
Abstract
According to widely held views in cognitive science harking back to David Hume, causality cannot be perceived directly, but instead is inferred from patterns of sensory experience, and the quality of these inferences is determined by perceivable quantities such as contingency and contiguity. We report results that suggest a reversal of Hume's conjecture: People's sense of time is warped by the experience of causality. In a stimulus-anticipation task, participants' response behavior reflected a shortened experience of time in the case of target stimuli participants themselves had generated, relative to equidistant, equally predictable stimuli they had not caused. These findings suggest that causality in the mind leads to temporal binding of cause and effect, and extend and generalize beyond earlier claims of intentional binding between action and outcome.
Cognitive scientists usually agree that the human sensory system is not equipped to perceive causal relations directly, but instead infers them from information available to the senses, such as contingency and contiguity (Shanks, Holyoak, & Medin, 1996). Even though some situations give rise to immediate and irresistible visual causal impressions (Michotte, 1946/1963), recent evidence suggests that such visual processes recruit inferential processes (Scholl & Nakayama, 2002; Scholl & Tremoulet, 2000), albeit not necessarily the kind of deliberative processes involved, for instance, in determining whether smoking causes lung cancer (Cheng, 1997). This inferential stance dates back to David Hume's (1739/1888) famous conjecture that a cause is “an object precedent and contiguous to another and so united with it, that the idea of the one determines the mind to form the idea of the other” (p. 170).
Cognitive constructs such as attitudes, prejudice, and causal beliefs have long been known to bias people's judgments of probability and frequency (Tversky & Kahneman, 1974). Similarly, remembrance for past events is vulnerable to distortions because of the constructive (as opposed to reproductive) nature of memory (Faro, Leclerc, & Hastie, 2005; Suddendorf & Corballis, 2007). More specifically, Faro et al. demonstrated that people's estimates of how much time passed between various historic events are determined by underlying causal beliefs, such that events that are thought to be causally related are judged to be closer together in time than are unrelated events.
Here we demonstrate that causal beliefs not only exert influence over other cognitive constructs or memory, but also lead to profound changes in perception and action. Our research was inspired by an earlier demonstration (Haggard, Clark, & Kalogeras, 2002) of systematic changes in event perception. Using the Libet clock method (Libet, Gleason, Wright, & Pearl, 1983), Haggard et al. showed that in subjective awareness, voluntary actions and resultant consequences were shifted closer together in time than were unrelated events. Participants monitored a fast-moving clock and had to report the position of the clock hand when certain events occurred. Comparison of reported with actual clock-hand position allowed an appraisal of judgment error. Participants reported that their actions (key presses) occurred later when those actions subsequently caused an effect (a tone), compared with when they had no consequence; likewise, tones that had been preceded by causal key presses were judged to have occurred earlier than tones that occurred on their own. Haggard et al. interpreted this apparent perceptual attraction as evidence for intentional binding: The brain “binds intentional actions to their effects to construct a coherent conscious experience of our own agency” (p. 385).
The results we report here advance beyond this finding in two important ways. First, our method relied on behavioral data (motor planning), rather than on subjective reports or estimates, and the elimination of (postperceptual) response biases allowed a more convincing appraisal of attraction effects. Second, we show that temporal binding serves a more fundamental purpose than supporting the experience of intentionality: It is driven by impressions of causality, even when intentionality and agency are controlled for. Thus, Hume's famous conjecture that perceptual experiences of temporal contiguity lead to mental constructs of causality appears to describe one half of a bidirectional relation: Causal inferences analogously lead the sensorimotor system to perceive temporal contiguity.
EXPERIMENT 1
Each participant in Experiment 1 performed a stimulus-anticipation task in two conditions, baseline and causal control. Each condition was run with three target intervals (500, 900, and 1,300 ms) and comprised three phases (see Fig. 1).
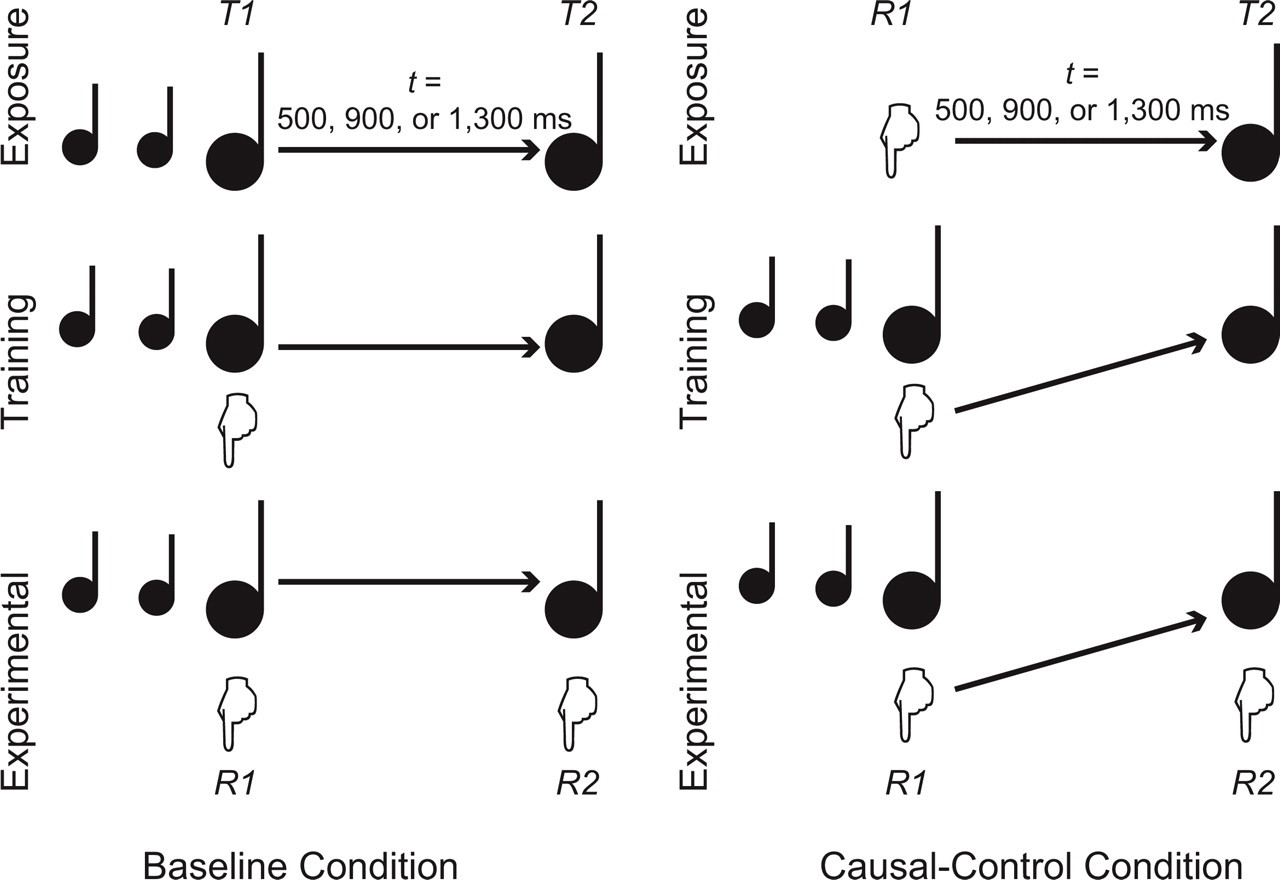
Method used in Experiment 1. The task was to synchronize two responses, R1 and R2, to two targets, T1 and T2, respectively. In the baseline condition, T1 and T2 were separated by an interval (t) of 500, 900, or 1,300 ms. Participants were first exposed to this interval (exposure phase), then synchronized R1 to T1 (training phase), and finally synchronized both R1 and R2 to T1 and T2 (experimental phase). In the causal-control condition, R1 generated T2 after an interval (t) of 500, 900, or 1,300 ms (exposure phase). Participants then synchronized R1 to T1 (training phase) and finally R1 and R2 to T1 and T2 (experimental phase).
On each trial in the exposure phase of the baseline condition, participants were exposed to two 100-ms target tones (T1 and T2) separated by an interval of 500, 900, or 1,300 ms. T1 was a 1000-Hz pure tone, and T2 was a 1100-Hz pure tone. T1, which marked the beginning of the interval, was always preceded by a go signal consisting of two 600-Hz preparatory tones (P1 and P2), each lasting 100 ms; there was a 200-ms interval between P1 and P2 and between P2 and T1. In the baseline training phase, participants were instructed that on each trial they should synchronize a key-press response (R1) to T1, which was still preceded by P1 and P2, and followed by T2 (as in the exposure phase). In the baseline experimental phase, participants both synchronized R1 to T1 and synchronized another key press (R2, on a different button) to T2.
In the causal-control condition, the exposure phase taught participants that R1 triggered T2 after one of the three target intervals (500, 900, or 1,300 ms). Participants learned this causal relation in a free-operant manner: They were free to press the key whenever they wanted, and did not need to respond to any signals. In the training phase, participants had to synchronize R1 to T1, just as in the baseline condition, and R1 triggered T2 after the same interval. In the experimental phase, participants had to synchronize R1 to T1 and R2 to T2; again, the timing of T2 was triggered by R1.
Thus, the experimental phase in the two conditions involved the same intentional actions, emitted in anticipation of the same two target signals. Critically, however, R1 was causally linked to T2 only in the causal-control condition. We recorded the absolute timing of R1 and R2 in the experimental phase of each condition and subtracted from these values the actual timing of the onset of T1 and T2. This provided an objective index of response accuracy, with a negative number indicating that a response preceded its target, and a positive number indicating that a response occurred after its target. Our prediction, based on the results of Haggard et al. (2002), was that R2 would occur earlier (relative to T2) in the causal-control condition than in the baseline condition, reflecting participants' subjective impression that T2 appeared earlier when it was caused by R1 than when it was not.
We also analyzed accuracy of R1 with respect to T1. According to the causal-binding hypothesis, causal actions and their outcomes are fused together in time, such that the cause appears to occur subjectively later than it does in a noncausal condition, and the effect appears to occur subjectively earlier; this results in an overall shortening of the interevent interval. If participants perceived a causal R1 as later than a baseline R1, how would this affect their behavior when they tried to synchronize R1 to an external signal? We suggest that R1 would be executed objectively earlier in the causal-control condition: Because causal binding would shift awareness of R1 forward in time (nearer to its effect), a veridically synchronized R1 might subjectively appear to be “too late”; to correct this, participants might adjust the timing of R1 backward in time, so that R1 would occur objectively earlier than T1, but subjectively in synchrony with it.
Method
Participants
Thirty-four Cardiff University undergraduates (28 female, 6 male) participated in return for course credit or £4.
Materials and Apparatus
The experiment was presented individually to participants on two iMac computers, located in separate cubicles. Each computer was running PsyScope (Cohen, MacWhinney, Flatt, & Provost, 1993) and was connected to a NewMicros (Dallas, TX) PsyScope button box. Two 100-ms 600-Hz pure tones were used as the go signals (P1 and P2), a 100-ms 1000-Hz pure tone was used for T1, and a 100-ms 1100-Hz pure tone was used for T2. All instructions were presented on the computer screen, and the experimenter clarified the procedure in case participants had questions. The timing of R1 and R2 (presses of separate keys on the button box) was recorded with millisecond accuracy. The target interval (between T1 and T2 in the baseline condition, between R1 and T2 in the causal-control condition) on each trial was 500 ms, 900 ms, or 1,300 ms.
Design and Procedure
Condition (causal-control or baseline) and target interval (500, 900, or 1,300 ms) were varied within subjects. The experiment was preceded by a preparatory phase during which participants learned to synchronize R1 to T1. To this end, participants were presented with 10 trials on which P1 was followed 200 ms later by P2, which was followed 200 ms later by T1. Participants were instructed to synchronize R1 with T1. In this training phase, T2 was never presented.
After this preparatory phase, the experiment proper began. Each participant completed the task in both the baseline and the causal-control conditions, and trials were blocked so that all trials of one condition were completed before the other condition began. We counterbalanced the condition that was presented first across participants. Each condition was repeated three times (i.e., three runs), once with each of the target intervals (500, 900, and 1,300 ms); the order of intervals within a condition was random. One run through a particular condition-interval combination consisted of three phases: exposure, training, and experimental.
In the baseline condition, the exposure phase consisted of 15 presentations of the target interval preceded by the go signal (i.e., P1, P2, T1, and T2). Participants were not required to make any responses at this stage, and merely paid attention to the interval between T1 and T2. After a random delay of 1,000 to 2,000 ms following the end of T2, the next trial commenced. In the training phase, participants were again presented with 15 complete sequences of tones, but this time were instructed to perform R1 in synchrony with T1. Finally, the experimental phase consisted of 30 presentations of the P1-P2-T1-T2 sequence, and participants had to synchronize R1 and R2 to T1 and T2, respectively.
In the causal-control condition, the exposure phase was a free-operant training phase in which no go tones were present. Instead, participants were requested to perform R1 at a time of their choosing and thus to cause T2 to appear after the relevant interval. Once they had done this 15 times, they proceeded to the training phase, which consisted of 15 presentations of the P1-P2-T1 sequence. Participants were instructed to synchronize R1 with T1. R1 triggered T2 after the same interval as in the exposure phase, and participants were encouraged to occasionally “fail” to synchronize R1 to T1, and to observe that this meant an earlier or later delivery of T2 (relative to T1). Finally, the experimental phase consisted of 30 trials of the P1-P2-T1 sequence. Participants were now instructed to synchronize R1 to T1 and R2 to T2, which, they were reminded, was caused by R1.
The experimental phase in both the causal-control and the baseline conditions employed a feedback mechanism: If participants failed to perform the relevant response, an error signal indicating the miss appeared 1,000 ms after the relevant target. During all trials, a diagrammatic representation of the task was displayed. P1, P2, and T1 were delivered from a speaker on the participant's left, and T2 was delivered from a speaker on the right.
Results and Discussion
For each participant, we calculated the mean accuracy scores for R1 and R2 for each interval in each of the two conditions. Each participant thus provided 12 data points for further analysis: one accuracy score for R1 and one accuracy score for R2 per combination of target interval and condition. Figures 2 and 3 display the accuracy scores for R2 and R1, respectively. Inspection of Figure 2 shows that R2 was consistently executed earlier (relative to T2) in the causal-control condition than in the baseline condition. A 3 × 2 repeated measures analysis of variance (ANOVA) with the factors interval and condition confirmed this observation. The effect of condition was highly significant, F(1, 33) = 11.22, MSE = 3,812.31, p rep = .98; neither the effect of interval, F(2, 66) = 2.39, MSE = 7,271.16, nor the interaction of condition and interval, F(2, 66) = 0.31, MSE = 2,619.89, was significant. Figure 3 suggests that R1 was also shifted in the causal-control condition, compared with the baseline condition, and that it too was executed earlier during the causal-control condition. An ANOVA corroborated this observation, yielding a significant effect of condition, F(1, 33) = 14.35, MSE = 1,044.99, p rep = .99. The effect of interval was also significant, F(2, 66) = 17.85, MSE = 1,017.20, p rep = .99; the interaction was not, F(2, 66) = 1.26, MSE = 743.41.
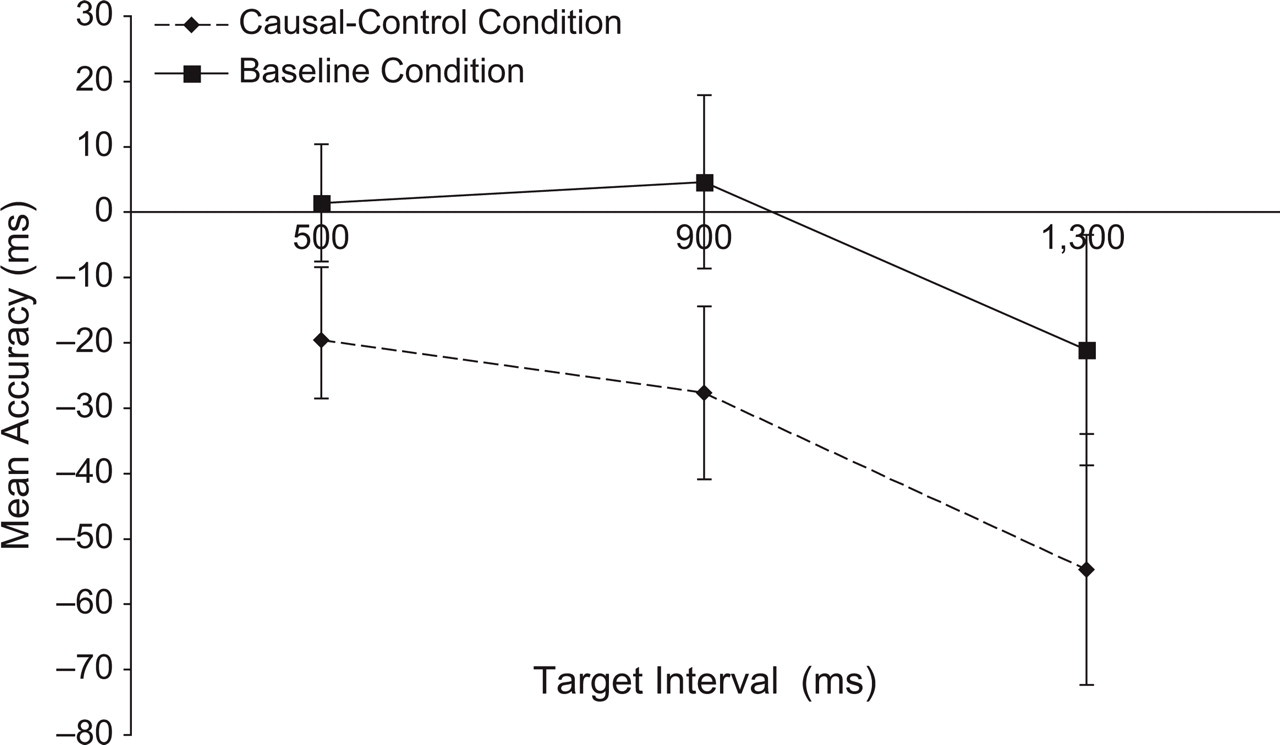
Results from Experiment 1: mean accuracy scores for the second response (R2) as a function of target interval (500, 900, or 1,300 ms) and condition (baseline or causal-control). Accuracy was calculated relative to the onset of the second target (T2). Negative numbers indicate that R2 was made before T2. Error bars indicate standard errors.
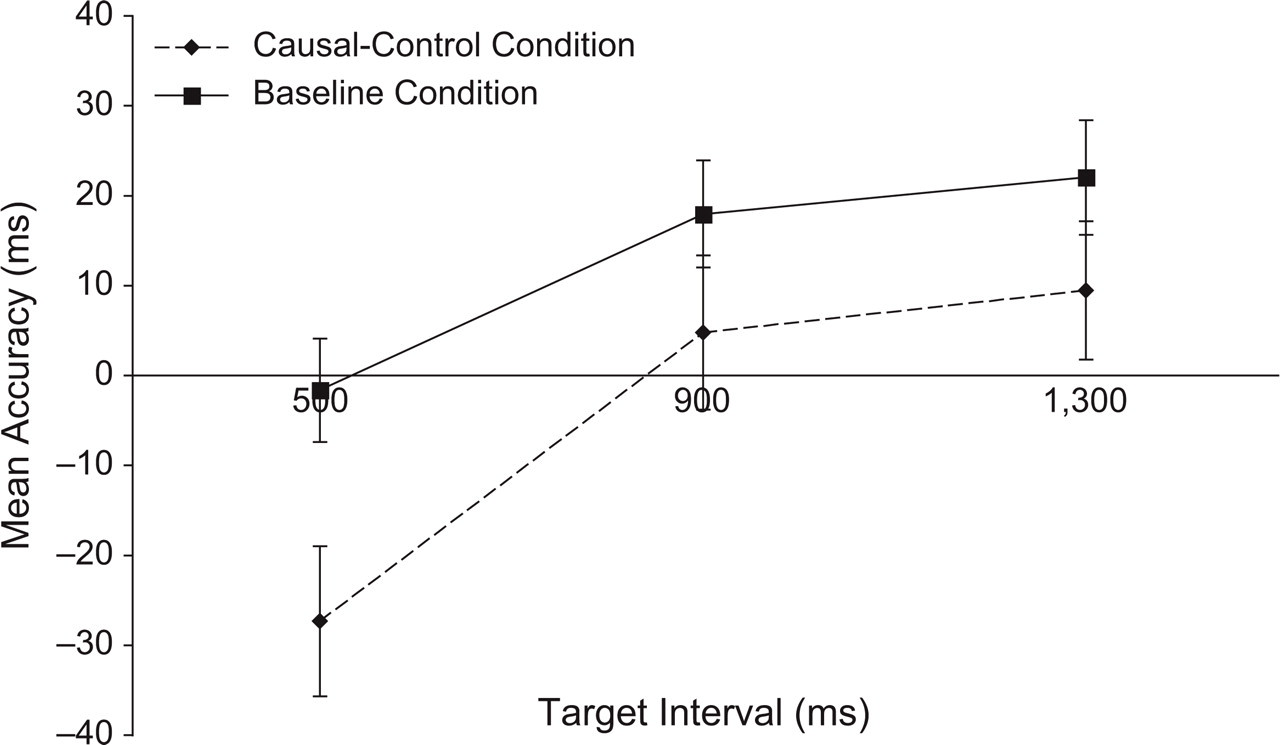
Results from Experiment 1: mean accuracy scores for the first response (R1) as a function of target interval (500, 900, or 1,300 ms) and condition (baseline or causal-control). Accuracy was calculated relative to the onset of the first target (T1). Negative numbers indicate that R1 was made before T1. Error bars indicate standard errors.
Both aspects of our predictions were thus supported. First, participants displayed evidence of causal binding by executing a response aimed to anticipate a target stimulus earlier if they had caused this stimulus than if they had not. This result suggests an earlier awareness of the stimulus in the causal-control condition compared with the baseline condition. Second, participants displayed evidence of causal binding by executing an action later in reference to an external signal if that action subsequently produced a stimulus than if it did not. This result suggests delayed awareness of the action in the causal-control condition compared with the baseline condition.
EXPERIMENT 2
Even though we strove to make the causal-control and baseline conditions absolutely identical on the surface, save for the causal relation between R1 and T2 in the former, the results of Experiment 1 could possibly be explained without drawing on the presence of causality because of a potential confound. In order to realize the causal connection, we tied the delivery of T2 to R1. This means that the R1-T2 interval was fixed, and therefore highly predictable in the causal-control condition. In contrast, in the baseline condition, the timing of T2 was tied to T1 and completely independent of R1, as would be required to rule out a causal connection. This meant, however, that participants' experience of the R1-T2 interval could have been considerably more variable in the baseline condition than in the causal-control condition: Unless they timed R1 perfectly to T1 on each occasion, they would have experienced a slightly different R1-T2 interval on each trial.
This difference in variability could have contributed to a stronger association between R1 and T2 in the causal-control condition than in the baseline condition. If temporal binding results from an association between action and outcome (Haggard et al., 2002), the effects we observed could be explained merely by a difference in variability of experience, without drawing on higher-level concepts of causality. We sought to rule out this alternative explanation in Experiment 2 by modifying the causal-control condition so that T2 occurred after the same fixed interval relative to T1 as in the baseline condition (which in turn would make the R1-T2 interval equally liable to variation in the two conditions). We preserved the causal link by delivering T2 only if R1 was made within a 300-ms interval centered around T1: If the participant pressed the key at any point within that window, T2 was delivered at the relevant interval after T1; if he or she pressed the key outside this window (too early or too late), T2 was withheld. This causal connection was explicitly mentioned to participants, and they were encouraged to try it out during the training phase, that is, to deliberately execute R1 out of sync with T1, to see that no T2 would occur. Figure 4 illustrates the procedure.
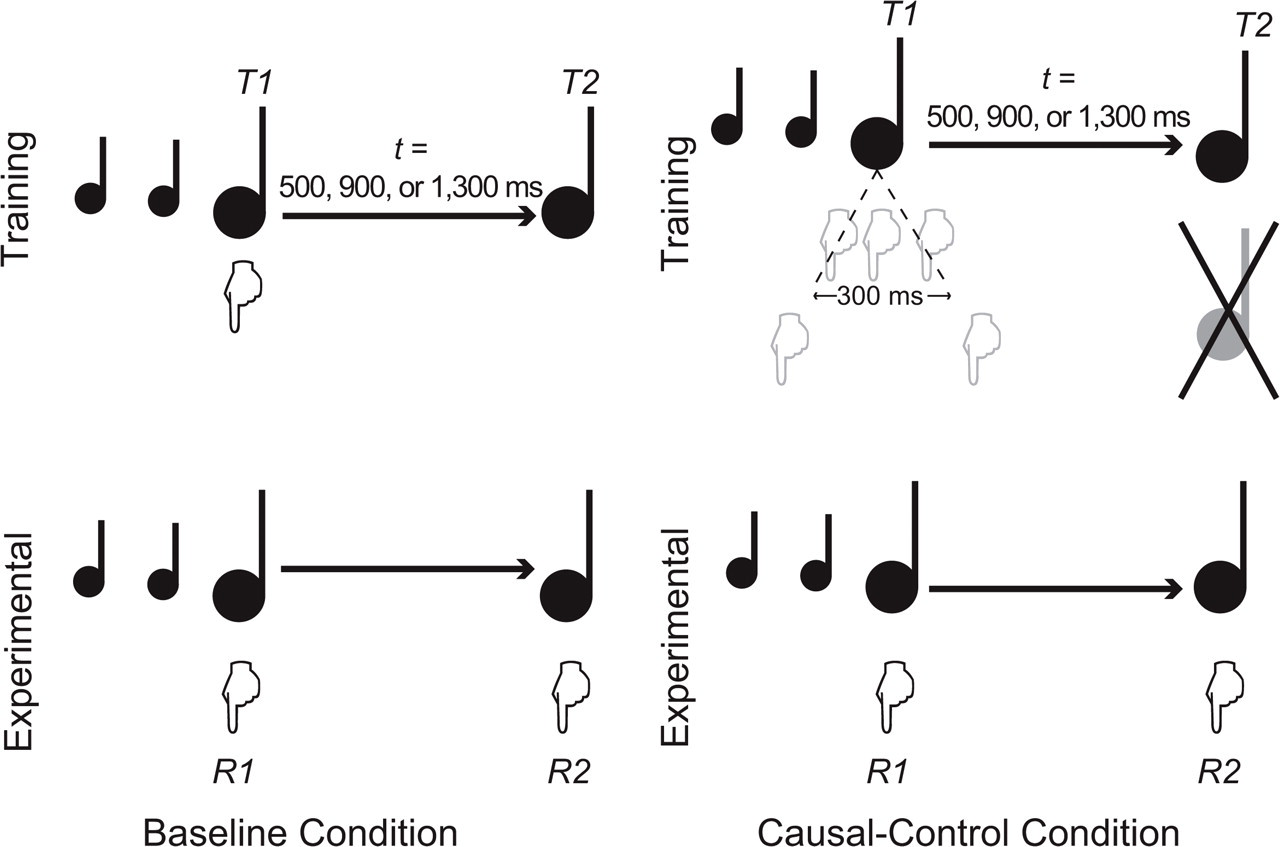
Method used in Experiment 2. As in Experiment 1, the two responses, R1 and R2, had to be synchronized to the two targets, T1 and T2, respectively. In the baseline condition, T1 and T2 were separated by an interval (t) of 500, 900, or 1,300 ms. Participants first synchronized R1 to T1 (training phase) and then synchronized both R1 and R2 to T1 and T2 (experimental phase). In the causal-control condition, participants learned that if R1 was synchronized within a 100-ms window around T1, T2 would occur at an interval (t) of 500, 900, or 1,300 ms after T1; if R1 was not executed within this window, nothing would happen (training phase). Participants then synchronized both R1 and R2 to T1 and T2 (experimental phase).
Method
Participants
Thirty-five Cardiff University undergraduates (29 female, 6 male) participated in return for course credit or £4.
Materials, Design, and Procedure
The following changes relative to Experiment 1 were made: First, the exposure phase was removed from both conditions. Second, the number of trials was increased to 30 in the training phase and to 60 in the experimental phase. Third, T2 was linked to the start of T1 in the causal-control condition, so that T2 appeared after the same fixed interval, as it did in the baseline condition. However, to conserve the causal relationship between R1 and T2, we defined a “critical zone” around T1 that began 100 ms before the start of T1 and extended until 100 ms after the end of T1. Because T1 lasted 100 ms, the critical zone was 300 ms, centered around T1. If R1 was executed within this 300-ms period, T2 was delivered (timing fixed relative to T1), but if R1 occurred outside this period (or not at all), the presentation of T2 was canceled.
Results and Discussion
Figures 5 and 6 show the accuracy scores for R2 and R1, respectively. As the graphs show, the results were markedly different from those of Experiment 1. First, there were no clear differences in accuracy of R2 between the causal-control and baseline conditions. Second, the pattern of results for R1 was distinctly reversed compared with Experiment 1; that is, in Experiment 2, R1 occurred reliably later during the causal-control condition relative to the baseline condition. Two separate repeated measures ANOVAs corroborated these impressions. For R2, the effect of condition was not significant, F(1, 34) = 0.46, MSE = 5,666.53, n.s., p rep = .50; the effect of interval and the interaction of condition and interval were also not reliable. For R1, the main effects of both condition, F(1, 34) = 16.46, MSE = 921.36, p rep = .99, and interval, F(2, 68) = 15.11, MSE = 490.38, p rep = .99, were significant; the interaction was not significant.
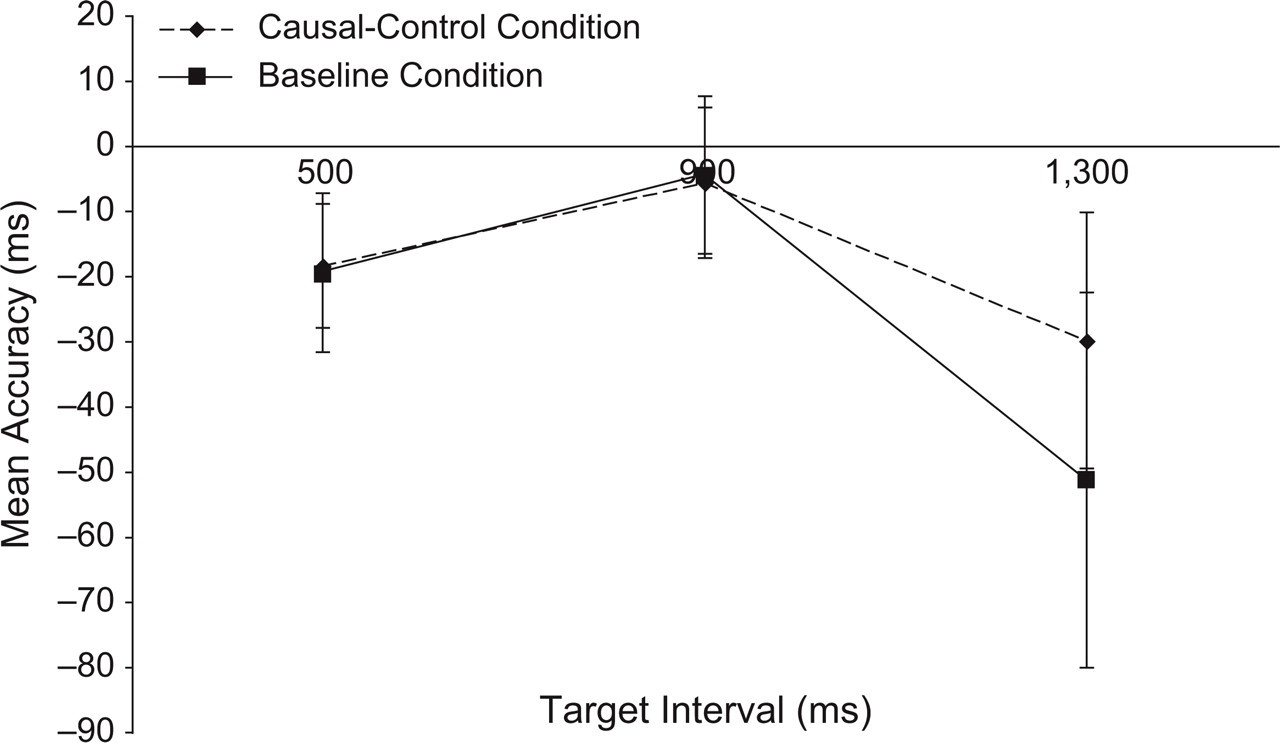
Results from Experiment 2: mean accuracy scores for the second response (R2) as a function of target interval (500, 900, or 1,300 ms) and condition (baseline or causal-control). Accuracy was calculated relative to the onset of the second target (T2). Negative numbers indicate that R2 was made before T2. Error bars indicate standard errors.
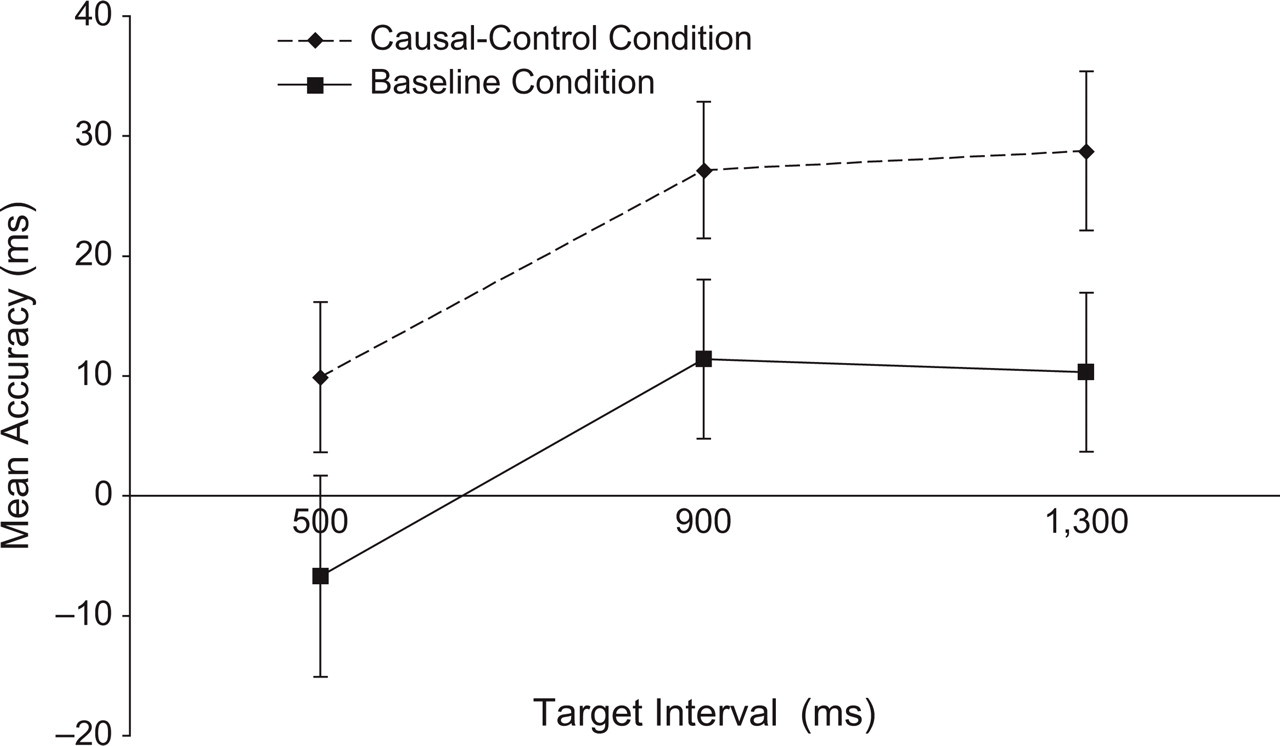
Results from Experiment 2: Mean accuracy scores for the first response (R1) as a function of target interval (500, 900, or 1,300 ms) and condition (baseline or causal-control). Accuracy was calculated relative to the onset of the first target (T1). Negative numbers indicate that R1 was made before T1. Error bars indicate standard errors.
On first impression, these results might appear to contradict Experiment 1, but closer reflection suggests that they provide even stronger evidence for causality-induced temporal binding. Let us begin our analysis with R1. Causal binding suggests that R1 would be subjectively perceived as later in the causal-control condition than in the baseline condition. In Experiment 1, this meant that R1 was emitted earlier in the causal-control condition, so as to preserve the subjective impression of synchrony with T1. In Experiment 2, however, this behavior would have created an objectively longer R1-T2 interval, because T2 was temporally fixed to T1, rather than R1. This would have clearly undermined causal binding, which serves to shorten the subjective impression of this interval. Therefore, our participants shifted R1 forward in time, so as to bring it closer to its effect, T2, which was temporally fixed despite the causal connection. In other words, our participants “acted out” causal binding themselves, by shifting their R1 in time. Apparently, this objectively later R1 still satisfied their impression of synchrony with T1. Interestingly, perceptions of T2 were not shifted. Participants must have realized that T2 was timed relative to T1, and therefore did not shift R2. Thus, causal binding under these circumstances was driven solely by shifts of the causal action toward a temporally fixed outcome.
GENERAL DISCUSSION
Our experiments compared two conditions that were absolutely identical in surface appearance and in their perceptual and motor demands. The difference between the conditions was that R1 was mentally linked to T2 in the causal-control, but not the baseline, condition. This causal link was established before the experimental phase during which measures were taken; it would have been impossible for an onlooker to distinguish the conditions in the experimental phase. Nonetheless, the mere presence of causality in the mind led to powerful and reliable shifts in event perception and motor planning. Note that the causal-control and baseline conditions were matched in the degree of intentional action control they recruited: Both required the conscious planning of motor responses to target signals. The presence of a causal connection in the former but not the latter condition resulted in pronounced temporal binding. Our results indicate that the mental construct of causality—over and above an experience of intentionality in action control—leads to changes in event perception and motor timing in line with one of Hume's (1739/1888) principles of causality: Events that are known to be causally related are experienced as closer in time than unrelated events. “Intentional binding” thus might be a misnomer; the label “causal binding” might be more appropriate.
How do these results fit in with the large body of research on time perception? Typically, subjective experience of time is viewed as a function of attention (e.g., Zakay & Block, 1997): According to the attentional-gate theory of timing, an arousal-regulated pacemaker emits pulses, which are accumulated by a counter; critically, accumulation is regulated by an attentional gate, such that attention to passing time opens the gate, whereas attention to other events (divided attention) permits fewer pulses to pass through the gate. It is hard to see how an attentional model could account for our results, however: The baseline and causal-control conditions were perfectly matched in terms of attentional demands; both required exactly the same two motor responses in anticipation of exactly the same two external signals. The critical difference was whether or not the first response was causally linked to the second target. One might, of course, speculate that the mere presence of causality draws attention away from passing time (perhaps toward the constituent events, cause and effect), and this is an avenue we will pursue in future research.
Our results can also be considered from the perspective of event predictability. Pariyadath and Eagleman (2007) recently demonstrated that duration judgments vary as a function of predictability, over and above attention, such that predictable events are judged to be shorter than unpredictable ones. For example, an oddball stimulus is judged to have a longer duration than its companions in a series of repeating stimuli. Moreover, the first of a sequence of numbers is judged longer both if the subsequent numbers are repetitions of the first number and if they form a predictable sequence (1, 2, 3, 4, 5), but not if they form an unpredictable, random sequence. Thus, according to Pariyadath and Eagleman, predictability leads to a compression of subjective time.
Our stimuli were equally repetitive and predictable in the two conditions, however. If anything, the baseline condition was more predictable than the causal-control condition, because it unfolded exactly the same way every time, independently of the participant's actions; in contrast, the causal-control condition harbored an element of uncertainty in that failure to emit R1 on time resulted in correspondingly earlier or later delivery of T2 (Experiment 1) or cancellation of T2 (Experiment 2). The interevent intervals themselves, however, did vary in predictability. In Experiment 1, the T1-T2 interval was constant and thus predictable in the baseline, but not the causal-control, condition. Conversely, the R1-T2 interval was constant during the causal-control, but not the baseline, condition. If constancy implies a shortening of duration perception, as Pariyadath and Eagleman (2007) suggested, and if participants based their timing of R2 on their subjective impression of the R1-T2 interval, this could explain the shifts in R2 in the causal-control condition. It could also explain why no such shifts occurred in Experiment 2, in which the predictability of the R1-T2 interval was identically variable in the two conditions. It is doubtful, however, that in the baseline condition participants based their execution of R2 on their estimation of the R1-T2 interval. Why should an external event (T2) be timed relative to an earlier action that is known to bear no relation to it, especially when another external event (T1) offers not only a more natural basis of timing, but also a more reliable one? More critically, however, the predictability account cannot explain the shifts in R1 we found in Experiment 2. Anything leading up to R1 (the go signal and T1 itself) was just as predictable in the baseline as in the causal-control condition.
Interestingly, Stetson, Cui, Montague, and Eagleman (2006) demonstrated that violations of predictability can also lead to subjective reversals of temporal order. The participants in that study learned that an action typically produced a stimulus after 135 ms, but that sometimes that same stimulus might occur at other times, irrespective of the action, and might even occur before the action was taken. Under this manipulation, stimuli that occurred objectively up to 44 ms after the action were judged to have occurred before it. Although this finding is interesting from the perspective of temporal adaptation (Kennedy, Buehner, & Rushton, 2009), it is doubtful that the participants would have claimed causal control over those events that they thought had occurred before their actions.
In conclusion, it appears that the shifts in event perception and action planning we observed were genuinely induced by causality and cannot be reduced to variations in attentional demands or predictability. Causal binding—changes in event perception and action planning induced by mental constructs of causality—fits well within the broader framework of automaticity and priming (Bargh, 2006). More specifically, it suggests that the connection between mental constructs, on the one hand, and perception and action, on the other hand, is a two-way street (cf. Eagleman & Holcombe, 2002). People's mental representations, beliefs, and stereotypes arise as a function of perception, as outlined by Hume (1739/1888). Sometimes people even explain their conduct retrospectively, ascribing conscious will or causal agency to actions and outcomes over which in reality they had no control, if external cues or their own behavior is consistent with a causal explanation (Wegner, 2003). At the same time, however, low-level perception and motor action can be shifted by the presence of higher-level mental constructs, such as causality or trait constructs, even when people are unaware of them.
Footnotes
Acknowledgements
This research was funded by Engineering and Physical Sciences Research Council Grant EP/C004469/1, awarded to M.J.B. We thank Patrick Haggard, Kai Engbert, and Jon Kennedy for helpful discussion.