
Abstract
Background
High glucocorticoid serum levels and prednisone (PDN) therapy have been associated with depression, posttraumatic stress disorder, and some types of cognitive dysfunction in humans.
Objective
The aim of this study was to assess whether chronic (90 days) PDN administration produces disturbance in learning and memory retention associated with neuronal degeneration and cerebral glial changes.
Methods
Male Wistar rats were studied. Controls received 0.1 ml distilled water vehicle orally. The PDN group was treated orally with 5 mg/kg/d PDN, which is equivalent to moderate doses used in clinical settings. Learning and memory retention were assessed with the Morris water maze. The index of degenerated neurons as well as the number and cytoplasmic transformation of astrocytes and microglia cells were evaluated in the prefrontal cortex and the CA1 hippocampus.
Results
PDN-treated rats showed a significant delay of 20% in learning and memory retention as compared with controls. In addition, in the PDN group, the neuronal degeneration index was two times higher in the prefrontal cortex, and approximately 10 times higher in the CA1 hippocampus, than in control animals. The number and cytoplasmic transformation of astrocytes were also significantly higher in the PDN group than in control animals. In the PDN-treated group, isolectin-B4-labeled microglia cells were higher in the prefrontal cortex but not in the hippocampus.
Conclusion
These results suggest that chronic exposure to PDN produces learning and memory impairment, reduces neural viability, and increases glial reactivity in cerebral regions with these cognitive functions.
Introduction
Sixty-six years have passed since Wintersteiner, Pfiffner, and Mason discovered cortisone, and 53 years have gone by since Hench reported its dramatic effect on a patient with rheumatoid arthritis. Even with the dramatic advances in steroid chemistry and biology that have occurred since then, important evidence about the role of natural and synthetic glucocorticoids on brain physiology and disease is still emerging. For instance, high cortisol levels are associated with reduced hippocampal formation volume and memory dysfunction in patients with Cushing's syndrome. 1 Decreased cortisol level reverses hippocampal atrophy after treatment of Cushing's disease. 2 Plasma adrenocorticotropic hormone and cortisol levels are increased and correlated with the severity of hippocampal atrophy and memory impairment in several disorders, including Alzheimer's disease, schizophrenia, depression, and dementia associated with aging.3,4
Prednisone (PDN) is a synthetic glucocorticoid that is used widely to treat patients with many acute and chronic medical conditions. Studies have suggested that PDN may directly affect the brain by participating in the development of cognitive dysfunction, cerebral atrophy, depression, and anxiety, as well as the well-known effect of steroid psychosis. For instance, the acute administration of high-dose PDN or dexamethasone to healthy volunteers produced impairment in some aspects of cognitive functioning.5,6 Apparent cerebral atrophy on computed tomographic scans has been found in individuals who were taking steroids. 7 Patients with asthma, Crohn's disease, and other systemic conditions had mild subclinical psychiatric disturbance and selective neuropsychological deficits associated with steroid treatment.8,9 Some mood disturbances were also found in PDN-treated children with asthma. 10 It is also known that a subgroup of patients with systemic lupus erythematosus with no history of neurologic involvement may develop cognitive dysfunction after several years with the disease. 11 Although controversial, 12 it is our clinical impression that PDN treatment may produce this effect.
In view of obvious ethical and methodologic difficulties, we chose a pharmacologic approach with the use of rats as a first step to evaluate the effect of PDN on cognitive function and brain structure. We previously showed that subacute exposure to PDN induced anxiety-related behavior and reactivity of microglia. 13 In this study, we assessed the effects of chronic administration of PDN on memory acquisition and retention, neuronal viability, and reactive gliosis in rats. The dose of PDN administered to the study group is equivalent to moderate doses used in clinical settings, according to an interspecies scale. Memory acquisition and retention were assessed with the use of a common behavioral test. The brain regions considered for analysis were those with higher glucocorticoid receptors, such as the CA1 hippocampus and the prefrontal cortex. 14 Histologic assessments of neuronal damage and immunocytochemical analyses of astrocyte and microglial populations were performed in these regions. Astrocytes and microglia cells were selected because they are the first ones to respond to neuronal damage and play an important role in restoring injured nervous tissue.15–17
Methods
Experimental Animals
Experiments were performed on 8-week-old male Swiss-Wistar rats weighing 120-140 g. All animals were housed at 22±1°C under 12-h light-dark cycle conditions and were allowed free access to food and tap water. Two groups (15 rats/group) were assembled to assess memory acquisition and retention, neuronal degeneration, and glial reactivity. PDN-group received PDN (Sigma Chemical Co., St. Louis, Mo) 5 mg/kg/d orally, dissolved in 0.1 ml distilled water, every morning starting at 8:00 am for 90 days. We used this dosage because we wanted to assess the effects of moderate doses of PDN, similarly to those prescribed for several clinical conditions. According to an interspecies scale, a moderate dose of PDN in humans (45 mg/d) corresponds to 5 mg/kg/d in rats. This interspecies scaling takes into account mg/kg/U of physiologic time (mg/kg/pt). 18 The control group received 0.1 ml distilled water orally every morning, starting at 8:00 am, for 90 days. The same person administered PDN and water, and all animals were handled in similar conditions. All experimental procedures were performed in accordance with the institutional guidelines on animal care of the Centro de Investigacion Biomedica de Occidente.
Memory Tests
At the end of the administration period, the Morris water maze was used to assess memory acquisition and retention, as previously described.19,20 Briefly, the water maze is a circular tank of 1.80 m diameter and 0.48 m height filled with water (27°C) to a depth of 25 cm. It contained a 10x25-cm immersed platform placed at the center of a given quadrant. On the first day of the test (learning phase), the animals try to find and remember the correct position of the immersed platform. This phase consisted in eight swimming trials starting from four different positions that were equally distributed around the perimeter of the maze. After the animals found the platform, they were allowed to stay on it for 20 seconds and then placed into a warm cage for 30 seconds, until the start of the next trial. The recorded variables were the time required to localize the escape platform, swimming distance, and navigation velocity. After training, the animals were returned to their home cages. Twenty-four hours later, memory retention was assessed without the platform, allowing rats to swim for a single 60-second trial. The registered variable was the time spent in the quadrant where the platform was located. All tests were videotaped and analyzed by an independent observer who was blinded to group assignment.
After memory assessment, all animals were anesthetized with an intraperitoneal injection of pentobarbital (50 mg/kg) and killed by intravascular perfusion. At this time, blood samples were collected for serum glucose and electrolyte determination (sodium, potassium, and chloride). Afterward, 150 ml saline solution was instilled, followed by 150 ml 0.4% paraformaldehyde fixative in 0.1 M phosphate-buffered saline (PBS; pH 7.4). After perfusion, the brains were removed and immersed in the same fixative at 4°C for 24 h.
Degenerated Neurons Assessment
To assess neuronal damage in the prefrontal cortex and the CA1 hippocampus, a technician took five brains from each group at random to be embedded in paraffin, and coronal slides (4 μm) were cut at -0.7 to -3.8 mm from bregma 21 and stained with hematoxylin and eosin. Neuronal degenerative changes were identified by light microscopy as exhibiting at least one of the following characteristics: pyknosis, karyolysis, karyorrhexis, and cytoplasmic eosinophilia. Shrunken, dark, and scalloped neurons were also taken into account; 22 120 microscopic fields per brain and per cerebral region were assessed under x400 magnification. An observer who was blinded to group assignment performed the histologic assessment, counting the number of degenerated neurons and the total number of neurons per field (approximately 0.78 mm2). The neuronal degeneration index per field was calculated by dividing the number of degenerated neurons by the total number of neurons per field, multiplied by 100.
Astrocytes and Microglial Assessment
Five other brains were processed to identify astrocyte and microglial cells. Brains were cut using a vibratome, and 30-μm coronal sections were obtained at -0.7 to -3.8 mm from the bregma 21 and kept at 4°C in 0.1 M PBS. Immunocytochemical identification of astrocytes was performed as follows. Washes and incubations were performed on free-floating tissue sections under moderate shaking. Tissue sections were incubated in 1% hydrogen peroxide and 10% methanol for 20 minutes to block endogenous peroxidase staining. Slices were then washed twice for 20 minutes with 0.1% bovine serum albumin and 0.1% Triton X-100 in 0.1 M PBS. This buffer also was used for the following washes and incubations. Sections were then incubated overnight at 4°C with an anti glial fibrillary acidic protein (GFAP) polyclonal antiserum (Dako, Carpinteria, Calif) at a dilution of 1:800. Sections were then washed twice in buffer and incubated 2 hours at 22°C with antirabbit immunoglobulin G (Dako) at a dilution of 1:250. Peroxidase-antiperoxidase system incubation was performed in darkness for 2 hours (Dako) at a dilution of 1:200. Peroxidase activity was revealed with 0.01% hydrogen peroxide using 3'3-diaminobenzidine as chromogen. The remaining brains (four controls and two PDN) were kept apart for further ultrastructural analysis.
Histochemical staining of microglia was performed on free-floating sections under gentle shaking. To inhibit endogenous peroxidase activity, sections were incubated for 20 minutes with 10% methanol and 1% H2 O2. Tissues were rinsed in 0.1 M PBS and incubated for 10 minutes in a cationic solution (0.1 mM CaCl2, 0.1 mM MgCl2, and 0.1 mM MnCl in 0.1 M PBS; pH 7.4). Next, they were incubated overnight at 4°C with peroxidase-labeled isolectin-B4 (Sigma Chemical Co.) at a 1:100 dilution. The reaction was revealed with 3'3-diaminobenzidine. All sections were processed simultaneously with the use of immunocytochemical and histochemical methods.
Immunostained sections were studied with an imaging analyzer (Leica Q5001W; Leica Imaging Systems, Ltd., Cambridge, UK). The astrocyte population was quantified within 120 microscopic fields/rat for each group with the use of x400 magnification (approximately 0.78 mm2 /field). To study the cytoplasmic transformation (expressed as the number of intersections), 600 cells per cerebral area for each group were assessed with a stereologic grid according to the Weibel point-counting method 23 and modified as follows: the test grid consisted of five concentric circles with a 90-μm separation between them. The astrocyte nuclei were placed at the center of the circle, and the number of points at which immunoreactive profiles crossed the test grid lines was recorded. Only cells and immunopositive branches seen in the same plane of focus were considered for quantification. Microglial evaluation was performed in the same way as it was for astrocytes. An observer who was blinded to group assignment performed the glial assessment.
Statistical Analysis
The results are expressed as mean±SEM. Differences between continuous variables in the morphometric analysis were determined by performing Student's t test. Differences between continuous variables in the cognitive assessments were determined by using the Mann-Whitney U test. The confidence intervals (CIs) for differences are reported at 95% (i.e., 95% CI). Statistical significance values were set at <0.05. Intraobserver variability was assessed with the use of Pearson's correlation in 25 randomly selected fields read twice 3 days apart by the same observer who was blinded to group assignment.
Results
Four rats were excluded from the behavioral tests and histologic analysis: one from the control group because it was unable to swim, and three from the PDN group because they showed incipient bacterial bronchitis during the routine necropsy performed in all studied animals. Serum levels of glucose, sodium chloride, and potassium in the PDN group were not different from those in controls (data not shown).
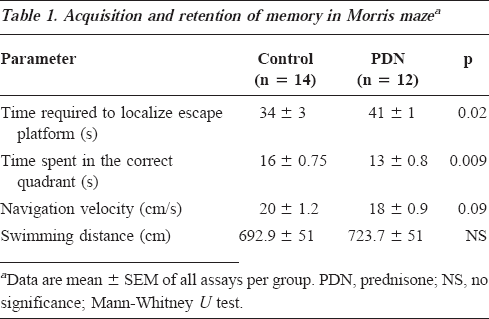
Memory acquisition and retention results are shown in Table 1. There was a significant delay of 20% in the learning phase, as assessed by the time spent to localize the escape platform, in the PDN exposed rats compared with controls (95% CI for differences -13 to -0.8 seconds). PDN-exposed rats also performed worse in memory retention, as assessed by the time spent in the correct quadrant, compared with controls (95% CI for differences 0.9-5 seconds). It seems that locomotor performance, as assessed by navigation velocity (95% CI of means for PDN group 15-19 cm/s vs. control group 17-22 cm/s) and swimming distance (95% CI of mean for PDN group 611-836 cm vs. control group 581-802 cm), was not modified by PDN administration. The power of the test 24 to assess differences in behavioral variables was 0.83.
Acquisition and retention of memory in Morris mazea
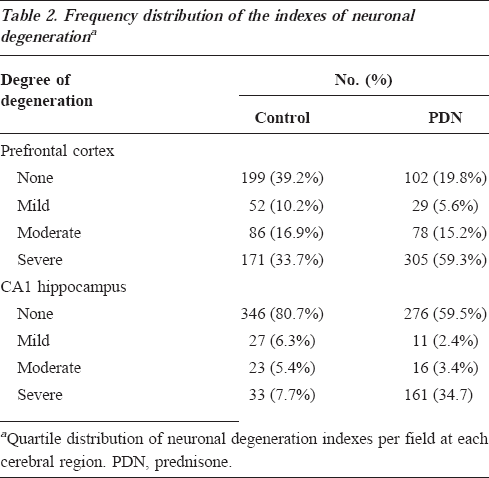
Neuronal damage, as assessed by the neuronal degeneration index, was two times higher for the PDN group in the prefrontal cortex (PDN group, 10±0.5, 95% CI, 9-11; control group, 5±0.3, 95% CI, 5-6, p<0.001, 95% CI for differences, -6 to -3), and approximately 10 times higher in the CA1 hippocampus (PDN group, 5±0.5, 95% CI, 4-6; control group, 0.5±0.6, 95% CI, 0.37-0.59, p<0.001; 95% CI for differences, -6 to -4) compared with controls. The neuronal degeneration index was arbitrarily categorized as none, mild, moderate, and severe degeneration based on quartile distribution. The PDN group showed almost two times more severe degeneration in the prefrontal cortex and almost five times more severe degenerations in the hippocampus compared with controls (p<0.001) (Table 2). Intraobserver variability was 0.87. Figure 1 shows neurons with degenerative signs.
Frequency distribution of the indexes of neuronal degenerationa
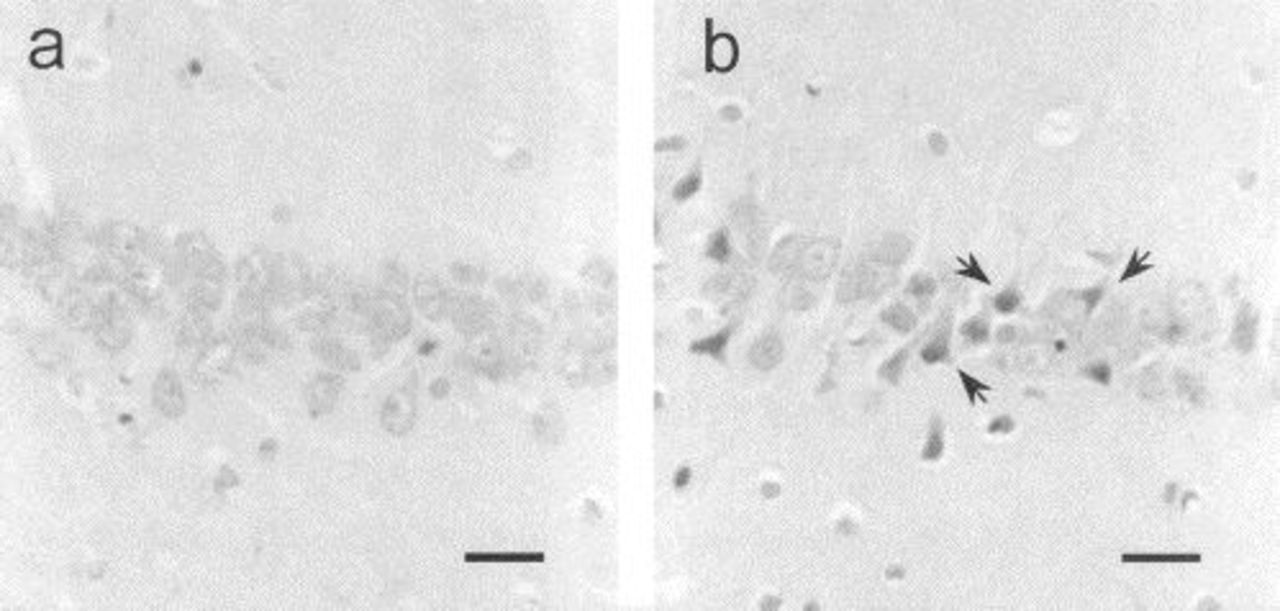
Effects of PDN on neuronal viability. (
The results of the morphometric analysis of astrocytes are illustrated in Table 3. The immunocytochemical reactivity of these cells was increased in the PDN-exposed rats compared with controls. The PDN group had a significantly higher number of astrocytes and cytoplasmic intersections in the prefrontal cortex and the CA1 hippocampus.
Glial fibrillary acidic protein immunoreactive astrocyte population analysisa
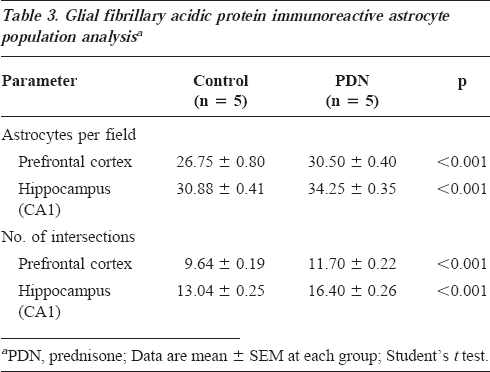
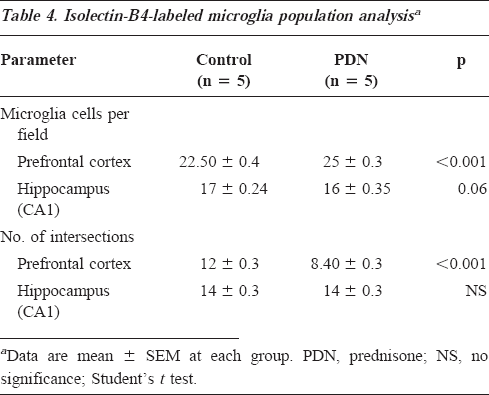
Table 4 shows the number of isolectin-B4-stained microglial cells and their cytoplasmic intersections per cerebral region. The PDN group had a significantly higher number of stained microglial cells in the prefrontal cortex (95% CI for differences, -3.3 to -1.4) and a borderline difference in the CA1 hippocampus (95% CI for differences, -0.039-1.6) compared with controls. In addition, microglial cytoplasmic transformation, as assessed by the number of intersections, was significantly higher in the PDN group in the prefrontal cortex (95% CI for differences, 3-4), but not in the CA1 hippocampus, compared with controls.
Isolectin-B4-labeled microglia population analysisa
Discussion
The Morris water maze is a reliable and commonly used test to assess the acquisition and retention of spatial memory in rodents. 19 With the use of this test, we have shown that chronic administration of a moderate dose of PDN impairs learning and memory retention. The locomotor performance in the study group was not impaired, suggesting that PDN did not have an effect on muscle function. The possibility of a Type II error is low, as shown by the power of the test and the overlap of the 95% CIs in the navigation velocity and swimming distance variables.
Our results coincide with those of other clinical reports in which PDN treatment was associated with cognitive dysfunction in humans.5–10 PDN could induce these cognitive disturbances through the same mechanism described for natural glucocorticoids. High glucocorticoid levels may affect the synthesis of 5-hydroxytryptamine (i.e., serotonin). 25 This neurotransmitter is associated with the adequate performance of cognitive functions such as learning, memory, and attentional processes. 26
Our results also showed that neuronal viability in the prefrontal cortex and the CA1 hippocampus was reduced as a consequence of PDN exposure. These degenerative changes are coincident to those found in other conditions that exhibit high glucocorticoid levels and would explain the hippocampal volume reduction.27,28 The relevance of establishing the precise moment at which the loss of neurons occurs is very important, because the reversible atrophy induced by glucocorticoids27,28 might be treatable at the time it is discovered, but the neuronal degeneration not. The neuronal loss induced by PDN can be due to glucose uptake disturbances.29,30 Other possible mechanisms include increasing synaptic glutamate concentrations, neuroexcitotoxicity, and alteration of neurotrophin expression.3,31
Assessment of astrocyte reactivity has been used as an indicator of cerebral tissue damage; this response is characterized by a rapid increase in the expression of GFAP.15,16 Our results revealed that PDN increased the GFAP immunoreactivity at prefrontal cortex and CA1 hippocampus. Neuronal degenerative changes were also present in both cerebral regions suggesting that GFAP overexpression can be related to astrocytes reactivity in responding to neural injury. Yet the upregulated expression of GFAP as a consequence of a decreased tryptophan hydroxylase mRNA cannot be excluded.32,33
It has been reported that dexamethasone increases microglia ramification in fetal rat brains. 34 In addition, our group previously reported that subacute exposition to PDN produce similar changes. 13 In this study, chronic exposure to PDN produced changes in the number and cytoplasmic transformation of isolectin-B4-stained microglia cells in the prefrontal cortex, suggesting an overexpression of galactosyl glycoproteins on microglia. 34 This assumption is supported by the fact that the concentration of isolectin B4 used in our study (1:100) stains mainly activated microglia. 35 A possible trigger for microglial response could be a neuronal overstimulation produced by glutamate excitotoxicity, as reported with high glucocorticoid levels.3,31 Yet other studies have found a downregulation of microglia induced by different glucocorticoids. 36 This apparent discrepancy in microglia transformation can be explained by differences in compounds and doses used, which may produce opposite effects, acting as inhibitors through glucocorticoid receptors or stimulators via mineralocorticoid receptors. The regional microglial changes between prefrontal cortex and CA1 hippocampus found in this study may also be due to dissimilar glucocorticoid receptor densities.3,14,37
Conclusions
Chronic exposure to PDN produced memory impairment and glial reactivity in rats. In addition, this study provides new evidence relating glucocorticoid therapy with neuronal loss. These features support further studies to assess whether PDN may produce similar effects in humans, mainly in light of the twofold increase in PDN and other glucocorticoid sales in North America during the past 5 years (data provided by Undra Farmaceutica, Schering-Plough de México, from IMS Health).
Footnotes
Acknowledgment
The authors were supported by Fondo de Fomento a la Investigacion Project FP0038/1191 of the Instituto Mexicano del Seguro Social, México.