
Abstract
How does a physical stimulus determine a conscious percept? Binocular rivalry provides useful insights into this question because constant physical stimulation during rivalry causes different visual experiences. For example, presentation of vertical stripes to one eye and horizontal stripes to the other eye results in a percept that alternates between horizontal and vertical stripes. Presentation of a different color to each eye (color rivalry) produces alternating percepts of the two colors or, in some cases, a color mixture. The experiments reported here reveal a novel and instructive resolution of rivalry for stimuli that differ in both form and color: perceptual alternation between the rivalrous forms (e.g., horizontal or vertical stripes), with both eyes' colors seen simultaneously in separate parts of the currently perceived form. Thus, the colors presented to the two eyes (a) maintain their distinct neural representations despite resolution of form rivalry and (b) can bind separately to distinct parts of the perceived form.
Perception of a round, yellow, arcing tennis ball depends on correct integration of the object's shape, color, and motion. The percept of a unified object requires feature binding because various features of a visual stimulus are represented within distinct neural processing streams. Properties of the parvocellular (PC) neural pathway are suitable for representing form and color, whereas properties of the magnocellular (MC) pathway are suitable for representing motion. Although complete segregation of the PC and MC pathways is not maintained in areas V1 and V2 of visual cortex (Sincich & Horton, 2005), the characteristics of these pathways suggest that primary processing of color and form takes place in the PC pathway, and primary processing of motion occurs in the MC pathway (Felleman & Van Essen, 1991; Livingstone & Hubel, 1988; Zeki, 1993).
Perceptually, errors, or “illusions,” in feature binding are well known. When three different letters, each one a different color, are presented together for a fraction of a second, the percept of one letter may take on the color of another (e.g., presenting a red O and green X among the three letters yields the percept of a red X; Treisman & Schmidt, 1982). Such conjunction errors are cited as evidence for a neural binding mechanism that integrates color and form (Robertson, 2003; Treisman, 1998), even though both of these features are represented within the same (PC) neural processing stream. These fleeting conjunction errors, however, depend on the objects being presented for only a fraction of a second, usually with a separate attention-demanding task (Treisman & Schmidt, 1982). Theories for brief binding errors posit spatial attention as an integral component for combining various features at a common location in the visual field (Robertson, Treisman, Friedman-Hill, & Grabowecky, 1997; Treisman, 1998; Treisman & Gelade, 1980) or an impoverished neural representation of objects' locations (Lehky, 2000).
By contrast, the experiments reported here used stimuli presented continuously for 1 min (not brief presentation) and without an additional task to divide attention. A grating with alternating colored-and-gray stripes was presented to each eye. The color, or both the color and the orientation, of the grating were different in the two eyes so as to cause binocular rivalry. Rivalrous color stimuli normally result in a percept of either binocular color mixture or alternation in dominance between the colors (de Weert & Wade, 1988; Ikeda & Sagawa, 1979). The present study revealed a different and informative resolution of color rivalry: The two colors were seen simultaneously, but in different locations. Consider the case of an equiluminant vertical magenta-and-gray grating presented to one eye and an equiluminant horizontal green-and-gray grating presented to the other eye. Viewing these rivalrous gratings for 1 min often gave the percept of a single magenta-and-green grating, which alternated in orientation between horizontal and vertical. Both colors, each from a chromaticity presented to only one eye, appeared vivid and homogeneous. The gray stripes of the dominant vertical or horizontal form were filled with the color from the opposite eye's suppressed form, which is an error in binding color to form.
This finding shows that separate neural representations of hue for both eyes' chromatic stimuli were maintained during binocular rivalry even when the form from one eye was suppressed. Further, the neural representation for hue often was bound to a part of the visual field not stimulated by the chromatic light (a color-binding error). The color-binding errors described here show also that color is represented separately from a form's orientation, spatial phase, and spatial frequency.
GENERAL METHOD
Stimuli
Various patterns (forms) that cause binocular rivalry were presented within 2° circular apertures (e.g., Fig. 1a). The luminance of chromatic and achromatic regions was fixed at 8 cd/m2 throughout the experiments. The chromaticities used in the experiments were specified in a cone-excitation space (MacLeod & Boynton, 1979). In Experiments 1, 3, and 4, the following chromaticities were used: [L/(L + M) of 0.718, S/(L + M) of 3.3], which appeared magenta, and [L/(L + M) of 0.611, S/(L + M) of 0.3], which appeared green. Additional chromaticities were used in Experiment 2. The unit of S/(L + M) is arbitrary and set to 1.0 for an equal-energy-spectrum (EES) “white.” In each pattern, a surrounding circular “guideline” was metameric to EES; in addition, top and left nonius lines were presented to the left eye, and bottom and right nonius lines were presented to the right eye. Nonius lines also were metameric to EES. The guideline and nonius lines aided stable fixation of the rivalrous stimuli. When the two eyes were centered at the same location in the visual field, the observer perceived two vertically and two horizontally aligned nonius lines (as in the illustrations at the bottom of Fig. 1).
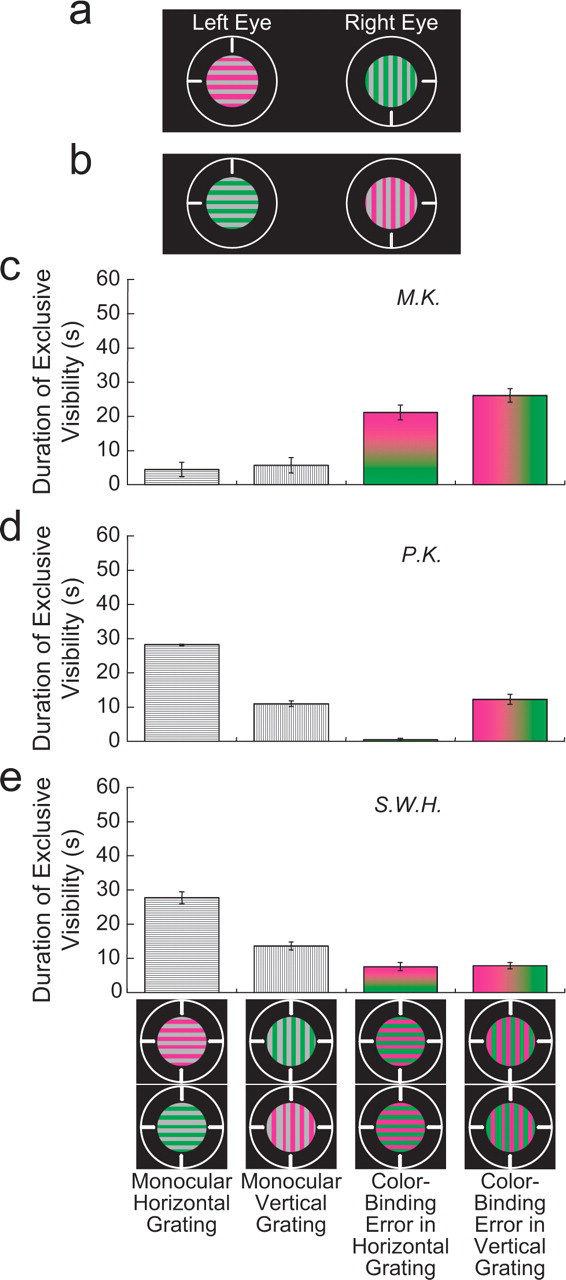
Stimuli and results for the Experiment 1 condition using perpendicularly oriented gratings. In this condition, the rivalrous stimuli consisted of equiluminant gratings with different orientations and chromaticities (a, b). The graphs (c–e) show the average duration of exclusive visibility for each of four percepts during 1 min of rivalrous presentation of the stimuli. Each graph presents the results for a different observer. The four percepts, which are illustrated at the bottom, included percepts of the monocular stimuli and percepts with color-binding errors. The heights of the bars do not sum to 60 s of presentation time because other percepts, such as a patchlike or piecemeal grating, were not included in the response options. Error bars are standard errors of the mean based on a single value from each of 3 days; each day's single value is the average duration of exclusive visibility from four counterbalanced runs (see the text).
Observers
Each observer had normal color vision as tested with a Neitz anomaloscope (Neitz Instrument Co., Tokyo, Japan). One of the observers was author S.W.H., who was knowledgeable about the experimental design and had prior experience making color matches. All other observers were naive. For all experiments, results from 3 observers are reported. An additional naive observer in Experiments 1, 3, and 4 showed color-binding errors in all conditions, though less frequently than the other observers did. Consent forms were completed in accordance with the policy of the University of Chicago's institutional review board.
EXPERIMENT 1: COLOR-BINDING ERRORS WITH RIVALROUS FORMS
Stimuli and Procedure
Experiment 1 revealed perceptual binding errors of color using rivalrous forms that differed in orientation or spatial phase. In one condition, a horizontal grating with one color was presented to one eye, and a vertical grating with another color was presented to the other eye (Figs. 1a and 1b). Both gratings were equiluminant and had a spatial frequency of 3 cycles per degree (cpd). In another condition, the two eyes' stimuli had regions that differed in spatial phase: a vertical grating (2 cpd) was presented to one eye, and a “tooth-shaped” pattern (2 cpd, 180° phase shift between upper and bottom halves) was presented to the other eye (Figs. 2a and 2b).

Stimuli and results for the Experiment 1 condition in which half of one grating was phase-shifted. In this condition, the rivalrous stimuli consisted of an equiluminant grating and an equiluminant tooth-shaped pattern that differed in chromaticity (a, b). The graphs (c–e) show the average duration of exclusive visibility for each of four percepts during 1 min of rivalrous presentation of the stimuli. Each graph presents the results for a different observer. The four percepts, which are illustrated at the bottom, included percepts of the monocular stimuli and percepts with color-binding errors. The heights of the bars do not sum to 60 s of presentation time because other percepts, such as a patchlike or piecemeal grating, were not included in the response options. Error bars are standard errors of the mean based on a single value from each of 3 days; each day's single value is the average duration of exclusive visibility from four counterbalanced runs (see the text).
Within a session, the rivalrous stimuli were presented in counterbalanced runs with the stimuli switched between the left and right eyes. This controlled for eye dominance. Each run began with the presentation to each eye of the circular guideline, the vertical and horizontal nonius lines, and a nonrivalrous achromatic (EES) 8-cd/m2 uniform field in the circular aperture where the rivalrous field would appear subsequently. The observer pressed a button after achieving stable alignment of the nonius lines. Then the rivalrous left-eye stimulus was presented alone for 5 s, followed by the rivalrous right-eye stimulus alone for 5 s. This allowed the observer to identify each monocular stimulus, including its perceived colors, so he or she could report the perceived stimuli during rivalry. When the observer made another button press, the rivalrous stimuli were presented dichoptically for 1 min.
Percepts were measured during the 1-min presentation of the rivalrous stimuli. Observers pressed buttons on a game pad to report their percepts. For example, observers pressed a particular button when they perceived the complete stimulus presented to the left eye (left-eye dominance). They held down this button until the percept changed. When the complete stimulus presented to the right eye was perceived, they pressed a different button. Two additional buttons were assigned for percepts that had color-binding errors (e.g., in Fig. 2, one for a tooth-shaped pattern with both eyes' colors and one for a vertical grating with both colors). Observers were instructed not to press any button if they experienced a different percept, such as a nonhomogeneous, piecemeal, or indistinct field, or a color other than those seen during the preliminary monocular viewing. The percept of binocular color mixture was not explicitly measured because pilot work showed that color mixture did not occur with these stimulus configurations. The percepts assigned to the buttons were salient and easily recognized. The total duration for each percept (exclusive visibility) was the dependent measure, as in other studies of binocular rivalry (Andrews & Blakemore, 2002; Blake, Yu, Lokey, & Norman, 1998; Paffen, Tadin, te Pas, Blake, & Verstraten, 2006). Each condition was run four times on a given day, counterbalancing across eyes for spatial pattern and color (one run with the stimuli in Fig. 1a, one run as in Fig. 1a but with the left- and right-eye stimuli reversed, one run with the stimuli in Fig. 1b, and one run as in Fig. 1b but with the left- and right-eye stimuli reversed). These four runs were repeated three times, each time on a different day.
Results and Discussion
When rivalrous equiluminant gratings differed in both orientation and color, the perceived orientation of the grating alternated between vertical and horizontal (dominance of one orientation or the other). The perceived color of the grating was either magenta and gray or green and gray (single-eye dominance) or magenta and green (a color-binding error). The color-binding error occurred with both perceived orientations of the grating. Figures 1c through 1e present the average duration of exclusive visibility of each percept during 1 min of stimulus presentation, for 3 observers. All observers experienced color-binding errors.
Perceptual color-binding errors were observed also when the difference in form was defined by a phase shift in half of one monocular stimulus (Fig. 2c–2e). In this case, both colors (a binding error) often were perceived in the tooth-shaped pattern and the vertical grating, which were perceived in alternation.
EXPERIMENT 2: COLOR-BINDING ERRORS WITHOUT FORM RIVALRY
Experiment 1 showed that color within a rivalrously suppressed form can appear in a nonretinotopic location of the dominant form, but is form rivalry necessary for color-binding errors? Experiment 2 was conducted to answer this question. The procedure of Experiment 1 was followed, using gratings with only color rivalry. An observer response for color mixture was added, and counterbalancing across eyes now required only two runs within a day because the spatial pattern in the two eyes was the same. For example, in one condition, both eyes saw a 2-cpd vertical equiluminant grating, but the grating was green and gray in one eye and reddish orange and gray in the other eye (Figs. 3a and 3b). The chromaticities that appeared green and reddish orange were [L/(L + M) of 0.611, S/(L + M) of 0.3] and [L/(L + M) of 0.718, S/(L + M) of 0.3], respectively. Color-binding errors were experienced even without form rivalry (Figs. 3c–3e): Observers sometimes perceived a two-color grating with green and reddish-orange stripes. In this condition, binocular color mixture also was seen occasionally in the retinotopic area with rivalrous colors (Ikeda & Sagawa, 1979).
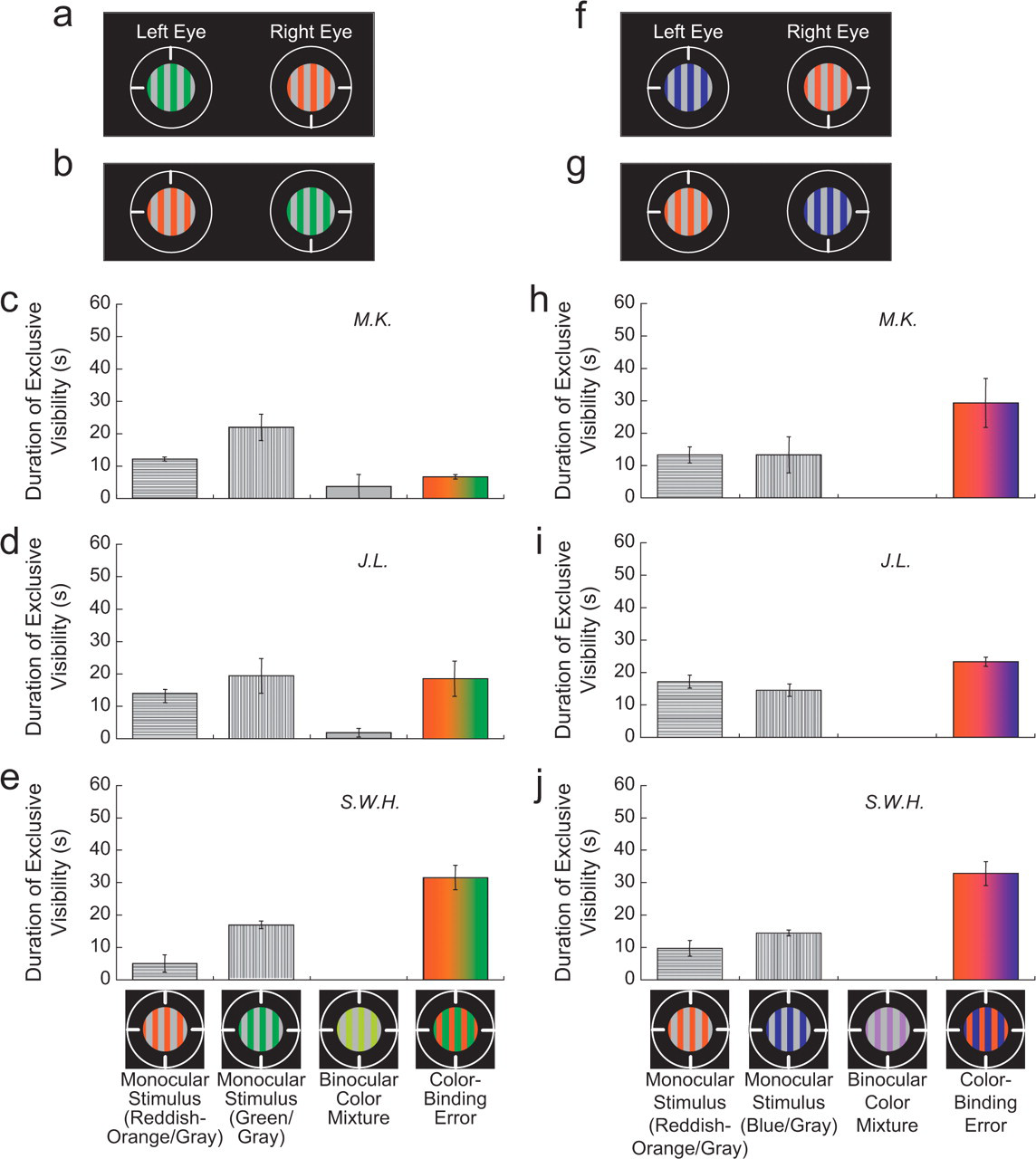
Stimuli and results from Experiment 2. In this experiment, the rivalrous stimuli were vertical gratings that differed in color. In one condition (a, b), the chromatic regions of one stimulus were reddish orange, and the chromatic regions of the other stimulus were green (see the text for precise chromaticities). In the second condition (f, g), the green-and-gray grating was replaced by a blue-and-gray grating (see the text for precise chromaticity). Results for each condition are shown in the graphs below the illustrations of the stimuli. The graphs (c–e, h–j) show the average duration of exclusive visibility for each of four percepts during 1 min of rivalrous presentation of the stimuli. Each graph presents the results for a different observer. The four percepts, which are illustrated at the bottom, included percepts of the monocular stimuli, a percept of binocular color mixture in the area of the rivalrous colors, and percepts with color-binding errors. The heights of the bars do not sum to 60 s of presentation time because other percepts, such as a patchlike or piecemeal grating, were not included in the response options. Error bars are standard errors of the mean based on a single value from each of 3 days; each day's single value is the average duration of exclusive visibility from two counterbalanced runs: the stimuli in (a) and (b) or the stimuli in (f) and (g).
A second condition was the same except that a blue-and-gray grating replaced the green-and-gray grating (Figs. 3f and 3g); the chromaticity that appeared blue was [L/(L + M) of 0.611, S/(L + M) of 3.3]. Again, color-binding errors were common (Figs. 3h–3j); observers often perceived a two-color grating with blue and reddish-orange stripes. In this condition, there was no possibility that the perception of a two-color grating could be due to simultaneous or successive color contrast, because the perceived two-color grating had the colors from both eyes' gratings, rather than a color that could be induced by a blue or reddish-orange monocular stimulus.
EXPERIMENT 3: TESTING THE JIGSAW-PUZZLE HYPOTHESIS
The percept of a two-color pattern in Experiment 1 may seem to imply separate resolution of color and form rivalry. That is, competing forms may be resolved by dominance of one form and suppression of the other, while competing colors are both expressed in the percept. Although the color-binding errors observed in that experiment suggest separate neural representations of form and color, there is an alternative possibility we call the jigsaw-puzzle hypothesis, which was considered in Experiment 3.
The alternative explanation for the results in Experiment 1 is that the color and form of local regions were bound inseparably at an early locus, with the neural representation of the local region wrongly located in the visual percept. According to this alternative, the percept of a two-color grating resulted not from errors in binding color to form, but from a kind of part-binding failure, which is a failure to locate the discrete pieces of the stimulus correctly. Equiluminant stimuli defined by only chromatic differences are susceptible to part-binding failure (Billock & Tsou, 2004).
The jigsaw-puzzle hypothesis is not excluded by Experiment 1 or 2 because the chromatic segments presented to one eye could have replaced the achromatic segments presented to the other eye (e.g., in Fig. 1a, the right eye's green segments could have replaced the left eye's gray segments). This hypothesis was tested using a spatial structure in which the chromatic regions had a different width than the achromatic regions. When the chromatic regions were narrower than the achromatic regions (Fig. 4a), each chromatic region taken as a unit could not cover a complete achromatic region. If the percept of a two-color grating followed from early color-form binding at a local level with subsequent mislocation of local regions, then the achromatic regions, which were wider than the chromatic regions, would not be completely filled with color in this condition. However, if chromatic information was bound incorrectly to an independently defined spatial region in the percept, then the percept would have no achromatic area, regardless of the relative widths of chromatic and achromatic regions.
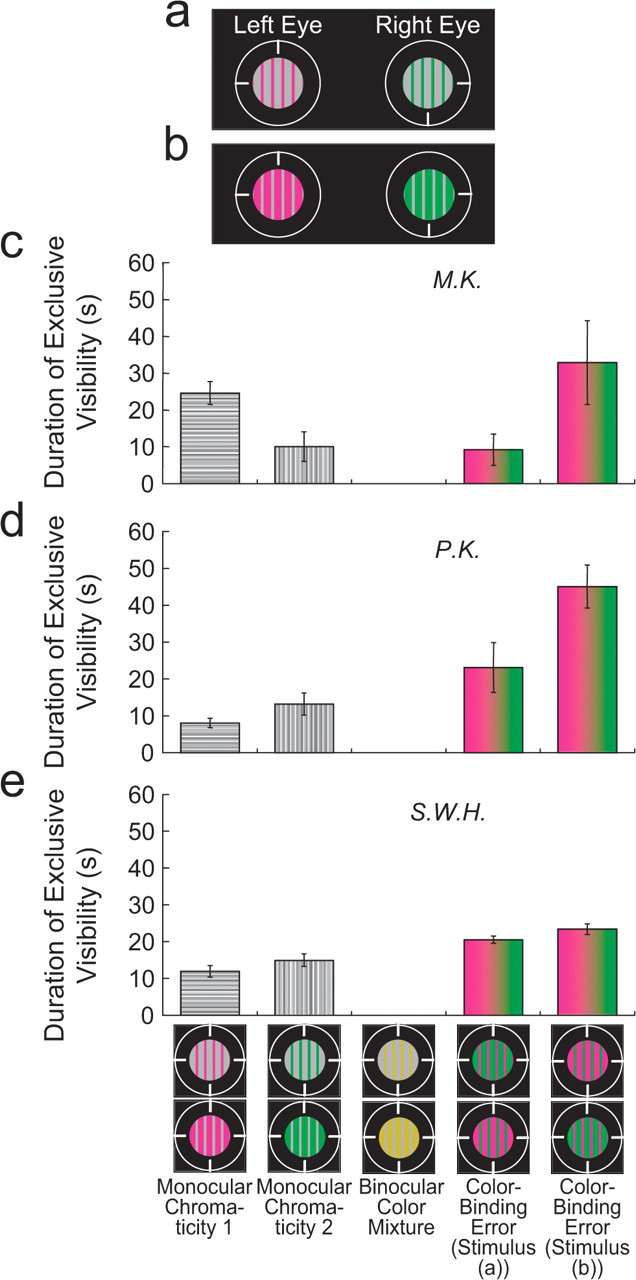
Stimuli and results from Experiment 3. In this experiment, the rivalrous stimuli consisted of vertical gratings that differed in chromaticity and also in whether the chromatic regions were narrower or wider than the achromatic regions (a, b). The graphs (c–e) show the average duration of exclusive visibility for each of five percepts during 1 min of rivalrous presentation of the stimuli. Each graph presents the results for a different observer. The five percepts, which are illustrated at the bottom, included percepts of the monocular stimuli, a percept of color mixture, and percepts with color-binding errors (separately for the two conditions). Error bars are standard errors of the mean based on a single value from each of 3 days. For the first three bars in each graph, each day's single value is the average duration of exclusive visibility from four runs (the stimuli in (a) and (b), and counterbalanced runs with stimuli in the left and right eyes reversed); for the fourth bar in each graph, each day's single value is the average duration of exclusive visibility from two runs (the stimuli in (a) and a counterbalanced run with the left- and right-eye stimuli reversed); for the fifth bar, each day's single value is the average duration of exclusive visibility from the two other runs (the stimuli in (b) and a counterbalanced run with the left- and right-eye stimuli reversed).
Experiment 3 followed the procedure of Experiment 1, but with different stimuli and an added observer response for color mixture. Counterbalancing across eyes required only two runs within a day because the spatial pattern in the two eyes was the same. In one condition of Experiment 3, the chromatic regions were narrower (0.125°) than the achromatic regions (0.25°); in a separate condition, the chromatic regions were wider than the achromatic regions (0.25° and 0.125°, respectively; Figs. 4a and 4b). Figures 4c through 4e report the results graphically. Monocular dominance sometimes occurred, though binocular color mixture of chromaticities in corresponding retinotopic locations was virtually never reported. In both conditions, however, every observer experienced the color-binding error, which covered the complete achromatic area. These results are inconsistent with the jigsaw-puzzle hypothesis. Thus, the percept of both colors within a single grating results from a neural representation for the color from one eye being bound to a nonretinotopic location within a form presented to the other eye.
EXPERIMENT 4: PERCEIVED HUES OF COLOR-BINDING ERRORS
If the percept of a two-color (magenta-and-green) grating in the case of the stimuli illustrated in Figures 1, 2, and 4 results from the intact neural representation of the color from each eye, then the appearance of each color during color-binding errors should be the same as its appearance when nonrivalrous stimuli are used. This prediction was tested by measuring hues perceived with and without color-binding errors.
Stimuli and Procedure
In the critical part of this experiment, two rivalrous 2-cpd vertical gratings, as in Experiment 2, were presented in the top half of the display, and a nonrivalrous stimulus, the matching stimulus, was presented to each eye in the bottom half. The rivalrous stimuli differed only in color (magenta and gray in one eye, green and gray in the other), to maximize the sustained periods with color-binding errors. The sustained percepts of two-color gratings facilitated accurate color settings in the separate matching field. The matching stimuli presented to the two eyes were identical, as in natural binocular viewing, so they were easily fused; they also were 2-cpd vertical gratings (the same size as the rivalrous stimuli) with colored-and-gray stripes. Observers used a game pad to adjust the hue, saturation, and brightness of the colored stripes in the nonrivalrous bottom grating to match a perceived color within the top grating (asymmetric color matching).
The matching protocol included three blocks, with the rivalrous stimuli presented in only the second. In the first block, the top stimuli were identical, rather than rivalrous. Observers matched the color in the magenta-and-gray or green-and-gray gratings (three trials per color). Matches to each color were averaged. Next, the rivalrous stimuli were presented. First, with the magenta-and-gray grating presented to one eye and the green-and-gray grating presented to the other eye, the observer made three matches to each color. Then, the eye stimulated by each grating was reversed, and the observer made three additional matches to each color. This controlled for eye dominance. Observers matched the appearance of the colors only when the two-color (color-binding error) grating was perceived. The six matches to each color were averaged. Observers were instructed not to attempt a match when the percept of the two-color grating disappeared, but to try again when that percept returned. In the third block, matches to the nonrivalrous gratings were repeated as a check on whether matches to the two-color grating were affected by chromatic adaptation during extended viewing.
Results and Discussion
If the same neural representation determined the perceived color regardless of whether the percept had a color-binding error due to rivalry or there was no binding error (nonrivalrous stimuli), then all the matches should have been the same. As Figure 5 shows, the matches from the three blocks were very similar and often overlapping. Thus, the appearance of the hues was virtually the same during color-binding errors as during normal, nonrivalrous viewing. This result is consistent with the neural representation for color in the binding-error percept being the same as the representation in normal viewing without rivalry.
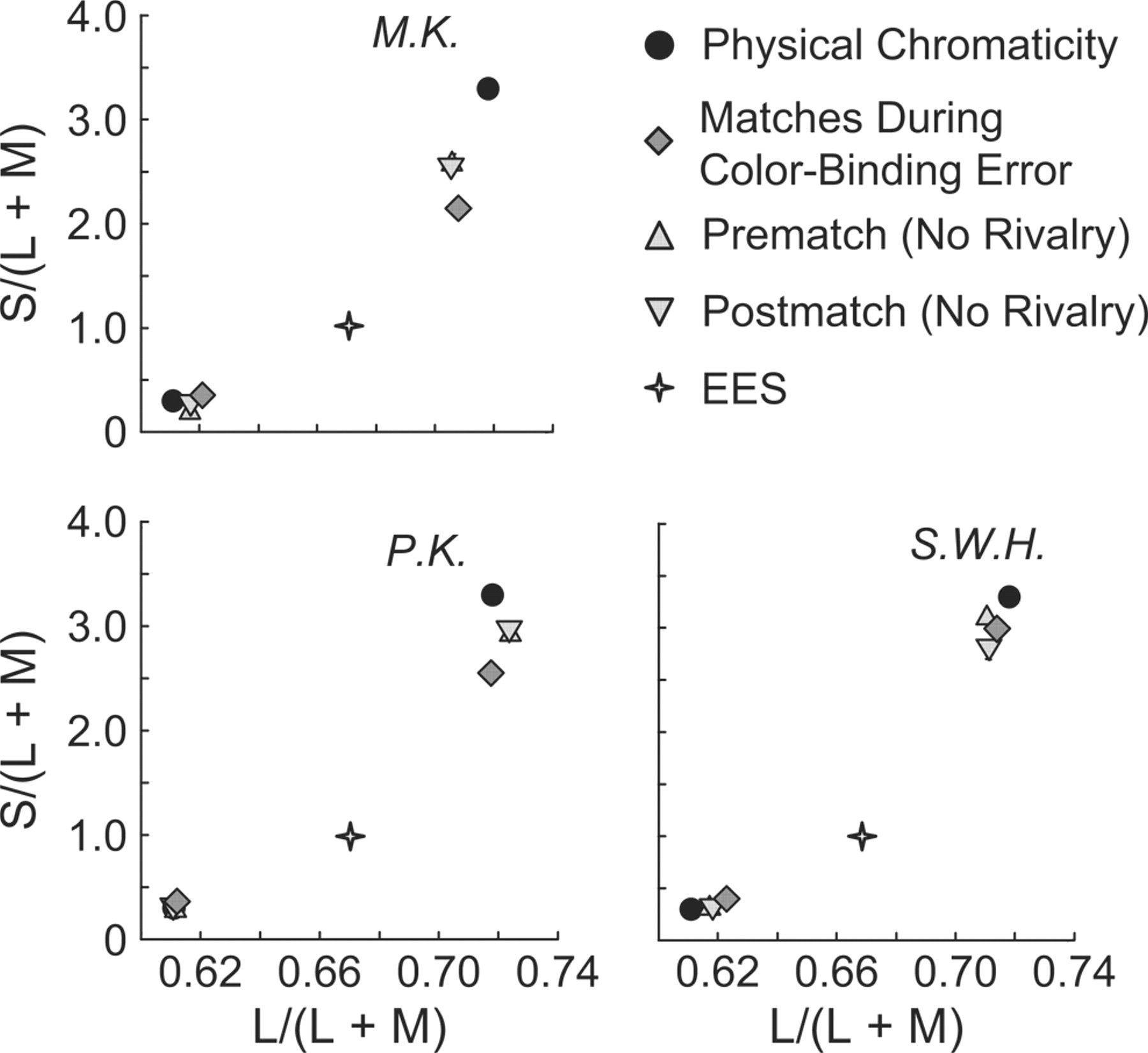
Results from Experiment 4: color appearance measured by asymmetric color matching of the chromaticities shown in Figure 1. The axes represent a cone-stimulation space (MacLeod & Boynton, 1979). Matches were made while a two-color grating was perceived during binocular rivalry (color-binding error). Observers also matched individual chromaticities presented to both eyes (no rivalry), both before and after the matches during color-binding errors. The equal-energy-spectrum (EES) chromaticity of the gray stripes in the gratings is also shown. Each panel presents results for a different observer.
These measurements of color appearance corroborate the conclusion that the perceived two-color gratings did not result from classical color contrast. A greenish inducing background causes nearby achromatic light to appear reddish (Valberg, 1974); similarly, a reddish background causes nearby light to appear greenish. Color contrast, however, induces far weaker color shifts into achromatic regions than shown in Figure 5 (Ware & Cowan, 1982). Moreover, if the perception of two-color gratings was caused by color contrast, then nonrivalrous magenta-and-gray gratings presented to both eyes (or green-and-gray gratings presented to both eyes) also would produce the percept of a two-color magenta-and-green grating, but this was never seen.
GENERAL DISCUSSION
The color-binding errors in these experiments demonstrate that color can be represented separately from orientation, spatial phase, and spatial frequency. Alternative explanations were excluded for the percept of a two-color grating arising when rivalrous stimuli, each having a different single color, were presented: Neither the jigsaw-puzzle hypothesis (part-binding failure) nor chromatic contrast can explain the results. Optical misalignment or vergence errors are not viable explanations for the two-color gratings perceived in Experiment 1 (see Figs. 1 and 2) because such errors cannot cause the chromatic regions of one eye's stimulus to align with the achromatic regions of the other eye's stimulus.
The color-binding errors reported here implicate a neural binding process that combines separate representations of color and form. The existence of a distinct process for binding color and form has been challenged by physiological studies showing that many color-selective cells in V1 and V2 are tuned also to orientation (Johnson, Hawken, & Shapley, 2001; Kiper, Fenstemaker, & Gegenfurtner, 1997). These multiselective cells conceivably could solve the binding problem for color and form with neural responses that are jointly selective for chromaticity, spatial frequency, and orientation. An extreme view based on such cells is that color and form are not separable even at an early stage of visual processing, so that “a binding problem does not exist for the representation of color and orientation in the visual cortex” (Friedman, Zhou, & von der Heydt, 2003, p. 610).
Joint selectivity for form and color within a single cell, however, does not imply that perceived form and hue are inseparably represented together. Cells driven by purely chromatic (equiluminant) gratings might determine percepts of only spatial frequency and orientation, not hue. Psychophysical studies support the view that cells with multiple selectivity contribute to form processing (Clifford, Spehar, Solomon, Martin, & Zaidi, 2003): The perceived orientation of a central grating is most strongly affected by the orientation of a surrounding grating when the two gratings vary along the same chromatic axis. This finding, however, sheds no light on the role of these cells in color perception.
The color-binding errors reported here reveal an unexpected fate for chromatic neural signals during binocular rivalry. Theories of rivalry posit that a stimulus disappears from the conscious percept as a result of suppression at some neural locus (Blake & Logothetis, 2002). Neural suppression during rivalry has been reported at several levels of the visual processing stream, including the lateral geniculate nucleus (Haynes, Deichmann, & Rees, 2005; Wunderlich, Schneider, & Kastner, 2005), cortical area V1 (Lee & Blake, 2002; Polonsky, Blake, Braun, & Heeger, 2000; Tong & Engel, 2001), and beyond (Logothetis, Leopold, & Sheinberg, 1996; Sheinberg & Logothetis, 1997; Tong, Nakayama, Vaughan, & Kanwisher, 1998). In comparison, rivalrous colors neither suppressed nor mixed with each other in the present study, but instead were both distinctly perceived, in separate locations.
Stirling (1901) was aware of the dissociation of color and form processing during dichoptic presentation of two different forms and chromaticities. He dichoptically presented two differently colored postage stamps with slightly different forms (faces), and occasionally one or the other face was perceived with a mixed color. In his study, the form information from one eye was dominant, but chromatic information from the two eyes was mixed. This result was later confirmed with more rigorous methods (Holmes, Hancock, & Andrews, 2006; Lange-Malecki, Creutzfeldt, & Hinse, 1985). Another example of the same dissociation is an occasional binding error that occurs when rivalrous stimuli of different colors and orientations are viewed following a binocular “priming” grating with the orientation of one eye's rivalrous stimulus and the color of the other eye's rivalrous stimulus (Holmes et al., 2006): The percept of the rivalrous stimuli after such priming can be a grating with the color from one eye and orientation from the other eye, as a result of separate dominance of one eye's orientation and the other eye's color. The results presented here, however, are fundamentally different from previous results because competing chromatic signals from the two eyes were resolved with a percept having the colors from both eyes. The two colors were seen simultaneously within distinct regions that did not correspond to the retinal location of chromatic stimulation. These errors in binding color to form show that rivalrous, competing chromatic signals from the two eyes have neural representations of hue that are fully available to be bound to different parts of the unified percept.
Footnotes
Acknowledgements
This work was supported by National Institutes of Health Grant EY-04802. We thank Randolph Blake and Howard Nusbaum for comments on an earlier draft.