
Abstract
Background
Large-scale epidemiological studies support an important role for susceptibility genes in the pathogenesis of ischemic stroke, with phosphodiesterase 4D identified as the first gene predisposing to ischemic stroke. Several single nucleotide polymorphisms within the phosphodiesterase 4D gene have been implicated in the pathogenesis of stroke.
Aim
Undertake a multivariate analysis of six single nucleotide polymorphisms within the phosphodiesterase 4D gene in a previously defined Australian stroke cohort, to determine whether these single nucleotide polymorphisms have an association with ischemic stroke.
Methods
This case–control study was performed using an existing genetic database of 180 ischemic stroke patients and 301 community controls, evaluated previously for cerebrovascular risk factors (hypertension, hypercholesterolemia, diabetes, paroxysmal atrial fibrillation, smoking and history of stroke in a first-degree relative). Based on previously reported associations with large vessel disease, ischemic stroke, cardioembolic stroke or a mixture of these, six single nucleotide polymorphisms in the phosphodiesterase 4D gene were selected for study, these being single nucleotide polymorphisms 13, 19, rs152312, 45, 83 and 87, based on previously utilized DeCODE nomenclature. Single nucleotide polymorphisms were genotyped using a sequence-specific polymerase chain reaction method and gel electrophoresis. Logistic regression was undertaken to determine the relevance of each polymorphism to stroke. Further analysis was undertaken to determine the risk of stroke following stratification for stroke sub-type and etiology.
Results
Significant odds ratios were found to be associated with cardioembolic strokes in two single nucleotide polymorphisms: rs152312 and SNP 45 (P<0·05).
Conclusions
Our findings demonstrated an association between cardioembolic stroke and phosphodiesterase 4D single nucleotide polymorphisms rs152312 and 45. No significant association was found for the other four single nucleotide polymorphisms investigated within the phosphodiesterase 4D gene. We propose that the results from this Australian population support the concept that a large prospective international study is required to investigate the role of phosphodiesterase 4D in the cardiogenic cause of ischemic stroke.
Introduction
Large-scale epidemiological studies support an important role for susceptibility genes in the pathogenesis of ischemic stroke (1–3). A unique Icelandic familial stroke study identified Phosphodiesterase 4D (PDE4D) as the first mapping of a gene to predispose to ischemic stroke, independent of conventional risk factors (4, 5). Vascular endothelial cells act as gatekeepers to control the infiltration of inflammatory cells via adhesion molecules that play a central role in the recruitment of inflammatory cells (6). The PDE4D degrades 3′, 5′-cyclic AMP (cAMP), which is a key signalling molecule involved in the inflammatory responses of vascular cells (6, 7). Inhibitors of PDE4 have been reported to increase cAMP levels and adhesion in vascular endothelial cells (8, 9) and decrease migration of vascular smooth muscle cells (10), as well as inhibiting the expression of the cell-adhesion molecule E-selectin on human lung microvascular endothelial cells (11). Because of these effects, PDE4D has been postulated to contribute to vascular disease through its role in processes important in the pathogenesis of atherosclerosis, for example, through inflammation and plaque instability (5, 12).
However, the findings of many subsequent individual studies have not substantiated the original Icelandic association, with meta-analysis of PDE4D single nucleotide polymorphisms (SNPs) casting doubt on the conclusion of individual studies (13). In this meta-analysis, based on all PDE4D studies before 2008, Bevan and colleagues found that although there were several significant associations between PDE4D and ischemic stroke, all became nonsignificant once the original Gretarsdottir et al. study (5) was excluded. When they pooled studies on Caucasian populations, there were significant associations between several SNPs and ischemic stroke sub-types, particularly SNP 45 and combined large vessel disease and cardioembolic stroke sub-groups. Nonetheless, when the original study by Gretarsdottir and colleagues was excluded, the results again became nonsignificant. Bevan et al. (13) concluded that meta-analysis did not support studies replicating the original Gretarsdottir and colleagues study for any association of PDE4D with ischemic stroke. Because the data they presented could not totally exclude a link between PDE4D and ischemic stroke, they proposed that any association that may exist was likely to be weak and possibly restricted to specific populations: the studies analyzed were undertaken on populations in Iceland, USA, Greece, Japan, Sweden, Pakistan, Australia, the Netherlands, UK and Germany.
A recent review (14) also commented on the relevance of differing findings in different populations, noting that many questions with respect to the role of PDE4D in stroke development remain unresolved. Genetic stroke studies from North India (15) and South India (16) showed that different populations demonstrated different linkages between SNP 83 and ischemic stroke, and SNP 83 with large artery atherosclerosis (LAA) and small artery occlusion, respectively. Two separate recent medium-sized studies of Shanghai Chinese (17) and Beijing Chinese (18) found an association between SNP 83 and the combined cardiogenic and carotid stroke sub-groups and of SNP 83 with atherothrombotic stroke, respectively. The only Australian study assessing the potential link between genetic factors and stroke was performed in Western Australia (19). The findings in this study of a significant association between PDE4D and ischemic stroke have not been replicated. We initiated an independent South Australian study seeking an association between six PDE4D SNPs, ischemic stroke and stroke risk factors in a previously defined South Australian stroke cohort (20) and sought to determine the relevance of these polymorphisms to either cardioembolic stroke or other stroke sub-types. We chose the six SNPs based on positive associations in previous studies, an SNP frequency in the population >10% and underlying biological plausibility. Three of these SNPs (45, 83 and 87) were studied independently by the Western Australian researchers (19).
Methods
We genotyped DNA from 180 patients with acute ischemic stroke and 301 age- and gender-matched controls, sourced from our earlier prospective case–control study and using the same method (20), with research ethics approval. Patients for this study were admitted with acute ischemic stroke to one of five major hospitals within metropolitan Adelaide, South Australia, and were invited to participate in the study. All participants were evaluated for known cerebrovascular risk factors, including:
age
gender
race ethnicity
hypertension
hypercholesterolemia
diabetes mellitus
past history of stroke
smoking history
family history of stroke, and
atrial fibrillation (AF).
A neurologist (J. Jannes) made the diagnosis of ischemic stroke in accordance with the World Health Organization definition (21). All patients underwent brain computerized tomography (CT) in order to allow the exclusion of intracerebral and sub-arachnoid hemorrhage.
Stroke sub-type was determined using the Oxford Community Stroke Project criteria (22). Subjects were further classified into lacunar syndrome vs. nonlacunar syndrome (i.e. total anterior circular syndrome, partial anterior circulation syndrome and posterior circulation syndrome) and also cardioembolic vs. noncardioembolic causes, based on the presence of AF, determined by review of the 12-lead electrocardiograms (ECG) performed on admission for all patients. AF was also deemed present if there was a reported history of paroxysmal AF, regardless of the cardiac rhythm on ECG. Unless other investigations (e.g. high-grade internal carotid artery stenosis or stroke affecting multiple cerebrovascular territories in the absence of an identifiable cause) showed otherwise, all patients with AF were recorded as cardioembolic in origin. Patients with cardiac valve prostheses were also coded as cardioembolic in origin if there was no clinical or radiological evidence of an alternate thrombotic source. The full study demographics and risk factor characteristics have been described previously (20) and are summarized in Table 1.
Demographic characteristics and prevalence of risk factors for patients with ischemic stroke and controls
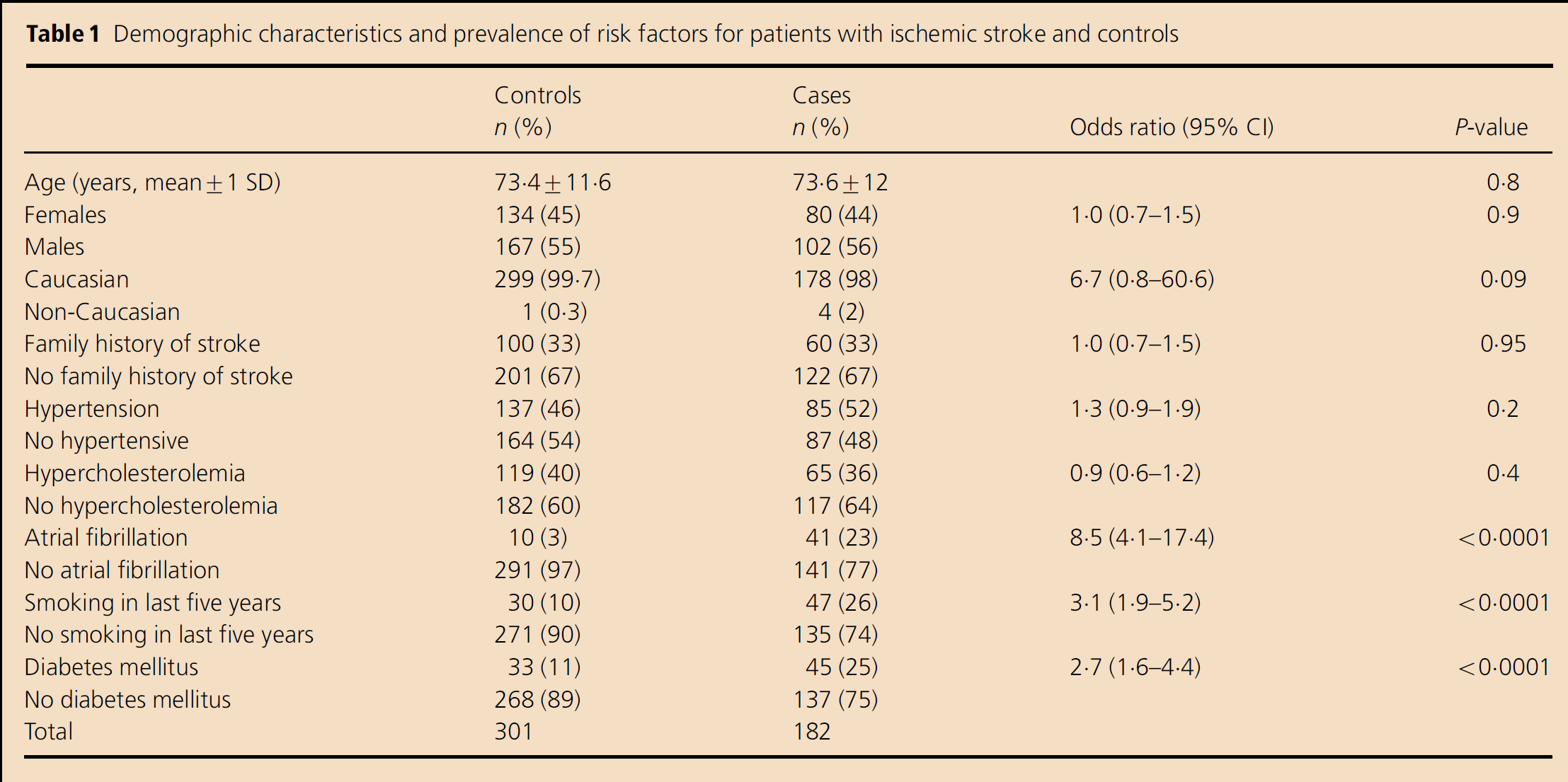
The control group consisted of nonhospitalized subjects from metropolitan Adelaide with no personal history of cerebrovascular disease. Controls, selected by random sampling of the South Australian electronic telephone directory, were group-matched with patients for age (within five-year strata) and gender.
In this study, 201 consecutive patients with acute ischemic stroke were asked to participate. One hundred and eighty-two (90·5%) agreed to participate, and 137 (75%) of these were first presentation ischemic stroke. No significant differences were observed between the two groups in terms of demographic variables (Table 1). Among the known risk factors examined, AF was found to be associated with the highest risk of ischemic stroke, with 23% of cases vs. 3% of controls affected (odds ratio (OR): 8·5; 95% confidence interval (CI): 4·1–17·4). A history of smoking within the past five-years (OR: 3·1; 95% CI: 1·9–5·2) and diabetes mellitus (OR: 2·7; 95% CI: 1·6–4·4) were also observed to be significantly associated with ischemic stroke. No association was found between ischemic stroke and a history of stroke in a first-degree relative, hypertension, hypercholesterolemia (Table 1) or for the use of antihypertensive, antiplatelet and lipid-lowering medication (results not shown). Two patients died after enrollment in the study and before venous blood sampling, so that genetic analysis could not be performed; hence, 180 patients remained for the full study.
Six SNPs were chosen based on positive associations in previous studies, an SNP frequency >10% and underlying biological plausibility. We retained the DeCODE nomenclature from the original Icelandic study mapping the PDE4D gene: SNPs 13, 19, rs152312, 45, 83 and 87 (5). These SNPs are located in a region near the alternative forms of exon 1 in the gene.
Allele-specific and consensus primers were designed using the sequences from the Entrez Human Genome public database. Specific sequences were synthesized by GeneWorks Pty Ltd, Adelaide, South Australia, and the resultant polymerase chain reaction (PCR) products were confirmed by sequencing (Department of Molecular Pathology, Institute of Medical and Veterinary Science, Adelaide, South Australia). Genotyping and quality control methods have been described previously (20, 23). An internal positive control was run for every PCR well, consisting of a pair of primers specific to an invariant section of the HLA DRB1 gene, to ensure that negative alleles were not due to failed polymerase reactions.
Power was estimated (with
All statistics were analyzed with the
Results
All SNPs were in accordance with expected Hardy–Weinberg equilibrium ratios. Data in Table 1 show that AF, smoking and diabetes were significant confounders affecting the risk of stroke in this population group, as reported previously (20).
The minor allele frequencies we found for each SNP in the control population for all patients with ischemic stroke and for distinctive sub-types are presented in Table 2. For all strokes and for the sub-types classified as lacunar stroke, nonlacunar stroke and noncardioembolic stroke, no significant associations with these six PDE4D SNPs were found. However, for the cardioembolic stroke sub-type, an association was found with two of the SNPs, rs152312 and SNP 45, as shown in Table 3. Significant confounding factors for cardioembolic stroke and rs152312 were AF, hypertension, male gender and diabetes, while those for SNP 45 were AF and smoking. However, the data for both of these SNPs remained significant, even after adjusting for these confounding factors.
Minor allele frequency of PDE4D SNPs
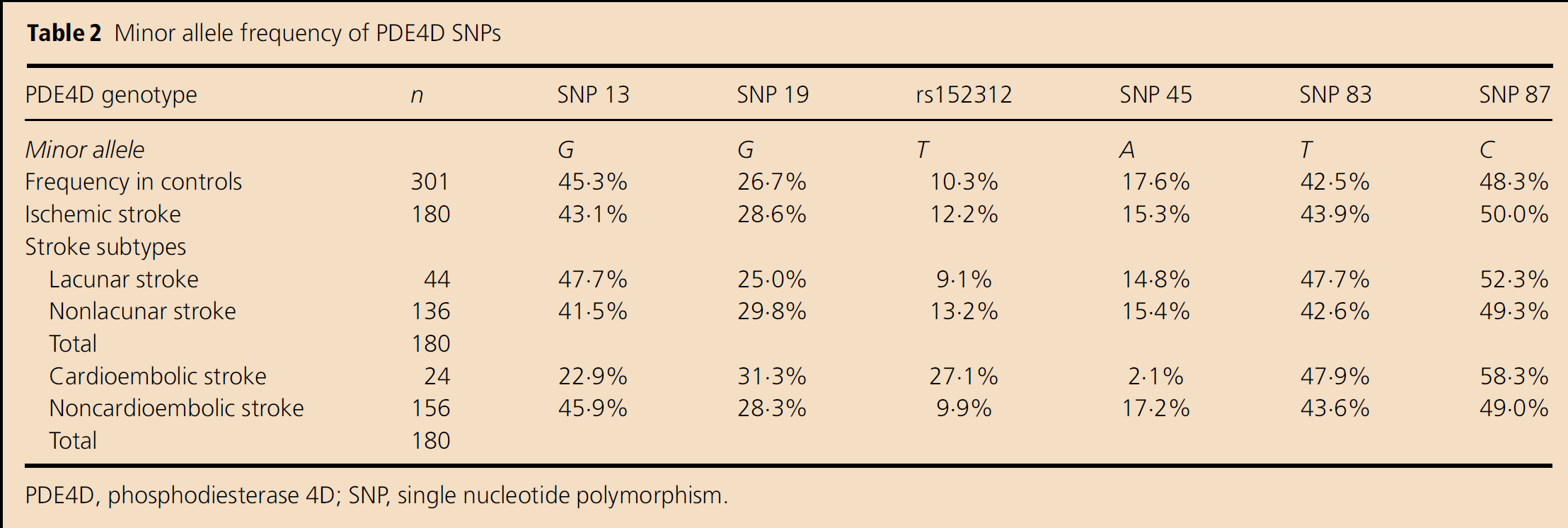
PDE4D, phosphodiesterase 4D; SNP, single nucleotide polymorphism.
Association of PDE4D rs152312 and SNP 45 alleles with cardioembolic stroke
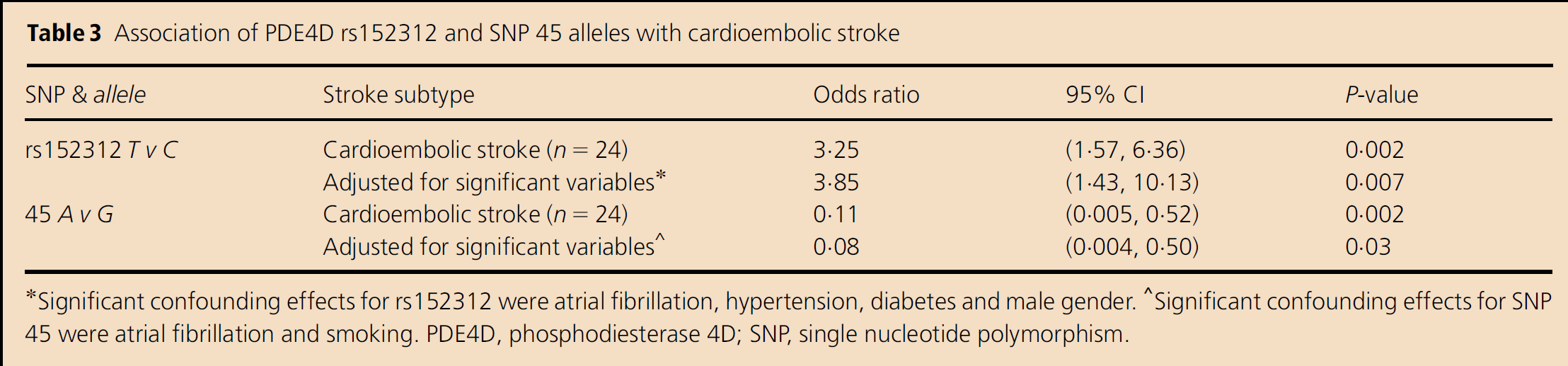
Significant confounding effects for rs152312 were atrial fibrillation, hypertension, diabetes and male gender.
Significant confounding effects for SNP 45 were atrial fibrillation and smoking. PDE4D, phosphodiesterase 4D; SNP, single nucleotide polymorphism.
Table 4 shows the genotypes for SNP rs152312 and SNP 45 and their association with the cardioembolic stroke sub-type (CE Stroke column) after important confounding factors were taken into account. This demonstrates that the SNP rs152312 exerts significant effects on cardioembolic stroke. The ORs for the SNP (TT) vs. the wild genotype (CC) (OR=16·37, CI 1·69–127·8, P=0·01) and the combined nonwild genotypes (CT+TT) vs. CC (OR=3·54, CI 1·05–12·33, P=0·04) for cardioembolic strokes are both significant. There are also significant effects with SNP 45 and cardioembolic stroke after the confounding effects are taken into account for the combined nonwild (GA+AA) vs. the wild (GG) genotype, although not significant for the single gene copy GA vs. the wild genotype (P=0·09). There is obviously no association with the noncardioembolic stroke sub-group (labelled Non-CE Stroke in Table 4), nor was there any association with the other four SNPs' genotypes found (data not shown).
Analysis of SNPs between controls and stroke subtypes, after adjusting for significant confounders
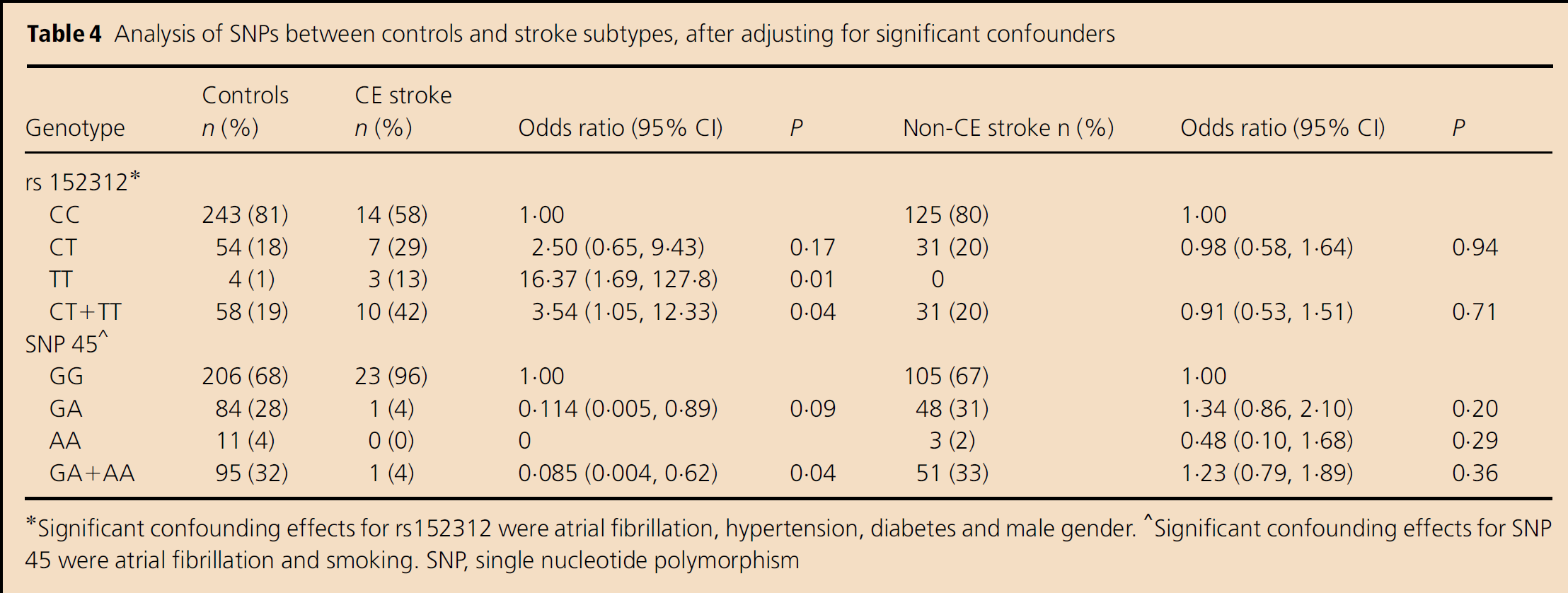
Significant confounding effects for rs152312 were atrial fibrillation, hypertension, diabetes and male gender.
Significant confounding effects for SNP 45 were atrial fibrillation and smoking. SNP, single nucleotide polymorphism
In the analysis of the other four SNPs, what appeared to be a third significant OR with the SNP 13 ‘G’ allele and cardioembolic stroke (OR=0·360; C.I. 0·17–0·70, P=0·002) became nonsignificant when adjusted for the confounding factors of AF, diabetes and hypertension (OR=0·45; C.I. 0·18–1·03, P=0·07) (data not shown).
Discussion
In our study, after allowing for important confounders such as AF, significant OR were found to be associated with cardioembolic strokes in SNPs rs152312 and 45 (P<0·05). We were powered sufficiently to show no association for any of the six SNPs with total ischemic stroke in our study. Although our sub-group analysis was not powered sufficiently to detect a significant association with sub-groups of ischemic stroke, we consider that our findings overall add to the body of evidence supporting the hypothesis that PDE4D may contribute to the risk of cardioembolic stroke.
The SNP 41 used in the Gretarsdottir et al. study is actually rs12153798 (26), which showed links with combined cardiogenic and carotid stroke (5). It was originally identified as rs152312, but this SNP, although studied by the Gretarsdottir group, was not reported in their supplementary tables as showing any association with stroke. Because of the potential for confusion, we decided to use the nomenclature rs152312 rather than SNP41 in this paper. Our SNP rs152312 data support the findings of Woo et al. (27), which reported an association between rs152312 and cardioembolic stroke in their Caucasian US population. However, in contrast to their reported association of SNP 87 with cardioembolic stroke, we found no association of SNP 87 with either cardioembolic stroke or total ischemic stroke.
With respect to SNP 45, we found that when compared with the major ‘G’ allele, the nonwild ‘A’ confers a protective effect (OR<1) against having a cardioembolic stroke, either before or after adjustment for confounding factors. A more recent Swedish study also found a protective association between SNP 45 and total ischemic stroke (28). In contrast to both these studies, in the original Icelandic study, an at-risk or positive association (greater risk of stroke) was reported between SNP 45 and the combined cardiogenic and carotid stroke sub-type patient groups (5). Our finding is also in contrast with a study from USA that found that the ‘A’ allele had a ‘nonsignificant at-risk effect’ in nonhypertensive ischemic stroke patients (29). A study in a Swedish population found a ‘nonsignificant link’ between the ‘G’ allele and stroke that increased in the LAA sub-type of stroke, and was a component of an SNP 45-SNP 41 ‘GA’ haplotype that showed a significant increased risk of stroke in both the LAA group and for the combined LAA and cardioembolic group (30). It remains to be seen whether the protective effect of the non-‘G’ allele is due to an allele exchange between the Icelandic population's ‘G’ allele and another nearby, undetected ‘at-risk’ allele.
Our study is the second study of PDE4D in an Australian cohort, with the first being in a population from the western state, while this study is from a southern state. In the Western Australian study, the authors reported a significant association between PDE4D and ischemic stroke (19). They also conducted a meta-analysis of nine case–control studies (including their own) that demonstrated a strong association with SNPs 41, 83 and 87. They noted that there was statistical heterogeneity in the direction of the association between individual SNPs and stroke that suggested that the SNPs tested were in linkage disequilibrium with the causal alleles (19). Some of the alleles that we examined were also investigated by their study: SNPs 45, 83 and 87. For these SNPs, we report the same minor alleles, which the Western Australian authors noted were the converse of the at-risk alleles reported by Gretarsdottir and colleagues. In contrast with the Western Australian study, which found an association of SNPs 83 and 87 with ischemic stroke (19), we found no association of these two SNPs with either total ischemic stroke or with sub-groups.
We assigned primers for rs152312 based on the original report detailing ‘SNP 41’ (5). In the Genebank sequence gb|:AC016607.6|AC016607 (Homo sapiens chromosome 5 clone CTD-2165C), rs152312 is 175 base pairs distant from SNP 41. Our chosen SNP rs152312, the correct SNP 41 (rs12153798) and SNP 45 are all contained within the promoter 1a isoform of the gene, being at relative positions in exon 1a of 144510, 144685 and 149009, respectively (31, 32). Because SNP 41 and SNP 45 (4324 base pairs apart) are highly linked (r2=0·94) (31), the linkage between rs152312 and SNP 41 (only 175 base pairs apart) would be expected by genetic theory to be even greater.
With regard to our finding of a lack of association between the SNP 13 ‘G’ allele and the cardioembolic stroke sub-type (P=0·07), it is of interest to note that a previous association has been reported in the United Kingdom between homozygous ‘G’ SNP 13 alleles and cardioembolic stroke after controlling for risk factors (33).
As reported previously, the conventional risk factors of AF, smoking and diabetes are highly significant confounders affecting the risk of stroke (P<0·001) in all stroke patients (20). In our present analysis of stroke sub-groups, hypertension and male gender were also found to be significant confounders in strokes of cardioembolic origin, although they had no significant impact in the noncardioembolic or the total ischemic stroke groups (noncardioembolic data not shown).
Populations of varied ethnic origins report different SNP frequencies for PDE4D in their control population. For example, SNP 45 is one of two PDE4D SNPs that showed linkage in the original Icelandic population (5), but was found to be monomorphic in an Asian population (34). Data from our control group for SNP 87 demonstrated that the ‘T’ major allele frequency is 51·8%, very close to the 51·7% found in a United Kingdom study (33), but at slight variance to the dataset from Western Australia that showed a control population frequency of 56·1% (19). Similarly, our control group had SNP 83 ‘C’ and SNP 45 ‘G’ frequencies of 57·7% and 82·3%, respectively; the Western Australian dataset had frequencies of 60·1% and 86·3% (19). These variances in frequency may represent different sampling methods or differences in the ethnic origins of the populations studied.
In relation to the cardiogenic causes of strokes, aortic arch atherosclerosis (AAA) is an important source of cerebral emboli, which may increase the risk for ischemic stroke (35). Atherosclerotic plaques that are at least 4 mm in thickness in the aortic arch have been shown to be significant predictors of stroke and other vascular events (36). Patients with severe AAA have over four times the risk of stroke or peripheral embolism and recurrent stroke is also more common in this same patient group (37). Indeed, the prevalence of severe AAA in stroke patients is over 20% (38). One study of 100 consecutive patients with embolic cerebral ischemia of the internal carotid or vertebrobasilar arteries found a cardiac source of emboli in 28% of their patients and AAA was found to be the probable source for embolic stroke in 14% of patients (35).
In our assessment of cardioembolic stroke, some patients may have had AAA. Cardioembolic stroke was defined as being due to an intracardiac source, as determined by the neurologist, for example, AF, recent myocardial infarction or thrombus on cardiac imaging. One plausible explanation is that there is coexistent AAA, but this was not actively investigated or assessed and requires further research. We are unaware of any publications linking AAA with genetic risk.
Recent meta-analysis studies of different ethnic populations provide conflicting results with respect to the importance of the role of PDE4D in the polygenic risk of ischemic stroke (13, 28, 31, 34). However, the PDE4D gene has been postulated to contribute to large vessel disease through its role in inflammation and plaque stability, with changes involving processes that are important in the pathogenesis of atherosclerosis (5, 12). As such, there is biological plausibility for the role of PDE4D in the pathogenesis of atherosclerosis and ischemic stroke, including strokes of cardiogenic origin. Further prospective studies of PDE4D in different populations with larger sample numbers may help clarify the role of PDE4D in ischemic stroke and sub-types.
Conclusion
We report new findings that SNP rs152312 and SNP 45 are associated with the cardioembolic stroke sub-type (P<0·05) in a mainly Caucasian population from South Australia. However, we found no association of SNP 13, SNP 19, SNP 83 or SNP 87 with ischemic stroke in our study. This is the second Australian study to investigate PDE4D and ischemic stroke (19). Our findings support the original Icelandic study suggesting that SNP 45 in the PDE4D gene may be an important association in the cardioembolic sub-type of ischemic stroke (5). These findings add to the body of evidence supporting the hypothesis that PDE4D may contribute to the risk of cardiogenic stroke. We propose that the results from our Australian study support the concept that a large, sufficiently powered, prospective international study is required to elucidate the role of PDE4D in cardioembolic ischemic stroke.
Footnotes
Acknowledgements
Dr John B. F. Field, Statistician, Health Sciences, University of Adelaide, for statistical analysis of the results The Department of Molecular Pathology, Institute of Medical and Veterinary Science, Adelaide, South Australia, for DNA sequencing services, and The Departments of Medicine (Prof. R. E. Ruffin) and Neurology (Dr M. K. Robinson), University of Adelaide and The Queen Elizabeth Hospital, respectively, for support.