either experiment), so the proportions of landmark and geometric searches were perfectly complementary, p(landmark search) = 1 − p(geometric search). Therefore, only landmark searches are reported.
Acknowledgements
Portions of this research were presented at the 48th annual meeting of the Psychonomic Society, November 2007, and at the 29th annual meeting of the Cognitive Science Society, August 2007. Funding was provided by National Science Foundation Grant BCS0414302.