
Abstract
Pavlovian fear conditioning is a widely used model of the acquisition and extinction of fear. Neural findings suggest that the amygdala is the core structure for fear acquisition, whereas prefrontal cortical areas are given pivotal roles in fear extinction. Forty-eight volunteers participated in a fear-conditioning experiment, which used fear potentiation of the startle reflex as the primary measure to investigate the effect of two genetic polymorphisms (5-HTTLPR and COMTval158met) on conditioning and extinction of fear. The 5-HTTLPR polymorphism, located in the serotonin transporter gene, is associated with amygdala reactivity and neuroticism, whereas the COMTval158met polymorphism, which is located in the gene coding for catechol-O-methyltransferase (COMT), a dopamine-degrading enzyme, affects prefrontal executive functions. Our results show that only carriers of the 5-HTTLPR s allele exhibited conditioned startle potentiation, whereas carriers of the COMT met/met genotype failed to extinguish conditioned fear. These results may have interesting implications for understanding gene-environment interactions in the development and treatment of anxiety disorders.
Pavlovian conditioning is the most basic of all paradigms for the study of associative learning. It provided a substantial portion of the empirical basis for the classical theories of learning, which formed the core of behaviorist psychology. The famous early work on fear conditioning in “Little Albert” (Watson & Raynor, 1920) pioneered the analysis of fear and anxiety in terms of learning, an enterprise that is still vigorously pursued (e.g., Lissek et al., 2005; Mineka & Zinbarg, 2006; Öhman & Mineka, 2001). Despite the iconic status of Pavlovian conditioning as an epitome of learning and environmentalism, we show in this article that Pavlovian fear conditioning and extinction, as measured by fear-potentiated startle reflexes, are modulated by common genetic polymorphisms in the serotonin and dopamine systems, respectively.
Pavlovian fear conditioning imbues a relatively neutral stimulus (the conditioned stimulus, or CS) with fear-evoking properties as the result of its co-occurrences with an aversive event (the unconditioned stimulus, or US) that threatens the well-being of the organism. As a result of fear conditioning, therefore, remnants of traumatic events will stick to trauma-exposed individuals as components of fear memories evoked by stimuli associated (more or less consciously) with traumas. Accordingly, fear conditioning has carried a heavy explanatory burden in psychological theories of anxiety disorders such as phobias (e.g., Öhman, Dimberg, & Öst, 1985; Seligman, 1971), panic disorder (e.g., Bouton, Mineka, & Barlow, 2001), and posttraumatic stress disorder (e.g., Orr et al., 2000). Extinction (i.e., the waning of fear as a result of the CS being presented in the absence of the US) has obvious implications for the treatment of phobias and other anxiety disorders (Anderson & Insel, 2006). Indeed, it inspired the exposure therapies that have provided highly effective treatments of anxiety disorders (Barlow, 2002). In direct support of the common assumption that fear conditioning is a potential mechanism of anxiety disorder, meta-analyses indicate that patients diagnosed with anxiety disorder show more rapid fear conditioning and slower extinction than control subjects (Lissek et al., 2005).
Important progress has been made in understanding the neural mechanisms of Pavlovian fear conditioning (see reviews by Davis & Whalen, 2001; Fanselow & Poulos, 2005; LeDoux, 2000). Briefly summarized, research has shown that cortical and subcortical sensory pathways converge on the lateral nucleus of the amygdala, which houses the molecular machinery for forming associations between the CS and the US (Fanselow & Poulos, 2005; LeDoux, 2000). As a result of this associative process, the CS may activate the central nucleus of the amygdala, which recruits various fear responses through pathways to the striatum, diencephalon, midbrain, and brainstem (Davis & Whalen, 2001). The neural basis of extinction is less well understood, but both animal (e.g., Quirk & Gehlert, 2003) and human (e.g., Phelps, Delgado, Nearing, & LeDoux, 2004) data suggest that extinction depends on structures in the medial prefrontal cortex.
Behavior genetic studies suggest that about one third of the variance in human fear conditioning (Hettema, Annas, Neale, Kendler, & Fredrikson, 2003) and the risk to develop anxiety disorders (Gordon & Hen, 2004) can be attributed to genetic factors. Given that a polymorphism in the serotonin transporter (5-HTT) gene (SLC6A4) has been convincingly related to amygdala reactivity (for a meta-analysis, see Munafo, Brown, & Hariri, 2008), this polymorphism is a likely candidate to be involved in conditioned fear. A 43-bp insertion/deletion in the 5-HTT promoter, referred to as 5-HTTLPR, yields a long (l) and short (s) allele, the latter of which reduces 5-HTT expression and serotonin uptake by close to 50% (Lesch et al., 1996). In addition to having a relationship to the amygdala, this “low activity” s allele is associated with higher neuroticism scores (for a meta-analysis, see, e.g., Sen, Burmeister, & Ghosh, 2004).
The potentially pivotal role of the prefrontal cortex in extinction points to a candidate gene for extinction processes, the gene coding for catechol-O-methyltransferase (COMT), which degrades released extracellular dopamine. The COMT gene harbors an interesting single nucleotide polymorphism: COMTval158met. Carriers of the met allele have a 4-fold reduction in enzyme activity compared with carriers of the val allele, and as a result have higher extracellular dopamine levels, particularly in prefrontal areas (for a review, see, e.g., Bilder, Volavka, Lachman, & Grace, 2004). The met allele enhances prefrontal cognition and working memory (for a review, see, e.g., Bilder et al., 2004) and has been associated with negative mood states (e.g., anxiety, dysphoria) and impaired pain regulation. Furthermore, the met allele is related to higher responsiveness in, and connectivity between, brain areas involved in the evocation and regulation of negative affective responses—in particular, the amygdala, hippocampus, and prefrontal areas (for a review, see, e.g., Heinz & Smolka, 2006).
The purpose of this study was to use psychophysiological indices of fear to examine whether these polymorphisms are related to fear conditioning and extinction in humans. Given some degree of heritability of both fear conditionability and anxiety disorders, such relationships could provide mechanisms behind the gene-environment interaction in anxiety disorder. First, because of the associations between the 5-HTTLPR s allele and amygdala reactivity, and between the amygdala and fear conditioning, we hypothesized that carriers of this allele would exhibit enhanced fear acquisition. Second, we hypothesized a selective effect of the COMT met allele on extinction because of the prefrontal focus of the actions of COMT and the suggested role of this brain area in extinction and other forms of emotion regulation.
Startle blink potentiation and skin conductance responses (SCRs) were used as measures of conditioning. Fear potentiation of the startle reflex is one of the most useful indices of defensive response mobilization (Lang, Davis, & Öhman, 2000). Neurally, it reflects the influence of direct and indirect connections from the amygdala to the primary startle-reflex pathway in the brainstem (Davis & Whalen, 2001). In contrast to the SCR, which can be dissociated from amygdala activations (Tabbert, Stark, Kirsch, & Vaitl, 2006) and appears to primarily reflect cognitive contingency learning (Lovibond & Shanks, 2002), startle potentiation appears to index a basic, affective level of fear conditioning largely independent of higher cognition (Hamm & Weike, 2005; Öhman & Mineka, 2001).
METHOD
Participants
Eighty-one students of the University of Greifswald were recruited by advertisements, filled in an informed-consent form (approved by the ethics committees at the Karolinska Institutet and the University of Greifswald), and donated 20 ml of blood for DNA extraction and genotyping. Thirty-four of these students completed the experiment before being genotyped, and 47 students were genotyped before being invited to participate in the experiment. Out of the latter 47 students, 29 were selectively invited to participate in order to balance out the allele frequencies and sex distribution in the experimental sample. Thus, 63 volunteers participated in the experiment (both participants and the experimenter were blinded to genotype) and were paid €15.
We excluded 6 (3 male, 3 female) participants from the data analysis because of technical problems and 9 (3 male, 6 female) participants because they could not correctly report the CS-US contingency at the end of the conditioning phase (see Procedure). Lack of awareness, which was unrelated to our specific genotype groups, typically compromises SCRs, but not startle conditioning (Hamm & Weike, 2005; Lovibond & Shanks, 2002), and thus would likely introduce an irrelevant source of variance. The final sample comprised 48 participants.
Genotyping
DNA extraction from whole blood was performed using standard methods (Autopure LS system, Gentra Systems, Minneapolis, MN; see Lindblom & Holmlund, 1988). For 5-HTTLPR genotyping, primers (Thermo Scientific, Ulm, Germany) flanking the 5-HTTLPR of the 5-HTT gene (forward 5′-TGAATGCCAGCACCTAACCCCTAA-3′ and reverse 5′-GAATACTGGTAGGGTGCAAGGAGA-3′) were used. Polymerase chain reactions (PCRs) were carried out using the following cycling conditions: an initial 5-min step at 95 °C; followed by 10 cycles consisting of denaturation at 95 °C for 1 min, annealing at 65 °C (decrease of 0.5 °C with each cycle) for 1 min, and elongation at 72 °C for 1 min; followed by 39 cycles consisting of denaturation at 95 °C for 45 s, annealing at 60 °C for 1 min, and elongation at 72 °C for 1 min; followed by a final step at 72 °C for 4 min. Reactions were performed in 10 × Reaction buffer (Finnzymes, Espoo, Finland), 1.5-mM MgCl2, 20 ng of genomic DNA, 0.75 μl of dNTPs (Larova, Teltow, Germany; 10 mM), 0.625 μl of each primer (10 mM), 0.0375 μl of 7-deaza-dGTP (Roche, Mannheim, Germany), and 0.5 μl of TaqPolymerase (Dynazyme, Finnzymes, Espoo, Finland). The PCR products (10 μl) were separated into short (336 bp) and long (379 bp) fragments by electrophoresis on 2.5% agarose gels with 2% normal agarose (Certified™ Molecular Biology Agarose, Bio Rad, Hercules, CA) and 0.5% low-melting agarose (Sea Plaque® GTG Agarose, Cambrex Bio Science, Rockland, ME) with ethidium bromide in TRIS-Borat-EDTA-Buffer (TBE). After 2.5 hr of electrophoresis in TBE, the products were visualized by ultraviolet illumination.
COMTval158met (rs4680) genotype was determined on an ABI HT7900 (Applied Biosystems, Foster City, CA) using the TaqMan® allelic discrimination (5′ nuclease assay; Livak, 1999).
Stimulus Materials
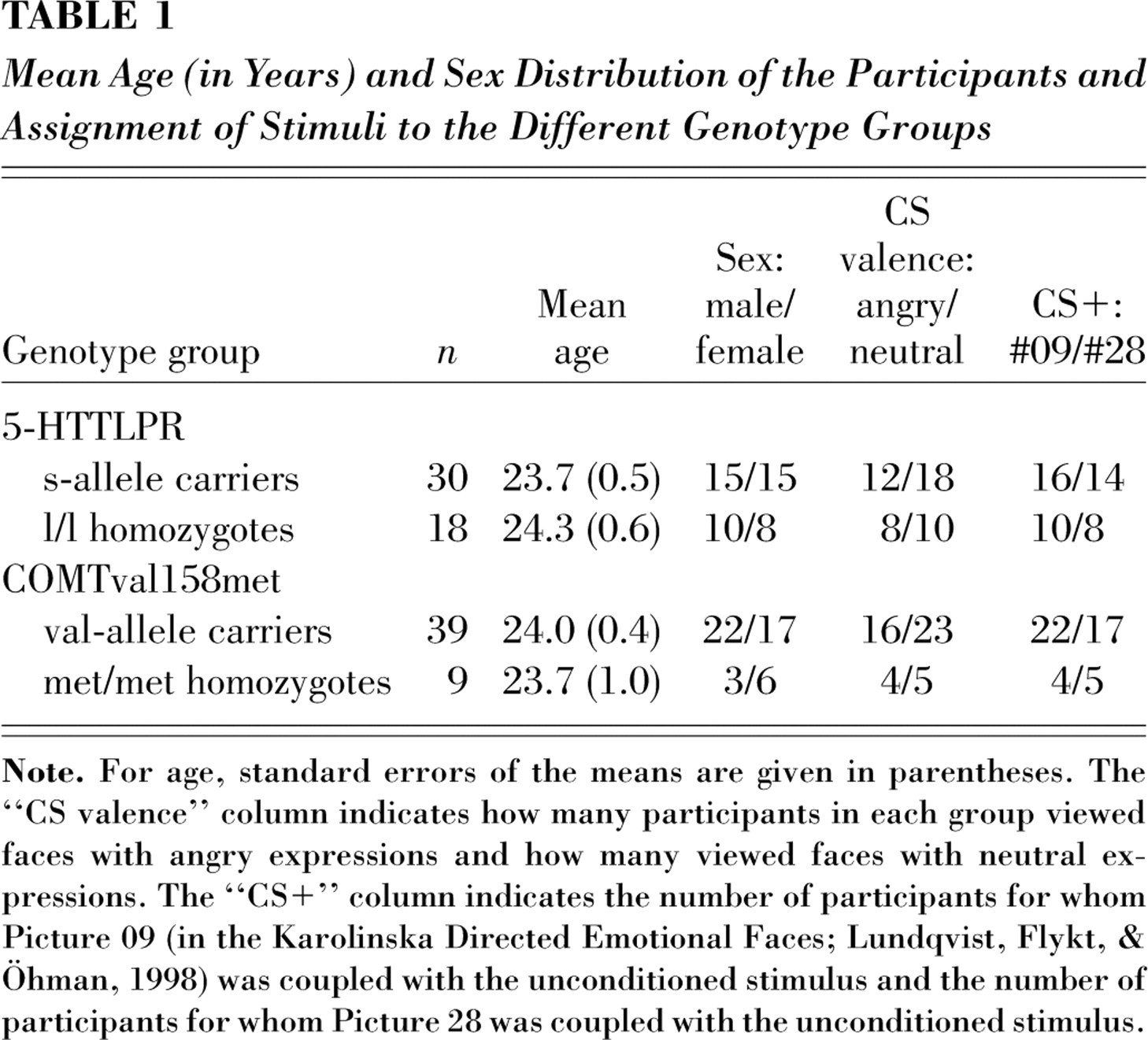
Four different color pictures depicting male faces were selected from the Karolinska Directed Emotional Faces (Lundqvist, Flykt, & Öhman, 1998) to serve as CSs (two pictures depicted neutral expressions, and two depicted angry facial expressions1). Which picture was coupled to the US, the valence of the expressions presented (each participant saw only neutral faces or only angry faces), and the stimulus sequence used (eight different sequences) were balanced across the genotype groups; Table 1 shows the distribution of the variables of interest in the final sample of 48 participants. The pictures (visible size: 126 cm × 93 cm) were projected onto a screen approximately 2 m in front of the participant using a projector (Sanyo PLC-XU86) that was situated in an adjacent room.
Mean Age (in Years) and Sex Distribution of the Participants and Assignment of Stimuli to the Different Genotype Groups
The US was a 500-Hz monopolar DC-pulse electric stimulation applied above the participant's right ankle in a 10-ms train of 1-ms single pulses. It was generated by a commercial stimulator (Grass Instruments S48K, West Warwick, RI), isolated (SIU5), and transmitted via a constant-current unit (CCU1) to a bipolar electrode (F-E10S2). The mean physical intensity chosen during the intensity adjustment for the experiment (see Procedure), the sensitization effect (increase in startle and SCRs after US intensity adjustment), and unconditioned SCRs to the US did not differ between the 5-HTTLPR groups or between the COMTval158met genotype groups.
Startle probes were 50-ms bursts of 95-dB[A] white noise (rise time < 1 ms). They were presented binaurally over Sony (MDR-CD 170) headphones.
Physiological Recordings
Startle responses were measured by recording electromyographic (EMG) activity over the orbicularis oculi muscle beneath the left eye using miniature Ag/AgCl surface electrodes. The raw EMG signal was amplified and filtered through a 30-Hz high-pass filter (Coulbourn S75-01) and a 400-Hz low-pass filter (Kemo KEM-VBF8-03; Beckenham, Kent, United Kingdom), rectified, and integrated with a time constant of 10 ms. Skin conductance was recorded using Hellinge Ag/AgCl standard electrodes placed adjacently on the hypothenar eminence of the right hand (see Weike, Schupp, & Hamm, 2007, for a more detailed description of the recording procedures).
Procedure
Experimental Groups
Carriers of the 5-HTTLPR s allele (s/s or s/l genotype) were combined to form the s-allele-carrier group and were compared with l/l homozygotes. Similarly, carriers of the COMT val allele were combined to form the val-allele-carrier group and compared with met/met homozygotes2 (see Table 1 and Table 2 for further description of these groups).
Distribution of Genotypes in the Sample

Day 1 (Fear Acquisition)
The experimental procedure on Day 1 included a baseline phase (5 min of familiarization with the lab situation and presentation of four startle probes for habituation), individual adjustment of the US to a level described as “highly annoying but not painful,” and conditioning. During conditioning (see Fig. 1a), participants viewed each of two CS pictures nine times, in a mixed order. Each picture was presented for 6 s. One of the pictures (CS+) was always paired with the 10-ms US, which occurred simultaneously with the offset of the picture (100% reinforcement, delay conditioning); the other picture (CS−) was never coupled to the US. Acoustic startle probes were presented 4 or 5 s after picture onset for six of the nine presentations of each CS and during six intertrial intervals (ITIs; ITI = 10–18 s). Participants were instructed to attend to the pictures, but no information about the CS-US contingencies was given. The conditioning phase ended with a standardized postexperimental awareness interview (cf. Bechara et al., 1995), so we could assess awareness of the CS-US contingency. Participants also retrospectively rated the US, acoustic startle probe, CS+, and CS− for valence and arousal using the Self-Assessment Manikin (Lang, 1980); these ratings did not differ between the 5-HTTLPR groups or between the COMTval158met genotype groups.
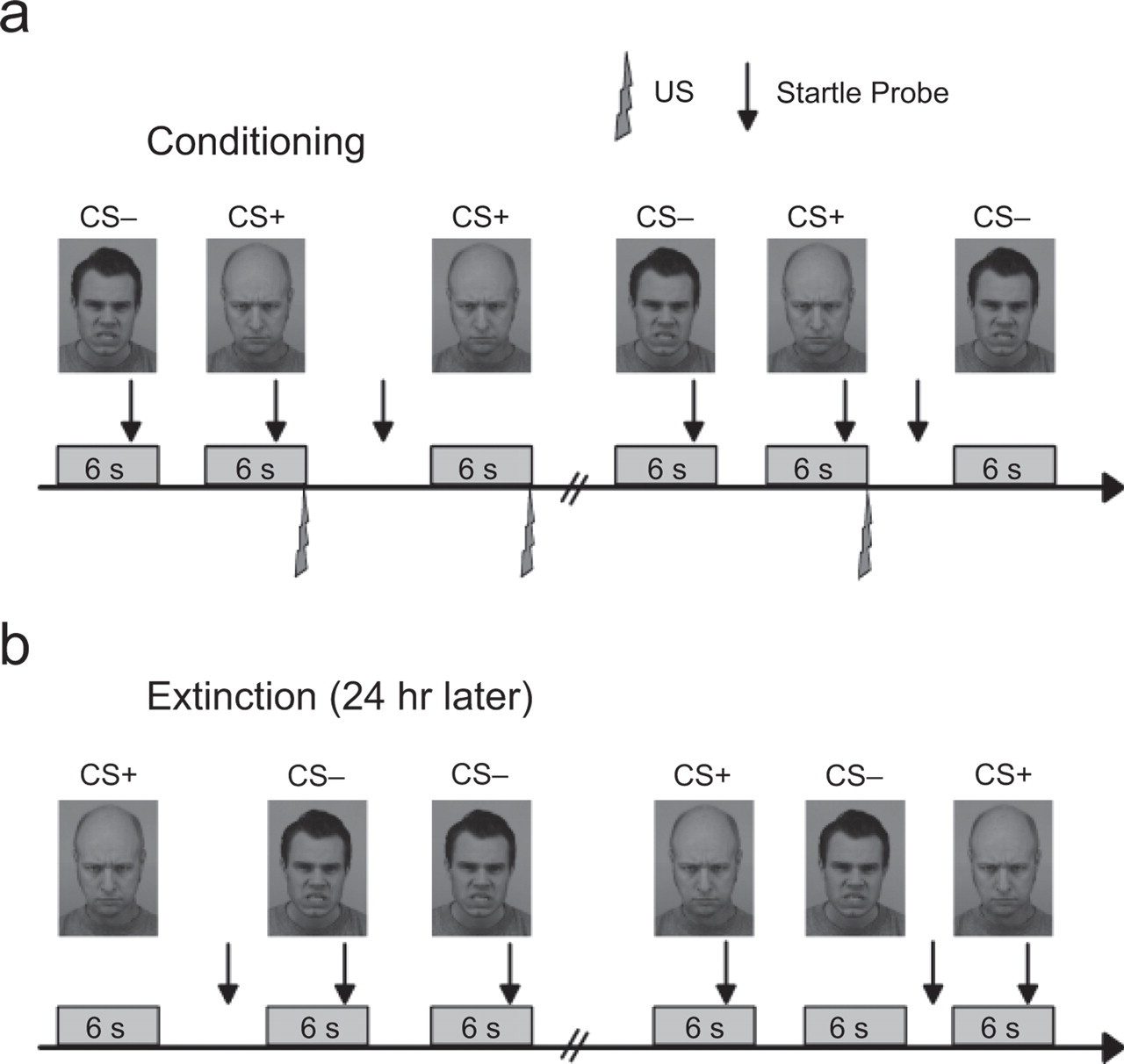
The fear-conditioning paradigm. During conditioning (a), participants viewed each of two conditioned stimuli (CSs) nine times, in a mixed order. One of the pictures (CS+) was always paired with the 10-ms unconditioned stimulus (US); the other picture (CS−) was never coupled to the US. Acoustic startle probes were presented 4 or 5 s after picture onset for six of the nine presentations of each CS and during six intertrial intervals (ITIs). During extinction (b), the CS+ and CS− were presented 18 times each without administration of any further USs. Startle probes were presented 4 or 5 s after picture onset for 12 of the 18 presentations of each CS and during 12 ITIs.
Day 2 (Extinction)
The experimental procedure on Day 2 (approximately 24 hr later; see Fig. 1b) included a baseline phase (presentation of four startle probes for habituation) and extinction. During extinction, the CS+ and CS− were presented 18 times each without administration of any further USs. Startle probes were presented 4 or 5 s after picture onset for 12 of the 18 presentations of each CS and during 12 ITIs. Again, participants were instructed to attend to the pictures, but no information about the CS-US contingencies was given. After completing the experiment, participants were debriefed and paid.
Data Reduction and Response Definition
The magnitude of the startle eyeblink (in microvolts) was measured from onset to peak, as described previously by Weike et al. (2007). Blink magnitudes were normalized using z-standardization and converted to T scores to ensure that all participants contributed equally to the group means. The T-score calculation, 50 + (z × 10), results in a distribution with an overall mean of 50 and a standard deviation of 10 for each participant.
SCR magnitude (in microsiemens) was scored as the largest response occurring 0.9 to 4.0 s after picture onset. Logarithms were computed for all values, to normalize the distribution (Venables & Christie, 1980), and these log values were range-corrected (individual score/individual maximum response) to account for interindividual variability (Lykken & Venables, 1971).
Startle and SCR measurements that showed recording artifacts or excessive baseline activity were discarded. Thirty-four of the 3,120 startle measurements were discarded (1.1%; 0–6 per participant), as were 70 of the 3,360 SCR measurements (2.1%; 0–13 per person).
Data Analysis
Data were analyzed separately for the conditioning and extinction phases using SPSS 15 for Windows. Fear-potentiated startle was measured by subtracting the mean magnitude of startle responses elicited by probes during ITIs from the mean magnitude of startle responses elicited by the same probes presented during the CSs. We calculated the mean startle potentiation to the CS+ and the CS− using individually standardized T-score differences. Furthermore, we calculated scores for mean CS discrimination by subtracting the mean T-score startle magnitude elicited during the CS− from the mean T-score startle magnitude elicited during the CS+. To make sure that T scores reflected the group differences in stimulus effects, rather than baseline differences, we examined group differences in raw-score startle responses to ITI probes separately. SCR conditioning was assessed as mean CS discrimination.
We performed repeated measures analyses of variance (ANOVAs) with stimulus (CS+ vs. CS− vs. ITI for startle; CS+ vs. CS− for SCR) as a within-subjects variable and 5-HTTLPR genotype (s carriers vs. l/l homozygotes) or COMTval158met genotype (val carriers vs. met/met homozygotes) as a between-subjects variable (N = 48). If an ANOVA revealed a significant main effect for genotype or a significant Stimulus × Genotype interaction, simple contrasts (CS+ potentiation, CS− potentiation, and CS discrimination) were calculated to specify this effect. For an exploratory within-subjects analysis in s-allele carriers, the same analyses were applied with COMT genotype as a between-subjects factor (N = 30). We adopted a significance level of .05, and Greenhouse-Geisser adjustments of degrees of freedom were used when appropriate. We report η P 2 as the estimate of effect size.
RESULTS
A 3 × 2 ANOVA revealed a significant interactive effect of stimulus and 5-HTTLPR genotype on startle blink responses during conditioning, F(2, 92) = 3.78, p = .026, η P 2 = .076, in the absence of significant group differences in blink response during the ITI (raw-score magnitudes in microvolts), F(1, 46) =1.07, p = .307. Contrasts showed that 5-HTTLPR s-allele carriers exhibited significantly stronger startle potentiation to the CS+ than did l/l homozygotes, p = .01, η P 2 = .132 (Fig. 2a); these two genotype groups did not differ significantly in CS discrimination or CS− potentiation. Within-group analyses revealed robust CS+ and CS− potentiation and significant CS discrimination in s-allele carriers, whereas the l/l group did not show significant CS+ or CS− potentiation or significant CS discrimination. COMTval158met genotype did not affect fear-potentiated startle during conditioning (Fig. 2c).
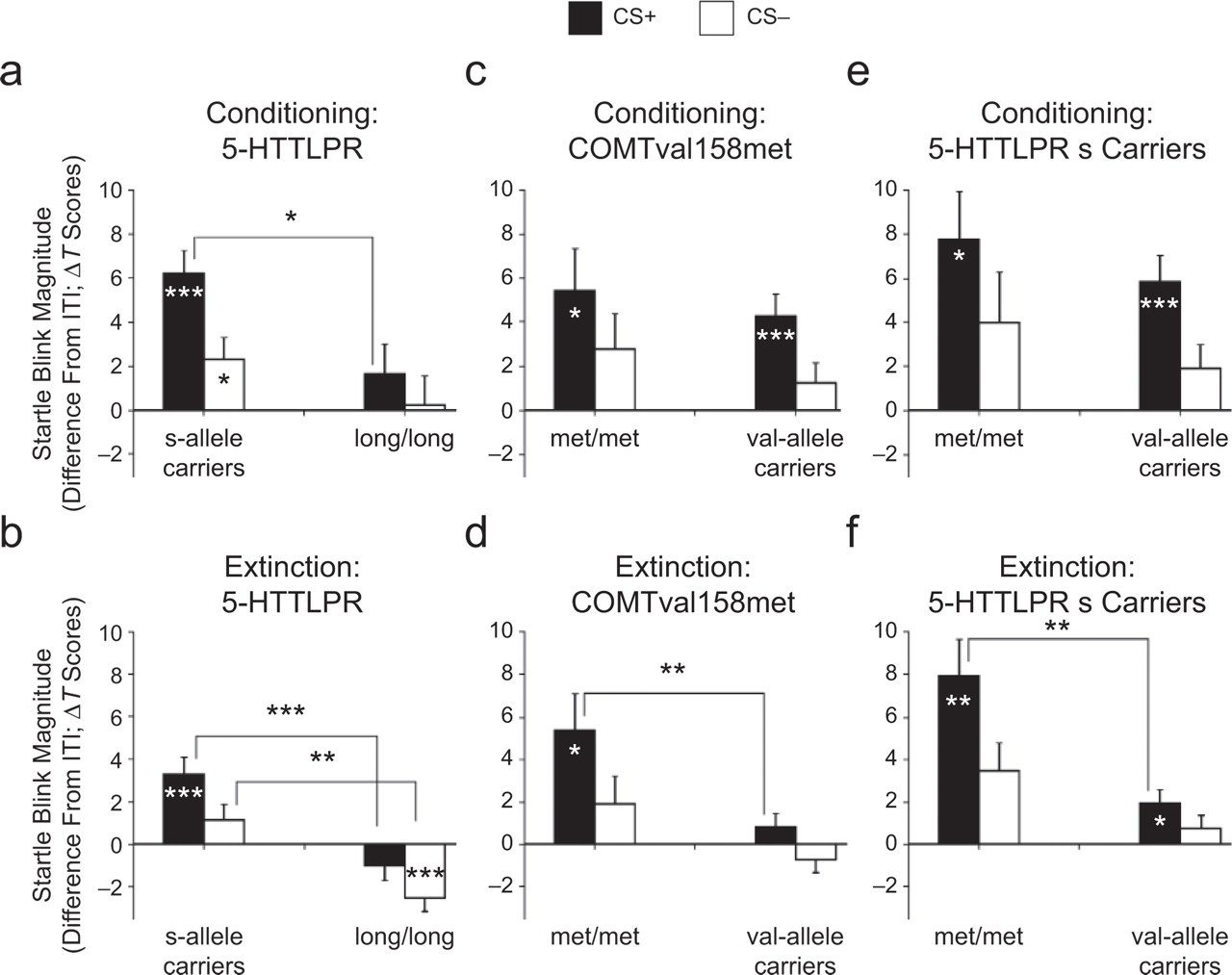
Potentiation of startle-response magnitudes as a function of genotype and stimulus. Black bars show the difference between magnitude of the startle response elicited during the CS+ (the conditioned stimulus coupled to the unconditioned stimulus) and magnitude of the startle response elicited during the intertrial interval (ITI); white bars show the difference between magnitude of the startle response elicited during the CS− (the conditioned stimulus never coupled to the unconditioned stimulus) and magnitude of the response elicited during the ITI. Results are shown for 5-HTTLPR genotype groups (a) during conditioning and (b) during extinction, for COMTval158met genotype groups (c) during conditioning and (d) during extinction, and for COMTval158met genotype groups within 5-HTTLPR s-allele carriers (e) during conditioning and (f) during extinction. Error bars represent standard errors. Asterisks indicate significant differences, ∗ p < .05, ∗∗ p < .01, ∗∗∗ p < .001.
During extinction, a significant Stimulus × 5-HTTLPR Genotype interaction was again found, F(2, 92) = 8.80, p < .001, η P 2 = .161, in the absence of group differences in startle response magnitudes during the ITI (uncorrected magnitudes in microvolts). As during conditioning, 5-HTTLPR s-allele carriers showed significantly more CS+ potentiation than did l/l homozygotes (p < .001, η P 2 = .221). In addition, the l/l group showed stronger CS− inhibition than did the s-allele carriers (Fig. 2b). CS discrimination was comparable in the two groups and significant for both.
In contrast to the conditioning phase, the extinction phase showed a significant Stimulus × COMT Genotype interaction, F(2, 92) = 5.09, p = .008, η P 2 = .100, in the absence of significant group differences in blink response during the ITI (uncorrected blink magnitudes in microvolts). The met/met homozygous group showed clearly more pronounced CS+ potentiation than did the val-allele carriers (p = .005, η P 2 = .159; Fig. 2d); CS discrimination and CS− potentiation were comparable in these two groups. Furthermore, although CS+ potentiation remained at the same level as during conditioning for the met/met group, it decreased to a nonsignificant level for val-allele carriers.
SCRs showed reliable CS discrimination in the absence of genotype effects during both conditioning and extinction. This result suggests that the participants, irrespective of genotype, successfully learned the CS-US contingencies on a cognitive level.
As the l/l group did not show reliable startle potentiation during conditioning, only s-allele carriers were selected for an exploratory within-subjects analysis assessing the effect of COMTval158met genotype on extinction (see Table 2 for the distribution of COMTval158met genotype groups within 5-HTTLPR s-allele carriers). As in the overall analysis, COMTval158met genotype did not modulate startle responses during conditioning (Fig. 2e). However, during extinction, a significant Stimulus × COMT Genotype interaction was found, F(2, 56) = 6.00, p = .004, η P 2 = .177, in the absence of significant differences in startle responses during the ITI (uncorrected magnitudes in microvolts). Those s-allele carriers who were homozygous for the COMT met allele showed significantly more pronounced CS+ potentiation than did those s-allele carriers who also carried a val allele (p = .002, η P 2 = .302), even though significant CS+ potentiation was observed for both groups (Fig. 2f). No significant differences in CS discrimination or CS− potentiation were found.
Analyses of SCRs within s-allele carriers revealed significant CS discrimination during conditioning, and a trend for CS discrimination during extinction, in the absence of an effect of COMTval158met genotype.
DISCUSSION
In summary, our results suggest the rather strong conclusion that the 5-HTTLPR and COMTval158met polymorphisms gate fear learning and extinction, respectively, as measured by fear-potentiated startle. On the one hand, only carriers of the 5-HTTLPR s allele acquired potentiated startle reactions to environmental stimuli associated with an aversive event through Pavlovian conditioning. On the other hand, homozygosity for the COMT met allele selectively blocked the ability to extinguish conditioned fear when the CS no longer was predictive of the US. Our exploratory analysis on s-allele carriers alone confirmed on a within-subjects basis that the combination of a 5-HTTLPR s allele and COMT met-homozygosity conferred an enhanced risk for acquiring fear that resisted extinction.
These findings undermine the commonly held (but mistaken) belief in an impenetrable barrier between genes and environment. According to this belief, genes are inherited, intraorganismic, causal factors, and the environment provides the arena for learned influences on behavior. Our results imply a more dynamic relationship by suggesting that genes may act through the environment by making carriers of particular gene combinations more likely than other individuals to easily pick up and retain fear of stimuli associated with threat and trauma. Thus, people carrying at least one 5-HTTLPR s allele and two COMT met alleles are likely to expand their sets of fear- and anxiety-evoking stimuli through facilitated fear conditioning and poor extinction. This process might be further accelerated by stimulus generalization and second-order conditioning, and hence such individuals may end up fearful of many stimuli that they are exposed to in their everyday environment. As a consequence, they might have frequent experiences of negative affect, which is a core characteristic of neuroticism (Clark, Watson, & Mineka, 1994). This could explain the relationship between the 5-HTTLPR s allele and neuroticism. If the fear elicited by many stimuli is intense enough to promote coping attempts in the form of avoidance, the result could be the restrictions in life options that characterize people with anxiety disorder. This could also explain why negative affect is a risk factor for anxiety disorder (Clark et al., 1994).
The two polymorphisms we studied will act synergistically in this process, one by promoting fear acquisition, and the other by slowing extinction. Nevertheless, they are independent of each other, as shown by the double dissociation, with the 5-HTTLPR polymorphism affecting acquisition but not extinction of fear, and the COMT polymorphism affecting extinction but not acquisition.
It is noteworthy that our conclusions were based on the results of conditioned startle potentiation and were not valid for SCRs. Our results show that the amount of CS+ startle potentiation differed between the 5-HTTLPR genotype groups and between the COMTval158met genotype groups during acquisition and extinction, respectively, whereas CS+/CS− differentiation was comparable between the two groups (the finding for s-allele carriers echoes results with anxiety-disorder patients; Lissek et al., 2005). Consistent with the lack of differences between groups in discriminative startle potentiation to the CS+ and the CS−, SCRs showed reliable CS+/CS− discrimination that was independent of genotype. The dissociation between our startle and SCR results in response to the CS+ probably reflects a selective impact of 5-HTTLPR and COMTval158met on a basic emotional level of fear that is manifested in startle potentiation, but not in conditioned SCRs (e.g., Hamm & Weike, 2005; Öhman & Mineka, 2001).
However, in an earlier study (Garpenstrand, Annas, Ekblom, Oreland, & Fredrikson, 2001) that measured only SCRs, s-allele carries were significantly overrepresented among individuals showing very good fear conditioning. The discrepancy between our results and those reported by Garpenstrand et al. may be attributed to different research strategies: Whereas the latter authors selected their participants on the basis of conditioning performance and compared the genotypes of people who exhibited extremely good and poor conditioning, we followed the hypothesized causal sequence and grouped participants a priori by genotype and subjected them to a fear-conditioning procedure.
The results for the COMTval158met genotype appear consistent with the tonic-phasic dopamine hypothesis for COMT (Bilder et al., 2004). Thus, the low-activity met allele should facilitate cognitive stability but jeopardize the cognitive flexibility (updating or resetting working memory content) needed to adapt to the change in conditioning contingencies during fear extinction. This emotional perseveration, observed as a failure of extinction in the met/met group, most likely reflects impaired cognitive control over emotional reactions. Our results are thus in line with the fact that the COMT met allele has been associated with both cognitive control and anxiety proneness.
Although our data suggest that the 5-HTTLPR s allele serves as a gate for fear conditioning and that carrying a COMT val allele serves as a gate for fear extinction, the generality of these conclusions across experimental conditions and measures remains to be determined. For example, one interesting possibility is that our results depended on our use of facial stimuli as CSs (Canli & Lesch, 2007). Another potentially critical factor is the intensity of the US. For ethical reasons, our US was of moderate intensity; therefore, the present findings should not be taken to imply that homozygosity for the 5-HTTLPR l allele precludes fear conditioning regardless of circumstances. Similarly, more prolonged extinction or the intense CS exposure used in single-session treatment of specific phobia (Öst, 1997) may overcome the apparent limitation to extinction we observed among met homozygotes. Examining these limitations, as well as realizing the promise of a deeper understanding of the dynamics inherent in vulnerability-stress conceptualizations of anxiety disorder, must await future research.
Footnotes
1CS valence was not included in the statistical analyses because of a nonsignificant Stimulus × Valence interaction for startle responses and SCR magnitudes (see the Data Analysis section for further information on the factors in these analyses).
2During conditioning and extinction, carriers of one 5-HTTLPR s allele (s/l genotype) and carriers of two 5-HTTLPR s alleles (s/s genotype) showed comparable CS+ potentiation, but significantly larger CS+ potentiation than the l/l homozygotes. The three COMT genotype groups did not differ from each other in CS+ potentiation during conditioning. During extinction, the met/met group showed larger CS+ potentiation than both the heterozygotes and the val/val group. However, the latter trend did not reach significance, likely because of the small sample size for this contrast (n = 9 for met/met and n = 12 for val/val). Thus, we considered it empirically justified to combine carriers of at least one 5-HTTLPR s allele into a single group and to combine carriers of at least one COMT val allele into a single group.
Acknowledgements
This work was supported by grants from the Swedish Science Research Council, the Nordic Research Council for the Humanities and Social Sciences (NOS-HS) Nordic Centre of Excellence in Cognitive Control, and the National Institute of Mental Health Center for the Study of Emotion and Attention to A.Ö., and by a grant from the Swedish Research Council to M.S. T.B.L. was supported by the German Academic Exchange Service (Deutscher Akademischer Austauschdienst). We thank Heino Mormann for technical assistance, Carmen Hamm for blood sampling, and Andreas Olsson and Armita Golkar for comments on earlier versions of this manuscript.