
Abstract
Migraine is a common, disabling, neurological problem whose acute management would benefit from the development of purely neurally acting therapies. The trigeminocervical complex is pivotal in nociceptive signaling in migraine, and is an accepted target for putative antimigraine agents. Whole-cell patch-clamp or extracellular recordings were made of trigeminal neurons identified in rat brainstem slices. Bath application of the large conductance calcium-activated potassium (BKCa) channel opener NS1619 caused a dramatic decrease of cell firing that could be reversed by the co-application of iberiotoxin. NS1619 hyperpolarized the resting membrane potential and reduced the frequency of spontaneous action potentials in these neurons. These data suggest the presence of BKCa channels in the trigeminocervical complex. In vivo in cat L-glutamate-evoked firing was facilitated in nociceptive neurons, also responding to stimulation of the superior sagittal sinus, in the trigeminal nucleus caudalis by the BKCa peptide antagonists, iberiotoxin and slotoxin. Of units tested, 70% responded to microiontophoretic application of the blockers, identifying a subpopulation of trigeminal neurons expressing toxin-sensitive BKCa channels. NS1619 inhibited 74% of cells tested, and this was reversed by slotoxin, suggesting that the action of NS1619 in these cells was mediated through BKCa channels. These data are consistent with the presence of BKCa channels in the trigeminal nucleus caudalis that are potential targets for the development of antimigraine treatments, and may also offer insights into receptor mechanisms involved in sensitization and thus allodynia, in migraine.
Introduction
Migraine is a common (1) and disabling (2) neurological disorder involving activation, or the perception of activation, of trigeminal neurons (3). It has been estimated to be the most costly neurological disorder in the European Community at more than €27 billion per year (4) and to cost the US economy $19.6 billion per year (5). Large conductance calcium-activated potassium channels, the BKCa or previously MaxiK channels, are ubiquitously expressed intrinsic membrane proteins that regulate many physiological functions (6). It has been postulated that BKCa channels constantly monitor the electrical and metabolic state of the cell based on their unique gating mechanism, being activated by a combination of membrane potential and intracellular calcium. BKCa channels are important in the nervous system (7, 8), where they act as regulators of neuronal excitability and neurotransmitter release (9, 10). Given the general concept of migraine pathophysiology including dyshabituation of normal sensory traffic (11), could BKCa channels be a target for antimigraine therapeutic development that may allow a suitable reduction in dural trigeminovascular traffic and thus be useful in migraine? Moreover, given the still substantial unmet need for new therapies in migraine of a non-vasconstrictor type, we have studied BKCa channels since they have both neural actions and no vasoconstrictor liability.
BKCa channels are very sensitive to the selective inhibition of the ‘pore-blocking’ peptides iberiotoxin (12) and slotoxin (13). Conversely, NS1619, a benzimidazolone analog, is an organic small molecule modulator of neuronal BKCa channels, and acts as a channel opener or activator (14); other novel substituted benzanilides have also been discovered (15). Compounds of this class have a potentially powerful influence in the modulation and control of numerous consequences of muscular and neuronal hyperexcitability. NS1619 activates BKCa channels in rat cortex (16, 17), and the effects of iberiotoxin and NS1619 on action potential firing of small and medium-sized sensory neurons from L6 and S1 dorsal root ganglia of adult rats have been examined in vitro (18). Iberiotoxin increases the firing frequency of the neurons in whole-cell patch-clamp studies, whereas NS1619 reversibly suppresses the action potential firing. These studies suggest that BKCa modulation can alter the firing properties of neurons in central and peripheral nociceptive pathways. Given the receptor-based targets of current therapies, such as serotonin 5-HT1B/1D receptor agonists, triptans (19), or calcitonin gene-related peptide receptor antagonists (20), a channel target would be a radically different approach to migraine therapy.
We made whole-cell patch-clamp and extracellular recordings from trigeminal nucleus caudalis (TNC) neurons identified in rat brainstem slices, with bath perfusion of NS1619 and iberiotoxin to examine the influence of BKCa channels on membrane potential and spontaneous action potential (sAP) firing in TNC neurons. The model of superior sagittal sinus (SSS) stimulation was also employed as it has proved useful in predicting agents that are effective in the acute treatment of migraine, such as the triptans (19), and has predicted lack of efficacy in other putative antimigraine agents, such as substance P/neurokinin 1 receptor antagonists (21). We explored whether modulation of BKCa activity in the cat trigeminocervical complex would effect nociceptive signaling. The effects of the selective BKCa peptide inhibitors iberiotoxin and slotoxin, as well as the activator NS1619, were tested using a combination of microiontophoresis and in vivo electrophysiology in the whole system cat SSS stimulation model.
Methods
Cat studies were conducted in accordance with a project licence issued by the Home Office of the UK under the Animals (Scientific Procedures) Act 1986 and all protocols were approved by the Institutional Animal Care and Use Committee of Amgen Inc. in accordance with the National Institutes of Health's Guide for the Care and Use of Laboratory Animals.
In vitro electrophysiology
Brain slice preparation
Transverse TNC slices were prepared from 10 Sprague-Dawley rats (2–4 weeks old; Hollister, CA, USA). The rats (35–110 g) were deeply anesthetized with an intraperitoneal injection of a pentobarbital sodium (60 mg/kg) and euthanized by decapitation, the brain and brainstem were rapidly removed and transferred to an ice-cold dissection solution containing 140 m
In vitro recording of trigeminal nucleus caudalis neurons in brain slices
TNC neurons were visualized by infrared differential interference contrast video microscopy (Axioskop 2; Zeiss, Oberkochen, Germany) using a C2741-60 Enhanced Charge-Coupled Device Camera C. Whole-cell patch-clamp and extracellular electrophysiological recordings were made using a Multiclamp 700B amplifier (Axon Instruments, Union City, CA, USA). Data were acquired using pClamp/Digidata 13200 (Axon Instruments). Patch pipettes had resistances of 3–5 MΩ for whole-cell recordings and 1–3 MΩ for extracellular recordings.
Whole-cell recordings of spontaneous excitatory post-synaptic currents (sEPSCs) were recorded from post-synaptic neurons in voltage-clamp mode. The external solution was ACSF. The holding potential was −57 mV with an internal solution containing 140 m
Application of drugs
All drugs were applied at known concentrations via a motorized perfusion system (403U/VM3; Watson, Wilmington, MA, USA) at a speed 2 ml/min. Stock solutions of the BKCa channel blocker iberiotoxin and slotoxin (Alomone Labs, Jerusalem, Israel), the AMPA/kainate antagonist 6-cyano-7-nitroquinoxaline-2,3-dione disodium (CNQX; Tocris Bioscience, Ellisville, MO, USA) and N-methyl-
Data analysis
Data were acquired with pClamp-9.2 and analyzed with Clampfit 9.2 (Axon Instruments) for spontaneous action potentials and Minis Analysis program (Synaptosoft, Decatur, GA, USA) for sEPSCs. The sEPSC events were counted if their amplitude was above the noise (5 pA) or significantly visible. These data are presented as mean ±
In vivo electrophysiology
Surgical preparation
Cats of either sex (n = 15; six female, nine male) weighing 3.2 ± 0.4 kg (mean ±
Arterial blood gases and other biochemical parameters, including pH, calcium and lactate concentrations, were monitored using an automated analyzer (Instrumentation Laboratory, Lexington, MA, USA). Core temperature was monitored and maintained between 37 and 39°C using a rectal thermistor probe and a low-electromagnetic-noise-emitting homeothermic heater blanket system (Harvard Apparatus, Holliston, MA, USA). Cats were ventilated with a 40% oxygen in air mixture (Model 6025 ventilator; UgoBasile, Comerio, Italy), and end-tidal CO2 was continuously monitored and maintained between 2.5 and 4.0% (Capstar-100 carbon dioxide analyzer; CWE) by adjustment of respiratory rate and depth to maintain arterial blood pH within physiological limits (pH 7.35–7.45). Heart rate was monitored by electrocardiography (CT-1000; CWE), and also derived from blood pressure changes. Depth of anesthesia was monitored periodically throughout the experiment by testing for sympathetic (pupillary and cardiovascular) responses to noxious stimulation and withdrawal reflexes in the absence of neuromuscular blockade. Supplementary doses of α-chloralose in 2-hydroxypropyl-β-cyclodextrin (Sigma) were given intravenously as required, at a rate of 5–10 mg kg−1 h−1 (24).
CNS surgery
A midline craniotomy (about 20 mm diameter) and C1–C2 laminectomy were performed in each cat, allowing access to the SSS and the area for recording neuronal activity in the TNC. To isolate the SSS, the adjacent dura mater and falx cerebri were dissected over a distance of about 15 mm. A small polyethylene sheet was inserted under the isolated sinus, laid over the outlying dura mater and tucked under the edges of the craniotomy. To prevent dehydration and to provide additional electrical insulation to the cortex, a circular polypropylene dam was sealed to the bone around the craniotomy with polymethylmethacrylate-based dental cement (Vertex, Zeist, the Netherlands) and filled with liquid paraffin (BDH Laboratory Supplies, Poole, UK). The likelihood of possible artefacts from arterial pulsation and respiratory movement (such as movement of the electrode from recording sites) was reduced by: bilateral pneumothoraces, kept patent with polypropylene tubes; immobilization of the spine by clamping a thoracic spinous process (1780 spinal unit; Kopf); clamping the C1 transverse processes, and clamping the remaining caudal portion of the dorsal C2 spinous process.
In vivo stimulation and recording
The isolated SSS was gently lifted onto a bipolar platinum hook electrode pair connected to a stimulus isolation unit (SIU5A; Grass Instruments, West Warwick, RI, USA). To activate primary trigeminal afferents and provide the initial search stimulus for responsive cells in the TNC, the SSS was supramaximally stimulated with stimulus-isolated (Grass SIU) square wave pulses from a Grass S88 stimulator (80–150 V, ∼50 µA; 250 µs, 0.3–0.5 Hz) after neuromuscular blockade with gallamine triethiodide (Concord Pharmaceuticals, Dunmow, UK; initially 5–10 mg/kg intravenously and maintained with 5–10 mg kg−1 h−1). The dura mater above the recording regions on the surface of the spinal cord was reflected after a midline incision and held to the bony edges of the laminectomy with N-butyl-cyanoacrylate, further stabilizing movement of the cord in this sling-like arrangement. Extracellular recordings and microiontophoretic delivery of test compounds were made using custom-built carbon-fiber containing multibarrelled-microiontophoresis combination electrodes (Carbostar 7S; Kation Scientific, Minneapolis, MN, USA). Recording electrode impedances were typically 1–3 MΩ when measured at 1 kHz in 0.9% saline (Impedance check module; FHC, Bowdoinham, ME, USA). After careful local removal of the pia mater, which was dissected away from a small area of the underlying spinal cord at the sulcus above the dorsal root entry zone, the electrodes were lowered into the cord substance around the C2 roots in the area of the dorsal root entry zone. The point of contact of the electrode tip with the pial surface was taken as the zero reference point. The electrodes were advanced or retracted in the cord substance in discrete 5-µm steps using an ultra-low-drift (<1 µm/h) LSS-100 microelectrode positioner system consisting of a piezoelectric motor (IW-711; Lateral Stability Option ± 0.2 µm lateral motion; Burleigh Instruments, Victor, NY, USA) and ultra-low-noise controller (6000ULN) attached to a heavy-duty micromanipulator (Kopf 1760–61). Tissue-culture grade agar (Sigma) 3% w/v in pyrogen-free saline (Baxter Healthcare) was set over the exposed cord after electrode insertion to further reduce cardiovascular-related movement of the spinal cord. Signal from the recording electrode attached to a high-impedance headstage preamplifier (NL100AK; Neurolog, Digitimer, Welwyn Garden City, UK) was fed via an AC preamplifier (Neurolog NL104, gain ×1000) through filters (Neurolog NL125; bandwidth about 300 Hz to 20 kHz) and a 50-Hz noise eliminator (Humbug; Quest Scientific, North Vancouver, BC, Canada) to a second-stage amplifier (Neurolog NL106) providing variable gain (to ×100). This signal (total gain about ×20 000 to ×95 000) was fed to a gated amplitude discriminator (Neurolog NL201) and analog-to-digital converter (Cambridge Electronic Design, Cambridge, UK), and to a microprocessor-based personal computer (DELL Latitude C800-PP01X; Bracknell, UK) where the signal was processed and stored. Filtered and amplified electrical signals from action potentials were fed to a loudspeaker via a power amplifier (Neurolog NL120) for audio monitoring, and were displayed on analog- and digital-storage oscilloscopes (Goldstar, LG Precision, Seoul, Korea and Metrix Electronics, Chauvin Arnoux, Paris, France, respectively) to assist the isolation of single unit activity from adjacent cell activity and noise.
To record the response of single units to SSS stimulation, post-stimulus time histograms were constructed on-line with Spike2 software (Cambridge Electronic Design) using 0.5-ms bins and saved to disk. During the experiments electrophysiological data, blood pressure, heart rate, core temperature and end-tidal CO2 were processed and recorded on VHS magnetic tape (Pulse Code Modulator; Vetter, Rebersburgh, PA, USA) and magnetic hard disk for documentation and later review.
Animals from which data are reported had cardiorespiratory parameters that were normal for α-chloralose anesthetized cats. Arterial blood gas and biochemical parameters were measured at intervals throughout experiments and were within normal limits (mean ±
Identification of receptive fields
Cells responding to SSS stimulation were characterized as receiving low threshold mechanoreceptor input if they responded to innocuous input, such as brushing or stroking of cutaneous receptive fields on the face or forepaws with a cotton pledget on a wooden shaft. They were characterized as nociceptive specific if they responded to noxious mechanical stimuli, such as pinching with toothed forceps or pricking with a needle, or wide dynamic range (WDR) if they responded to both [see Fig. 4B, (25)]. These cells usually had an increased firing rate in response to noxious stimuli.

Localization and classification of neurons recorded in vivo. (A) A transverse section through the spinal cord at the level of C2 is represented. Although the positions of the neurons are mapped to only one side of the cord in the figure, they represent results obtained from both the left-hand and right-hand side of the spinal cord. The scale bars represent a distance of 1 mm in both directions. Solid circles indicate sites where recording sites were marked with Pontamine Sky Blue (PSB; C.I. 24410) by microiontophoresis (−1.00 µA for 20–25 min) and identified histologically. Open circles indicate the positions of unmarked recording sites, or sites where marks could not be recovered, which were identified by reference to the position of dye marks at other recording sites or at the end of electrode tracks and electrode tip coordinates. (B) Representative rate histogram showing the responses evoked by mechanical stimulation of cutaneous VI receptive field on the head of the cat by innocuous stimuli (brush with a cotton pledget) and ‘noxious’ stimulus (pinch with toothed forceps). This representative cell was characterized as receiving wide dynamic range (WDR) input. (C) Representative bipolar compound action potential evoked by
Microiontophoresis
Micropipettes used for microiontophoresis had orifices in the range 1–3 µm and were filled with 200 m
After filling, electrode barrels passing useful (> 10 nA) iontophoretic currents at ±135 V compliance had resistances of 24–200 MΩin situ and impedances of 5.6–26.2 MΩ tested at 10 nA peak-to-peak at 1 kHz in saline. In particular, respective resistances and impedances for barrels containing 200 m
Identification of in vivo recording sites
The position of the recording electrodes was controlled by use of a heavy-duty stereotaxic micropositioner (Kopf 1760–61) with reference to the mid-point of the C2 dorsal roots. Together with the depth of the recording electrode tip with respect to the surface of the spinal cord at the dorsal root entry zone, as determined by the distance traveled display on the ULN6000 piezoelectric motor controller (Burleigh Instruments), this provided the coordinates of the recording sites. The location of selected recording sites and the end of electrode tracks were marked with microiontophoretically delivered Pontamine Sky Blue dye using a −1.00-µA current for 20–25 min. After euthanasia of the animals at the end of experiments with sodium pentobarbital (400 mg), followed by KCl (10% w/v; 5 ml), the section of spinal cord containing the recording sites was resected, fixed with neutral buffered 10% formalin, stored in phosphate buffered 30% sucrose, pH 7.4, until sunk and sectioned (40 µm; HM500 OM cryostat microtome; Microm Laborgeräte, Walldorf, Germany). Pontamine Sky Blue marks were counterstained with neutral red Nissl stain or nuclear fast red, procedures that allowed identification of the laminae of the gray matter. The positions of the recording sites within the cord were determined from histologically identified dye marks, and unmarked recording sites were located by reference to other dye marks, for example marking the end of recording tracks, and the stereotaxic coordinates of the electrode tip at the recording site.
Statistical analyses
Neuronal firing was distinguished from noise using an amplitude discriminator. Statistical evaluations were made for each unit using the mean rate of firing (Hz) evoked during each epoch of microiontophoretic
Microiontophoretic currents with a magnitude of at least 80 nA, and up to 400 nA for toxins, where ‘blocking’ did not occur at lower currents, were applied for at least 10 epochs of
Results
BKCa channels modulate the firing properties of trigeminal nucleus caudalis neurons post-synaptically
Whole-cell recordings in current-clamp mode were performed on TNC neurons in freshly sectioned slices of native rat brainstem. Individual neurons were visible under infrared differential interference contrast video microscopy. Spontaneous action potentials were verified by their firing properties, i.e. shape and size (Fig. 1A), and the effect of the sodium channel blocker tetrodotoxin (1 µ

Effects of iberiotoxin and NS1619 on the excitability of individual neurons from the trigeminal nucleus caudalis (TNC) in a slice preparation. (A) Expanded time-base view of spontaneous action potentials recorded from a single TNC neuron using the whole cell patch-clamp technique in current-clamp mode. (B,C) Representative traces illustrating the effect of 100 n
The impact of BKCa channels on the membrane potential and spontaneous action potentials was first examined by bath perfusion of the selective antagonist iberiotoxin. Bath application of iberiotoxin caused a significant depolarization (> 2 mV) in 10 of 18 neurons (56%). In seven out of 13 neurons (54%), iberiotoxin significantly increased the firing rate of spontaneous action potentials (Fig. 1B). Typically it took 1–3 min for a robust increase in firing frequency to occur, but in a couple of neurons the effect was very fast and occurred in as little as 15 s after drug perfusion was initiated. We believe the differences in time to reach peak effect were due to how deep the neurons were in the slice preparation. Interestingly, two of the seven neurons that showed positive modulation in firing in the presence of iberiotoxin exhibited decreased firing with long-term exposure (Supplemental Fig. S1). This effect could be analogous to the tachyphylaxis effect observed in the in vivo recordings (Supplemental Fig. S2). As seen in Fig. S1A, these neurons exhibited strong depolarization in the resting membrane potential in response to iberiotoxin. Presumably, the reduction in firing was due to sodium channel inactivation.
We then tested whether activating BKCa channels would have the opposite effect to iberiotoxin on the membrane potential and spontaneous action potentials. Indeed, 30 µ
Combined perfusion of CNQX and CGS19755 did not alter the firing frequency or amplitude of the spontaneous action potentials (Supplemental Fig. S3A,B).
BKCa channels also contribute to presynaptic glutamatergic neurotransmitter release
To evaluate the possibility that BKCa channels localized on presynaptic terminals may modulate transmitter release, sEPSCs were recorded before and after perfusion of iberiotoxin (Fig. 2). To prevent the influence of post-synaptic BKCa channels, the potassium channel blocker cesium (149 m
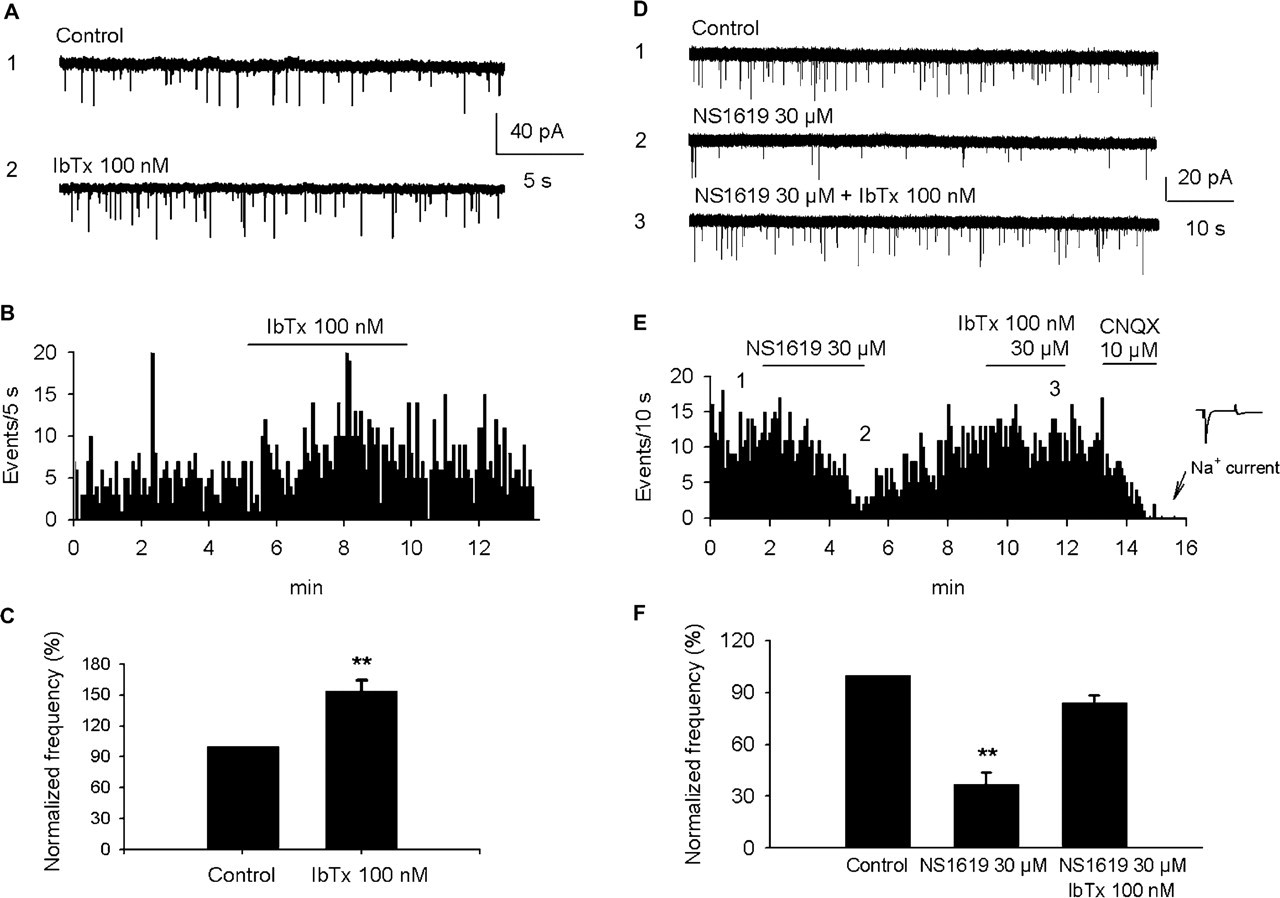
Effects of iberiotoxin and NS1619 on the frequency of spontaneous excitatory postsynaptic currents (sEPSCs) in individual neurons from the trigeminal nucleus caudalis in a slice preparation. (A) Traces illustrating sEPSCs and the effect of 100 n
Effects of NS1619 were also tested on the frequency of sEPSCs in TNC neurons (Fig. 2D–F). The raw traces and corresponding frequency histogram illustrate that 30 µ
BKCa channel blockers facilitated TNC firing in vivo
Extracellular recordings were made from 72 neurons in the trigeminocervical complex of cats (32). Neurons responding to the SSS search stimulus that were also activated by

Effects of iberiotoxin and slotoxin on
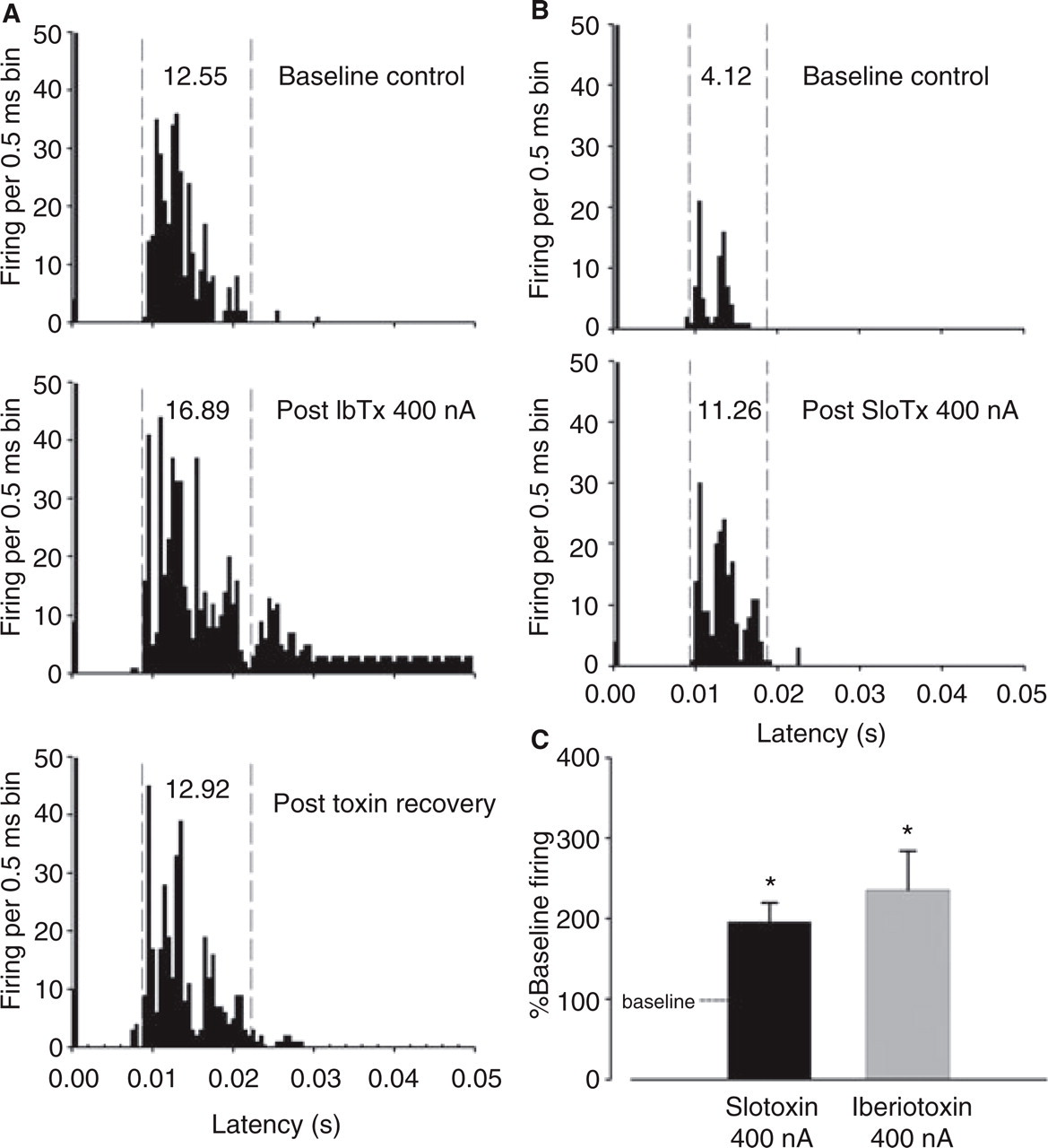
Effects of iberiotoxin and slotoxin on electrically evoked superior sagittal sinus (SSS)-linked trigeminal nucleus caudalis (TNC) neuron firing in vivo. (A,B) Representative post-stimulus histograms illustrating that direct electrical stimulation of the SSS recruits units (baseline controls) facilitated by iberiotoxin (IbTx) (A; 400 nA for 5 min) and slotoxin (SloTx) (B; 400 nA for 5 min). The histograms represent the cumulative frequency of action potential firing of cells in the TNC in response to 50 successive electrical stimuli of the SSS at latencies up to 50 ms divided into 0.05-ms bins. The stimulus artefact can be seen on the most left-hand side of each histogram. Supramaximal electrical stimuli were applied to the SSS via bipolar platinum hook electrodes carefully placed under the sinus. The dashed vertical lines represent cursor positions used for calculation of percentage change between various conditions and the bold figures indicate mean firing per bin between the cursors. (C) Mean effects of iberiotoxin and slotoxin on electrically stimulated SSS-evoked firing of cells showing significant facilitation by both BKCa channel blockers [P < 0.05, two-tailed one-sample t test vs. baseline (100%, dashed line), n = 5 and 5]. There was no significant difference in facilitation between the two toxins (t 4 = −0.88, P > 0.43, paired-sample two-tailed t test).
Sixty-five cells were reversibly excited by microiontophoretic application of
For neurons that had identified cutaneous receptive fields, which were always ipsilateral and mostly on the head above the ear or on the forepaw, iberiotoxin and slotoxin facilitated
In cats, microiontophoresis of iberiotoxin and slotoxin significantly facilitated
Interestingly, tachyphylaxis of the facilitation from iberiotoxin and slotoxin occurred in about one-third of the responding neurons [slotoxin six of 19 (32%); iberiotoxin 11 of 29 (38%)]. Typically the tachyphylaxis occurred at current ejections > 100 nA, at which the initial increase in firing rate was often large. Cells were considered to respond ‘tachyphylactically’ if the Pearson correlation across the first five epochs of
Facilitation of the
BKCa channel blockers facilitated electrically evoked SSS TNC firing in vivo
We then sought to determine if the toxins could facilitate neuronal firing in the TNC that was evoked by an electrical stimulus. In those neurons whose
NS1619 decreased l -glutamate-evoked firing in SSS-linked TNC cells in vivo
The effects of the BKCa selective toxins suggested that BKCa is expressed in the nociceptive pathway of the trigeminocervical complex, and that BKCa inhibition increases the firing frequency of most neurons that receive nociceptive input. Therefore, we hypothesized that BKCa agonism would reduce firing in a toxin-sensitive manner. Indeed, NS1619 significantly inhibited

NS1619 inhibits
Discussion
These data provide direct evidence that large conductance calcium-activated potassium (BKCa) channels modulate the firing properties of neurons with dural trigeminovascular nociceptive input. Three independent lines of evidence support this interpretation. First, the BKCa selective peptide inhibitors, slotoxin and iberiotoxin, dramatically facilitated the firing evoked by microiontophoretic application of
BKCa modulation of nociceptors
The data demonstrate that BKCa channel modulators affect neuronal activity in response to excitatory chemical and electrical stimuli within the trigeminovascular nociceptive pathway. However, we are unable to determine in the in vivo preparation whether the modulation occurs presynaptically, on the invading axon from the primary sensory afferent emanating from the trigeminal ganglion, or post-synaptically, on the second-order neuron within the TNC itself, or at both sites. Since combined perfusion of CNQX and CGS19755 did not alter the firing frequency or amplitude of the spontaneous action potentials in the brain slice preparation, this suggests that the spontaneous action potentials from TNC neurons in that preparation are generated from intrinsic activity of the post-synaptic neurons. In patch clamping experiments, cesium was used to block post-synaptic BKCa channels to evaluate the possibility that BKCa channels localized on presynaptic TNC terminals may modulate neurotransmitter release. NS1619 reduced the frequency of sEPSCs and iberiotoxin reversed the effect of NS1619 under these conditions, indicating that BKCa channels also contribute to presynaptic neurotransmitter release. The complete block of sEPSCs by CNQX at the conclusion of these experiments indicates that they were mediated by glutamatergic AMPA/kainate receptors after presynaptic glutamate release under the influence of BKCa channels.
Strong evidence supports the expression and physiological role of toxin-sensitive BKCa channels on nociceptors. Small and medium-sized neurons (putative nociceptors) from primary cultures and thin slice preparations of rat dorsal root ganglion express toxin-sensitive BKCa channels (8, 18). In these in vitro settings, voltage-clamp experiments of single BKCa channels, as well as current-clamp recordings of whole neurons, indicate that the agonists NS1619, ethanol, and trichloroethanol, as well as the pore-blocking peptide iberiotoxin, have profound effects on the single channel kinetics, the action potential shape, as well as intrinsic firing properties. Similarly, NS1619 reduced single unit firing of Aδ and C-fibers recorded from guinea pig trachea and blunted the citric acid aerosol cough response in an iberiotoxin-sensitive manner. Several of these studies reported that a majority of the neurons tested (∼55–70%) express iberiotoxin-sensitive BKCa channels. This is in direct agreement with our results, where ∼70% of the TNC neurons electrically linked to the SSS (Aδ fiber input) were toxin sensitive and more than half of the TNC neurons tested in slice preparations significantly depolarized or increased their rate of firing in response to bath application of iberiotoxin. Little has been reported about BKCa channel expression or modulation on second-order neurons within the dorsal horn of the spinal cord or in the TNC. However, given the ubiquitous nature of BKCa channel expression within the central nervous system, it would be surprising if BKCa channels were not also expressed in these neurons.
BKCa channels have different functional roles depending on their subcellular localization. On the cell body, for example, BKCa channels reduce repetitive firing by shortening the action potential duration, speeding repolarization, and contributing to the fast after-hyperpolarization. Alternatively, at the presynaptic terminal, BKCa channels contribute to neurotransmission by counteracting calcium influx, thereby limiting vesicle release. Therefore, it is entirely possible that different mechanisms underlie the BKCa modulatory effects found in the
Potential impact of regulatory β-subunits
BKCa channels can be formed by homotetramers of α-subunits or by complexes with at least four families of β-subunits (β1–4). The coexpression of β-subunits, which have two transmembrane domains and a large extracellular loop, modifies the BKCaα-subunit kinetic properties, intracellular calcium sensitivity, intrinsic modulation, and pharmacology such as toxin sensitivity. Slotoxin and iberiotoxin specifically and reversibly inhibit the BKCa channel pore-forming α-subunit as ‘pore-blockers’(35). However, slotoxin is capable of differentiating three types of BKCa channel complexes, α, α + β1, and α + β4 (13). Slotoxin inhibits the homomeric α-subunit with very low potency (IC50∼1–10 n
Clinical relevance
To explore a possible therapeutic potential for BKCa modulators we employed the model of SSS stimulation. Stimulation of dural structures is painful in humans (36), and in animal models, stimulation of dural sites, including the SSS and middle meningeal artery, results in neuronal activation in the trigeminovascular system. This activation is significantly inhibited by acute antimigraine drugs (37–45). Moreover, compounds found to be ineffective in acute migraine, such as substance P/neurokinin 1 receptor antagonists (21) and plasma protein extravasation inhibitors (46), are also ineffective in blocking trigeminocervical transmission in this model [e.g. (47, 48)]. This model compares favorably with investigation of the aura homologue phenomenon cortical spreading depression [CSD, (49)], which has not been predictive at all of the likely utility of acute antimigraine treatments (50, 51). The CSD model may be more useful in predicting effectiveness of preventive medicines (52, 53). Inhibition of trigeminal nerve activation is therefore helpful in evaluating new targets for the treatment of acute migraine. Another interesting aspect of the BKCa channel in migraine may be sensitization, the clinical manifestation of which is likely to be allodynia (54, 55). Our data indicate that should the BKCa channel be dysfunctional, as modeled by channel blocker treatments, trigeminal neurons become sensitized. Since not all migraineurs have allodynia, might it be that allodynia represents an additional functional burden in that group mediated by cosegregation of migraine biology and BKCa, or a similar channel, dysfunction? Finally, the widespread distribution of these channels on smooth muscle in vessels and in the lung may provide both challenges and opportunities. As dilators they might be predicted to reduce blood pressure or even have therapeutic actions in airways disease, although one might observe that simple is best in therapeutics and widespread actions may produce unwanted side-effect issues at some point (56).
Concluding remarks
BKCa channels provide a negative feedback mechanism for cells to pass strong hyperpolarizing potassium currents during times of elevated firing frequencies that cause prolonged depolarizations and elevated intracellular calcium levels. Substances such as slotoxin and iberiotoxin, which block these channels, remove the BKCa-dependent negative feedback loop to membrane polarization and cause neuronal hyperexcitability. Conversely, BKCa activators, such as NS1619, cause membrane hyperpolarization and reduce neuronal excitability and therefore have a potentially powerful influence in blunting the repetitive firing seen in many disease states. One such example of an important disease state where such mechanisms could be conceived would be migraine. Our results are consistent with the presence of large conductance, calcium-activated potassium channels on nociceptive trigeminovascular neurons. These channels may provide a completely novel target for drugs aimed at the treatment of primary neurovascular headaches, such as migraine and cluster headache.
Competing interests
D.C.I. and R.Y. are employees of Amgen.
Acknowledgements
The authors thank Paul Hammond and Michele Lasalandra for their excellent technical assistance. Dr Ken Hunter, Elizabeth Sharp, and Dr Michael Taylor of the Scottish Agricultural Science Agency for analysis of chloralose samples.