
Abstract
Background
Proinflammatory and immunomodulatory adipokines are linked to inflammation in critically ill patients but are poorly studied after injury. We hypothesized that trauma would induce systemic adipokine release and influence erythroid suppression.
Methods
Blood and bone marrow (BM) were collected from trauma patients (ISS > 15, n = 90) and compared to patients undergoing elective hip replacement (n = 37). Plasma adipokine levels were measured, and BM was assayed for adipokine transcription and erythroid progenitor growth potential. Differences were detected using t-tests and correlations using simple linear regression.
Results
Trauma patients exhibited decreased adiponectin (1.8* vs 3.4 mg/mL) and increased leptin (7.8* vs 4.6 ng/mL) and resistin (3.1* vs 2.5 ng/mL), with sex- and age-specific differences. They also showed increased BM visfatin transcription. Adipokine transcription negatively correlated with erythroid progenitor growth.
Conclusion
Adipose tissue activity is linked to inflammatory responses after injury, with variability by age and sex. Bone marrow adipose tissue may influence erythroid recovery after trauma.
Key Takeaways
• Trauma patients had decreased plasma adiponectin and increased leptin and resistin. • Trauma patients expressed higher bone marrow visfatin gene transcription. • Bone marrow adipokine expression correlated with low erythroid progenitor growth.
Introduction
Bone marrow adipocytes are metabolically active cells that maintain close interactions with endothelial cells, maturing hematopoietic cells and erythroblast-supporting macrophages, and sympathetic nerve terminals. 1 These cells are important providers of free fatty acids and both local and systemic signaling factors known as adipokines. 1 These include leptin, adiponectin, resistin, and visfatin and serve various regulatory functions within the bone marrow, including energy storage, endocrine function, and bone metabolism. 2 Changes in adipokine secretion can alter the bone microenvironment, progenitor cell growth, survival, and mobilization as well as potentially fueling bone destruction. 2
In plasma, adiponectin, resistin, and visfatin have been shown to play a role in both pro- and anti-inflammatory signaling. 2 Sepsis, burns, and critical illness can induce changes in the structure and activity of white adipose tissue, leading to changes in adipocyte size, collagen deposition, mitochondrial density, and overall adipokine release. 3 Plasma levels of proinflammatory resistin, visfatin, and leptin have been found to be elevated in sepsis and critical illness and associated with an increased inflammation, organ failure, and mortality.3-5 Conversely, adiponectin has predominantly an anti-inflammatory function, with lower levels in critically ill patients being linked to poor outcomes.6-8
Chronic inflammation and a hypercatecholamine state have been shown to influence hematopoietic dysfunction after injury.9,10 After traumatic injury, hematopoietic stem and progenitor cells in the bone marrow respond to cytokines like interleukin-6 (IL-6), interferons, and tumor necrosis factor-alpha (TNFα) that restrict stem and progenitor cell differentiation, as well as signals like granulocyte colony-stimulating factor that push bone marrow predecessors toward granulocyte formation.11,12
Although plasma adipokines have been studied in the setting of critical illness, their unique role within the bone marrow remains poorly understood. This study sought to investigate the role of these 4 major adipokines within the bone marrow after injury and examine correlations with bone marrow erythropoietic function. We hypothesized that severe trauma would lead to elevations in proinflammatory adipokines that would be associated with erythroid growth suppression.
Methods
Study Population
We performed a prospective observational cohort study comparing severely injured blunt trauma patients (n = 90) to patients undergoing elective total hip replacement (THR) (n = 37). Patients undergoing elective THR served as operative controls to control for anesthesia exposure, blood loss, and orthopedic manipulation at time of bone marrow collection. Trauma patients were screened on admission for inclusion in the study, with prior approval by the Institutional Review Board (IRB201601386). Inclusion criteria for the blunt trauma cohort were as follows: age ≥18, lower extremity long bone or pelvic fracture requiring open reduction and internal fixation, hemorrhagic shock at time of admission (defined by systolic blood pressure ≤90 mmHg, or mean arterial pressure ≤65 mmHg, or base deficit ≥5 mEq/L or lactate ≥2 mmol/L), and an injury severity score (ISS) ≥ 15. Exclusion criteria included survival <48 hours, incarceration, pregnancy, patients receiving chronic corticosteroids or immunosuppressive therapies, previous bone marrow transplantation, or end stage renal disease. This study was registered at ClinicalTrials.gov (NCT02577731). All THR patients and non-intubated trauma patients gave written informed consent for the research procedures. For those critically ill trauma patients who were not able to provide written informed consent for tissue sampling, written informed consent was obtained from their surrogate decision maker.
Demographic data included age, sex, admission vital signs, admission body mass index (BMI), intensive care unit (ICU) length of stay, and hospital length of stay, time from injury to bone marrow collection, and complications. Blood loss during the time of hip replacement or fracture fixation surgery was documented as estimated operative blood loss. Total hospital blood loss was calculated as the sum of documented blood loss from any surgeries or procedures that occurred during admission.
Bone Marrow and Blood Collection and Processing
Trauma bone marrow samples were obtained using an 11-gauge bone marrow biopsy needle (Argon Medical Devices, Frisco, TX) aspirated from either the ilium, femur, or tibial plateau under general anesthesia during open reduction and internal fixation of the pelvis, acetabulum, femur, or proximal tibia. During THR, bone marrow was aspirated from the femur.
Each bone marrow sample was passed through a 70 μm cell strainer into a 50 mL conical tube. Erythrocytes were lysed by adding at least 3 volumes of erythrocyte lysis buffer (Qiagen, Hilden, Germany). The mixture of bone marrow and lysis buffer was gently mechanically agitated for 5 minutes and centrifuged for 10 minutes at 300 g. After the second centrifugation, the supernatant was again aspirated, and the resulting white pellet was suspended in 1 mL Iscove’s Modified Dulbecco’s Medium (IMDM).
Cells in the resulting solution were counted using a TC20 automated cell counter (Bio-Rad, Hercules, CA). Two duplicate 50 000-cell aliquots were each transferred into 1 mL H4034 optimum methylcellulose medium (STEMCELL Technologies Inc, Cambridge, MA) and plated in duplicate 33 × 10 mm Petri dishes. These were incubated at 37oC in 5% CO2. After 14 days, erythroid colony forming units (CFU-E), erythroid burst forming units (BFU-E), and granulocyte, erythrocyte, monocyte, and megakaryocyte colony forming units (CFU-GEMM) were manually counted by a blinded observer.
Blood samples were obtained from both cohorts on day of surgery in a fully filled heparinized 6 mL tube (Becton Dickinson, Franklin Lakes, NJ) and plasma was stored at −80oC. Plasma adiponectin, visfatin, resistin, and leptin levels were measured using immunosorbent assays according to manufacturers’ protocols (adiponectin and visfatin: Invitrogen, Waltham, MA; resistin and leptin: Abcam, Cambridge, United Kingdom).
RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction
Bone marrow RNA was isolated using the PureLink RNA Mini Kit (Thermo Fisher Scientific, Waltham, MA) following the manufacturer’s instructions and stored at −80°C. RNA was quantified using a BioTek Micro-volume plate on an EPOCH plate reader (BioTek Instruments, Winooski, VT). Ratio of absorbance at 260/280 nm was taken for RNA purity.
Samples were diluted using RNA free water to a final concentration of 2 μg RNA/10 μL. Following synthesis of bone marrow RNA, cDNA synthesis was performed using High-Capacity cDNA Reverse Transcription Kits (Applied Biosystems, Vilnius, Lithuania).
Real-time quantitative polymerase chain reaction was performed to analyze the expression of bone marrow adiponectin (ADIPOQ), visfatin (NAMPT/PBEF), resistin (RETN), and leptin (LEP) using Brilliant II SYBR Green QPCR Master Mix (Agilent Technologies, Santa Clara, CA) and the Mx3005P qPCR System (Agilent Technologies, Santa Clara, CA). Cycle thresholds for each gene were normalized against the housekeeping gene β-actin (ACTINB).
Primers were designed using Primer3 Web software: ACTINB forward 5′-aagcatcgggtggttagttg- 3′, reverse 5′-gctacggttatcccaaagca-3’; ADIPOQ forward 5′-agtctcacatctggttgggg-3′, reverse 5′- gtgcctctggttccacaaag-3’; NAMPTforward 5′-ttcggttctggtggaggttt-3′, reverse 5′- agacgttaatcccaaggcca-3’; RETN forward 5′-cctcaaagaaagagctgcgg-3′, reverse 5′- gacagggaggaggaggaga-3’; LEP forward 5′- accaaggtcttcagccatca-3′, reverse 5′- ccctctgccctctctgaaat-3’. Data reported as log2 fold change relative to operative controls using ΔΔCt method.
Statistical Analysis
All statistical analyses were conducted in GraphPad Prism version 9.1 (GraphPad Software, La Jolla, CA). Differences in categorical variables (race, sex, and presence of diabetes or complication) were identified using Fisher’s exact test. Differences in clinical parameters, gene expression, and erythroid progenitor growth vs THR were detected using unpaired two-tailed Welch’s t-tests and correlated with erythroid progenitor growth using simple linear regression. Subgroup analyses looking at age and sex were made using multiple unpaired two-tailed Welch’s t-tests. Data presented as median [interquartile range] or n (%).
Results
Study Population and Outcomes
Summary of Enrolled Patient Characteristics.
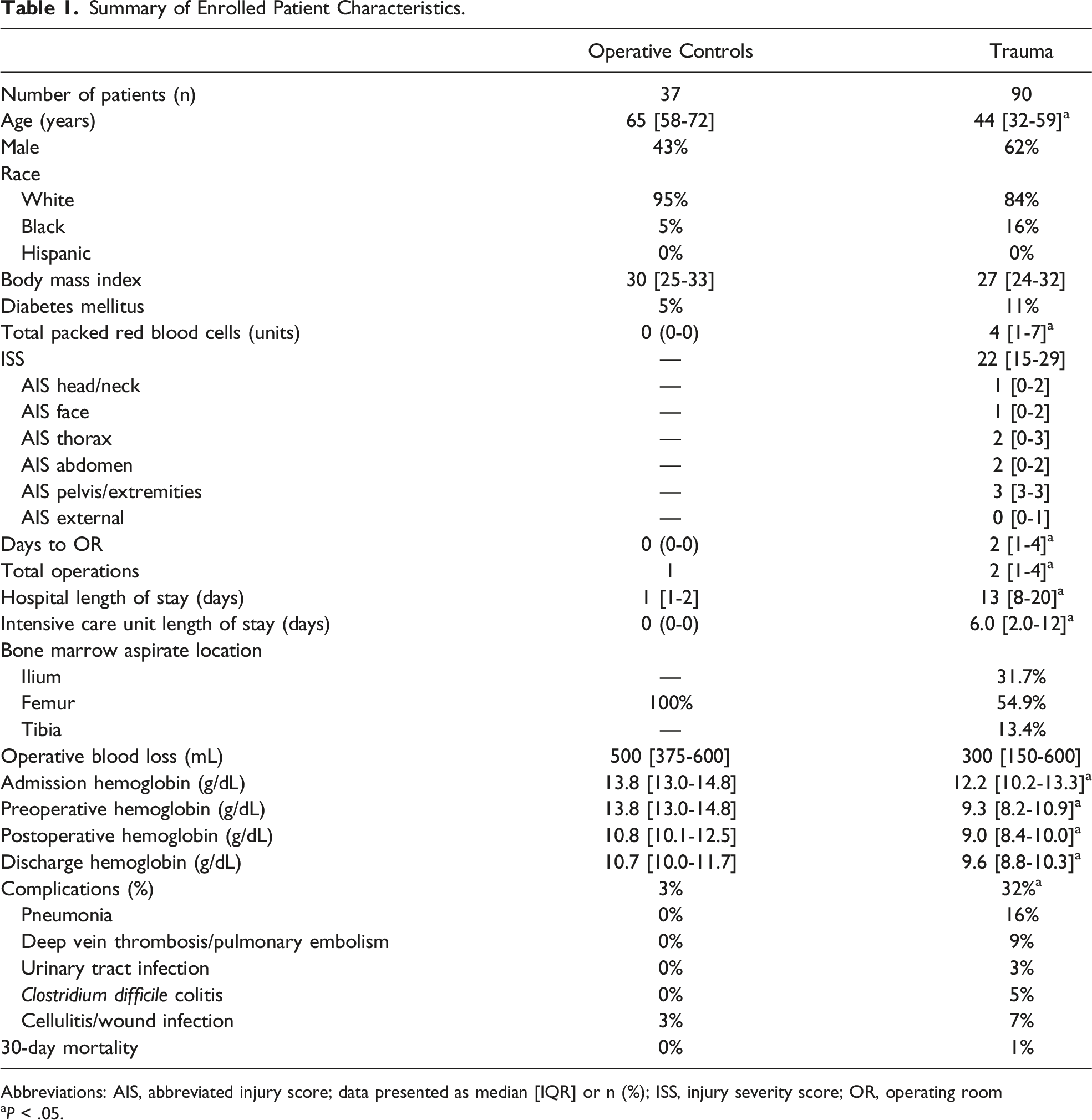
Abbreviations: AIS, abbreviated injury score; data presented as median [IQR] or n (%); ISS, injury severity score; OR, operating room
aP < .05.
Pneumonia was the most common complication in trauma patients.
Alterations in Plasma and Bone Marrow Adipokines After Trauma
Trauma patients exhibited decreased plasma levels of adiponectin (3.4 vs 1.8 μg/mL, P = .0001) (Figure 1A). Following trauma, there were increases in both plasma leptin (7.8 vs 4.6 ng/mL, P = .0002) and plasma resistin (3.1 vs 2.5 ng/mL, P = .0156). There was no significant difference in plasma visfatin levels between trauma and THR. There was no difference between transcription of ADIPOQ (adiponectin), RETN (resistin), or LEP (leptin) in bone marrow of trauma patients compared to THR (Figure 2). However, trauma patients saw a significant increase in bone marrow NAMPT (visfatin) transcription compared to THR (log2 fold change 3.3 [2.3-5.0], P < .0001). There was no correlation between plasma adiponectin, visfatin, resistin, or leptin and their bone marrow transcription. Trauma alters plasma levels of adiponectin (A), visfatin (B), resistin (C), and leptin (D) when compared to operative controls undergoing elective hip replacement. *P < .05. Differential mRNA expression of ADIPOQ, NAMPT, RETN, and LEP in the bone marrow of trauma patients compared to operative controls. ADIPOQ, adiponectin; NAMPT, visfatin; RETN, resistin; LEP, leptin; *P < .05.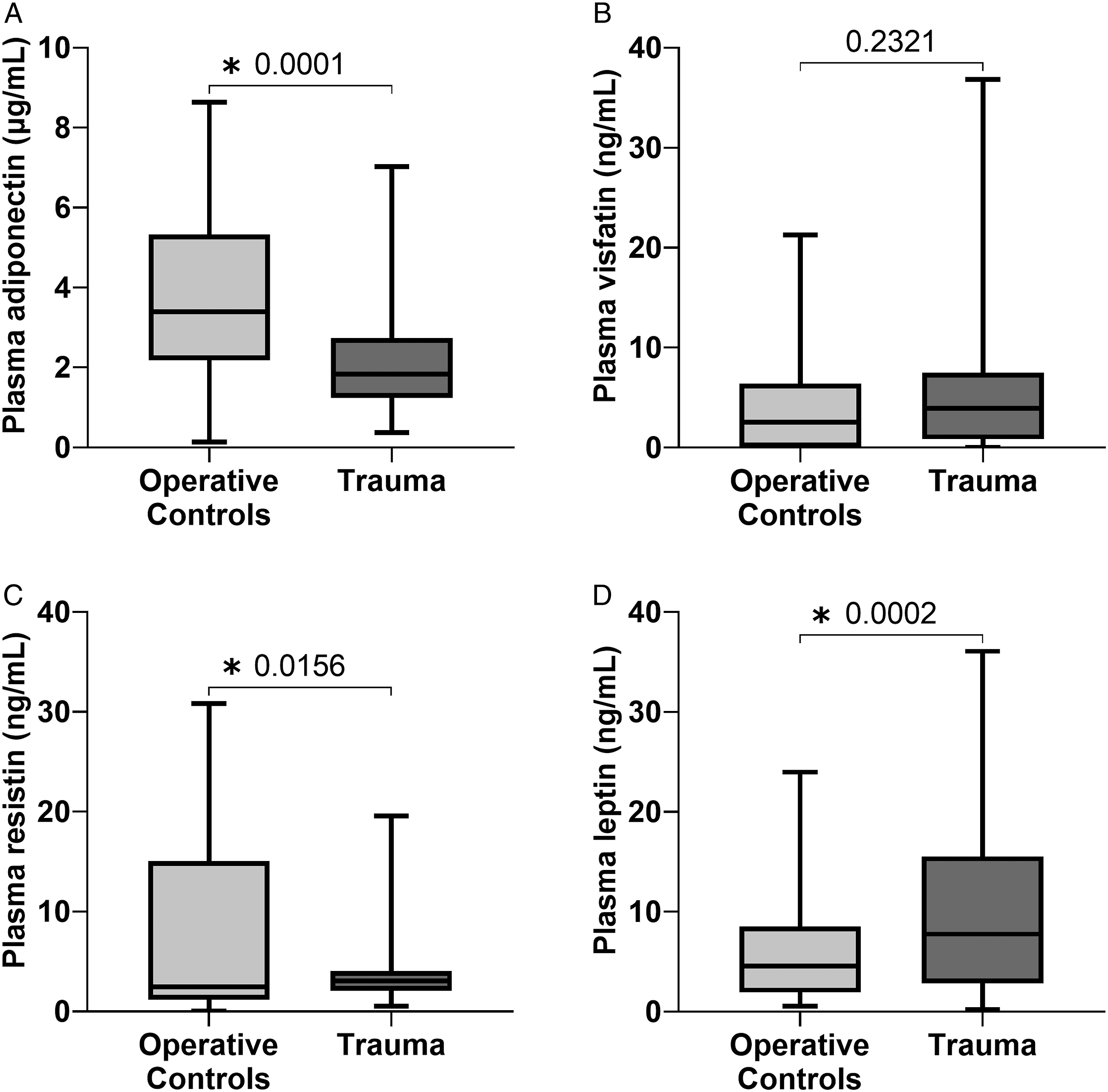
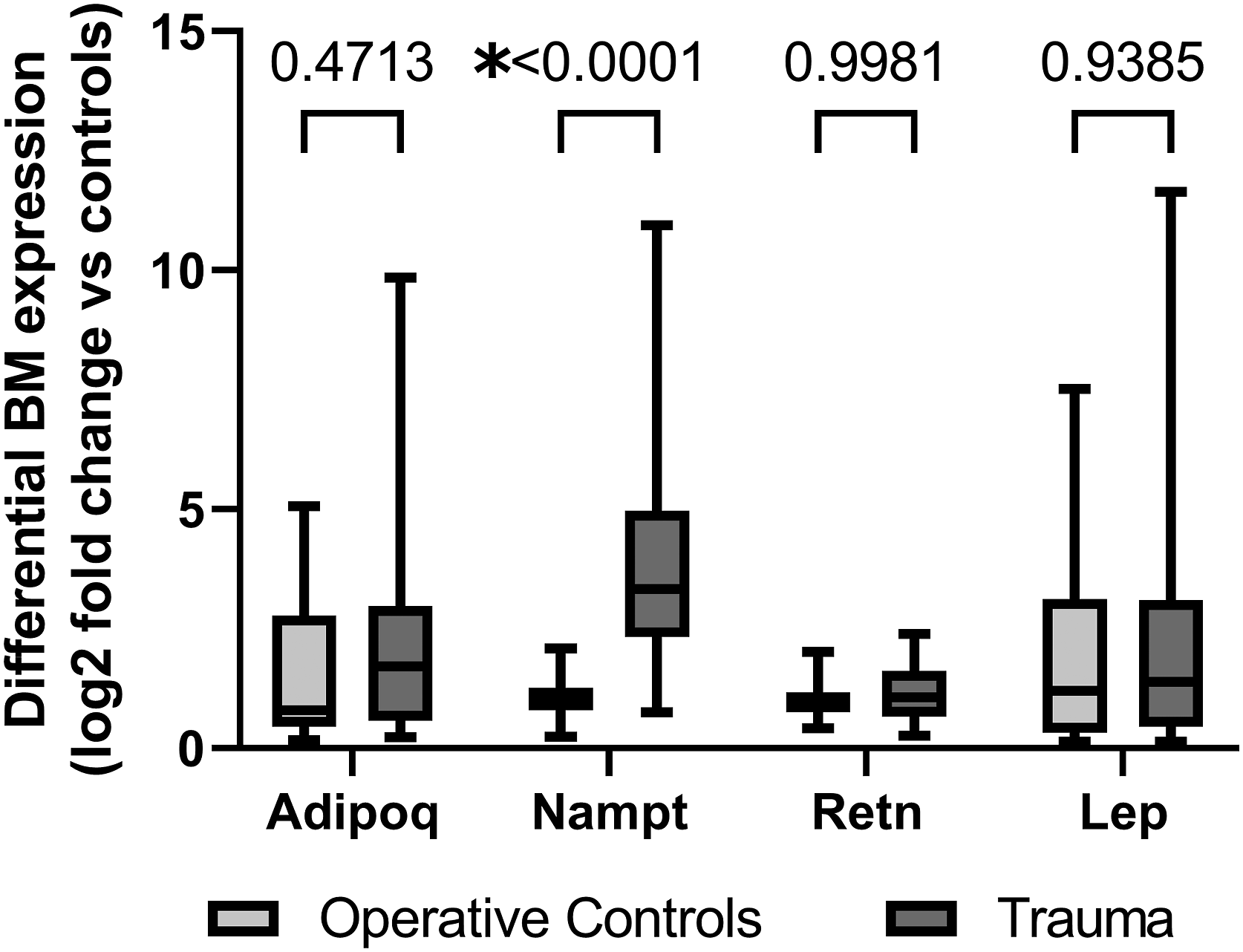
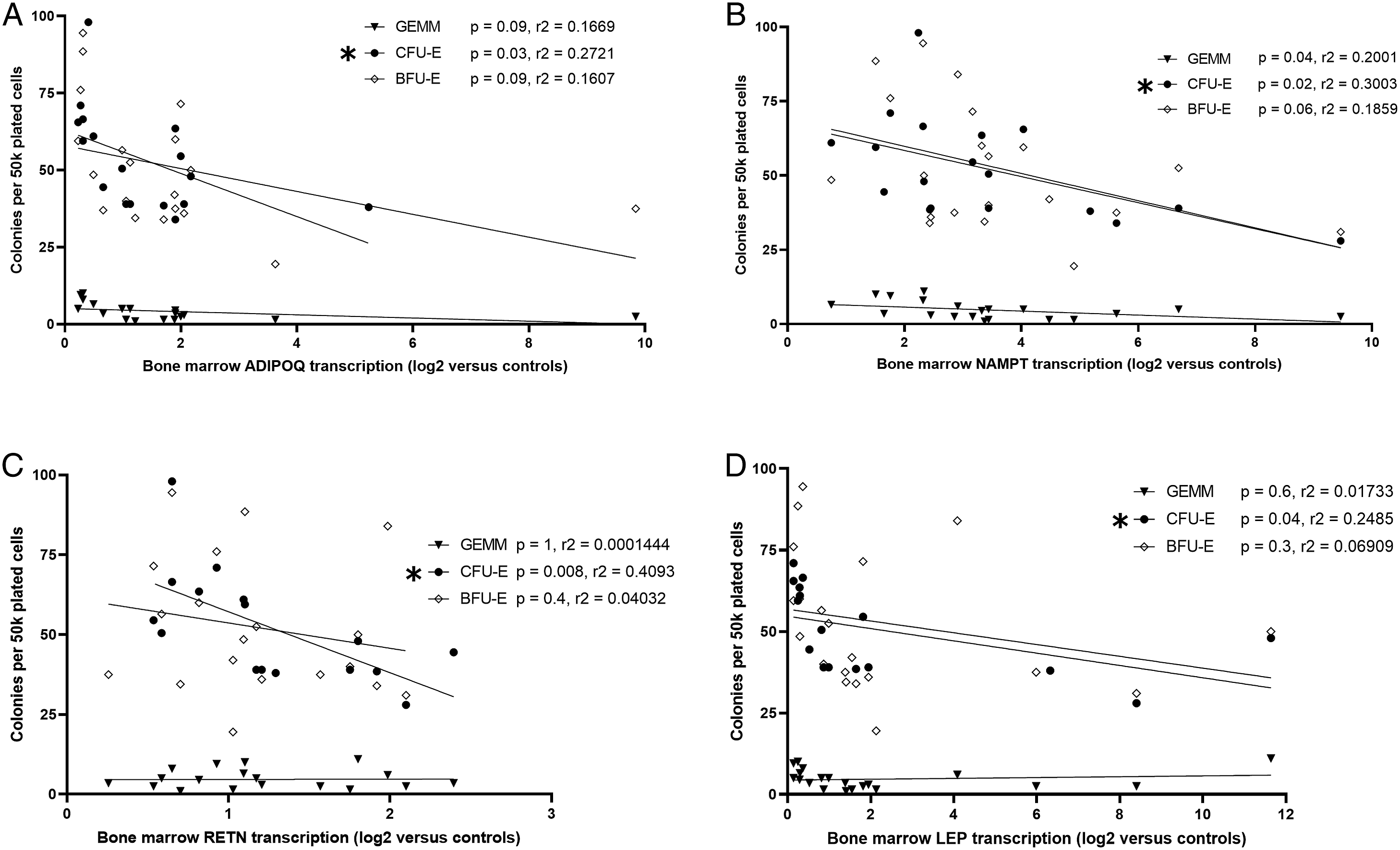
Correlation of Erythroid Progenitor Growth and Bone Marrow Adipokine Transcription
Trauma patients had decreased growth of all bone marrow erythroid progenitor colonies (CFU-GEMM, BFU-E, and CFU-E) compared to THR (Figure 3). Transcription of ADIPOQ, NAMPT, RETN, and LEP in trauma patients inversely correlated with CFU-E growth (Figures 4A-4D). Bone marrow transcription of ADIPOQ, NAMPT, RETN, and LEP did not correlate with decreased growth of CFU-GEMM or BFU-E. Trauma leads to significant erythroid progenitor suppression when compared to operative controls. CFU-GEMM, colony forming unit-granulocyte, erythrocyte, monocyte, and megakaryocyte; BFU-E, burst forming unit-erythroid; CFU-E, colony forming unit-erythroid; *P < .05. Negative correlations were noted between bone marrow transcription of ADIPOQ (A), NAMPT (B), RETN (C), and LEP (D) which significantly correlated with CFU-E growth in trauma patients. ADIPOQ, adiponectin; NAMPT, visfatin; RETN, resistin; LEP, leptin; CFU-GEMM, colony forming unit-granulocyte, erythrocyte, monocyte, and megakaryocyte; BFU-E, burst forming unit-erythroid; CFU-E, colony forming unit-erythroid; *P < .05.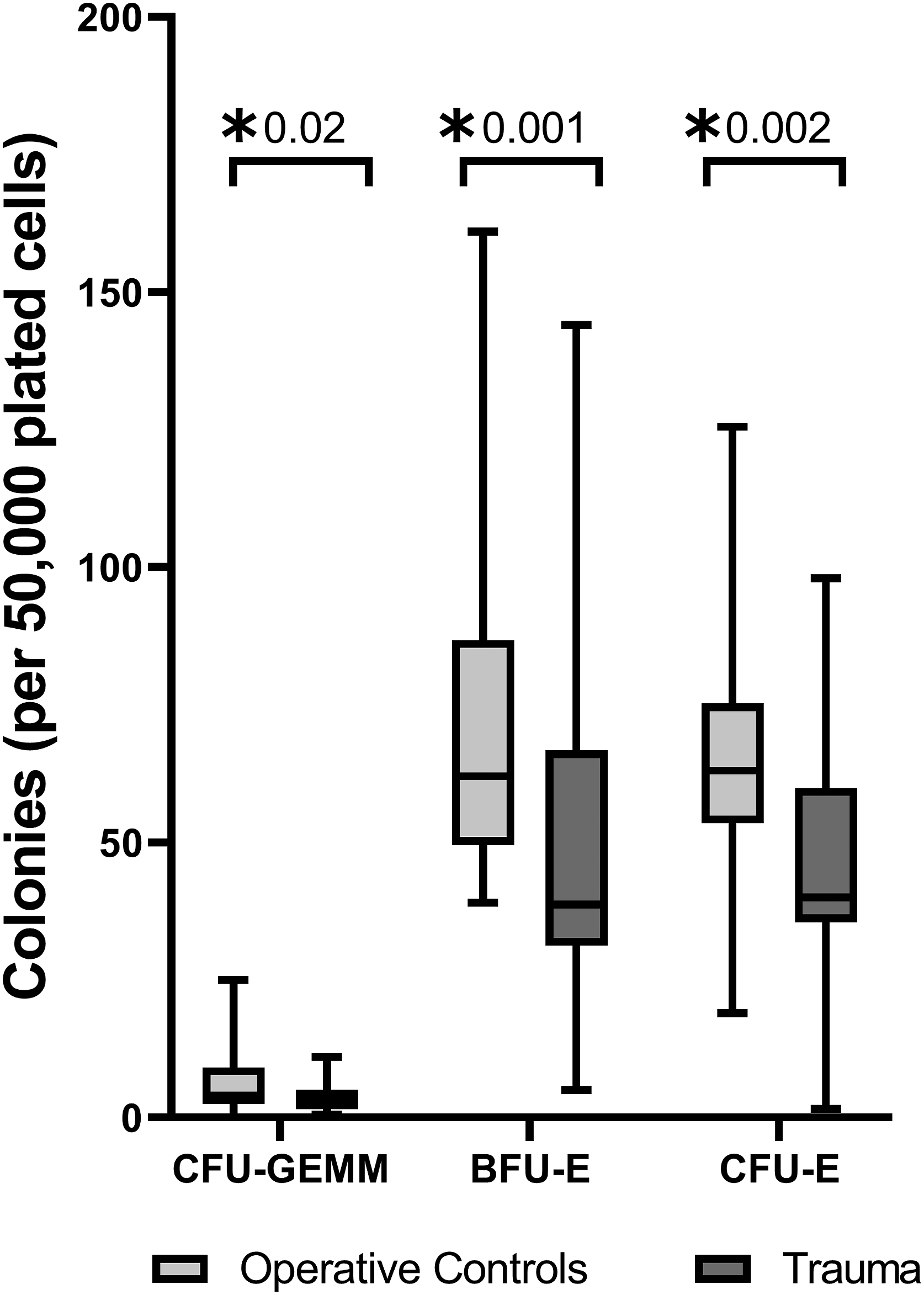
Adipokine Differences After Trauma Stratified by Sex and Age
Subgroup analyses were performed given known variations in adipokine release related to both sex and age. Both sexes demonstrated significant decreases in plasma adiponectin after trauma compared to THR (Figure 5A). Significant differences in plasma resistin and leptin between trauma and THR were only evident among females (Figures 5C-5D). Plasma visfatin levels demonstrated no differences when stratified by sex. Plasma levels of adiponectin (A), visfatin (B), resistin (C), and leptin (D) from trauma patients are compared to operative controls stratified by sex. *P < .05.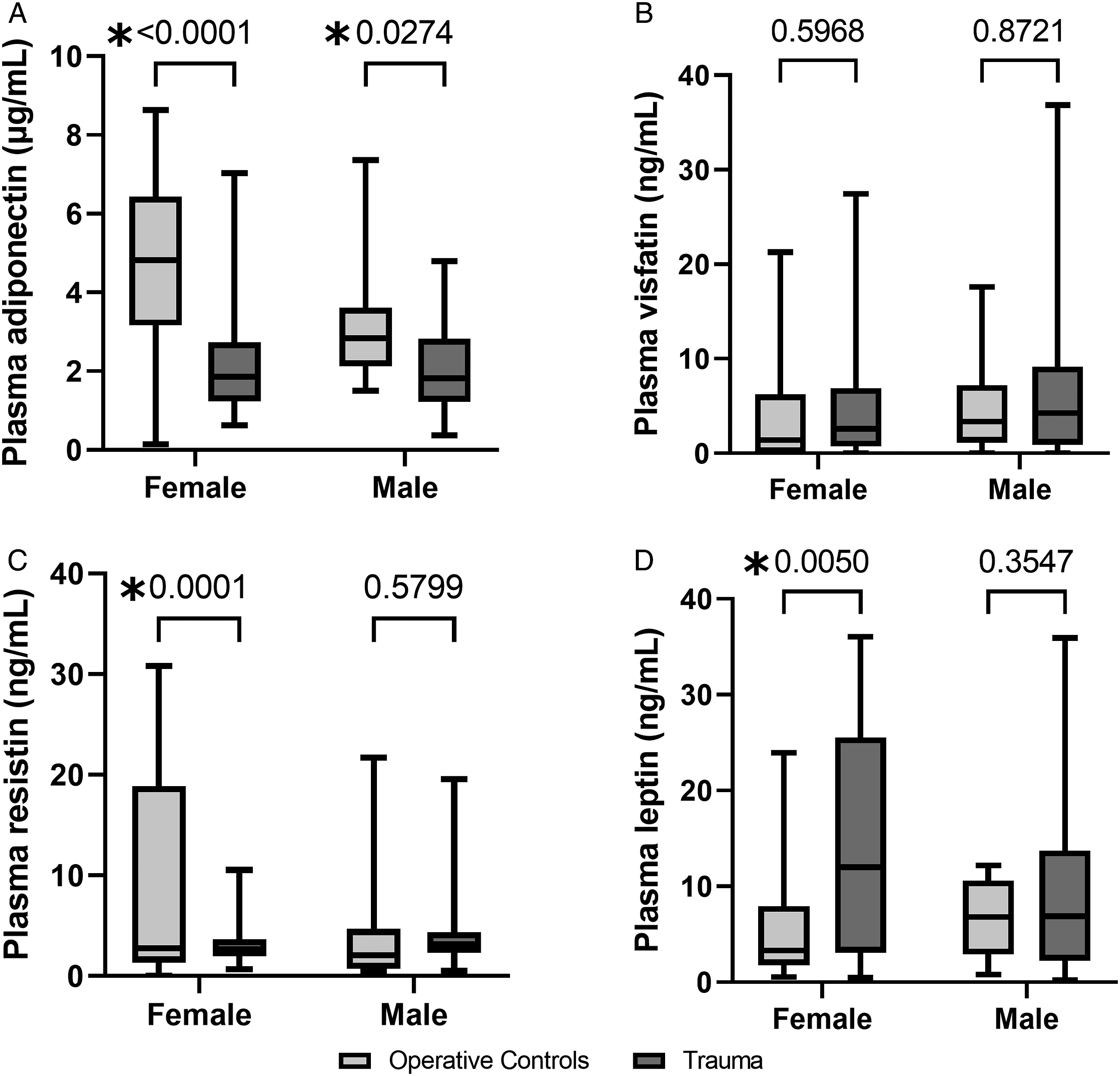
When stratifying by age, there was a difference in plasma adiponectin between trauma patients and THR for those under age 65 (1.7 vs 3.5 μg/mL, P < .0001), but these differences disappeared in those over age 65. However, differences in plasma leptin were significant only between trauma and THR over age 65 (8.3 vs 3.2 ng/mL, P = .02). There were no significant differences in visfatin or resistin levels when stratified by age.
Among trauma patients, plasma levels of resistin and leptin weakly correlated with a longer ICU length of stay (r2 = .1273, P = .0006; and r2 = .0554, P = .03, respectively). Trauma patients who developed complications had significantly higher plasma resistin compared to those who did not have a complication (3.8 vs 2.8 ng/mL, P = .009). There were no correlations identified between resistin, leptin, adiponectin, or visfatin and packed red blood cell administration, or injury severity.
Discussion
Proinflammatory leptin and resistin were significantly elevated among trauma patients, while anti-inflammatory adiponectin was decreased. In addition, trauma patients had a significant increase in bone marrow visfatin transcription. Bone marrow adipokine expression correlated with reduced CFU-E growth. By comparing severely injured trauma patients to THR, this study was able to identify differences in plasma adipokine levels as well as differences in adipokine transcription within the bone marrow. Both sex and age may play a role in levels of adipokines found after severe injury.
Peripheral white adipose tissue releases proinflammatory and anti-inflammatory cytokines like IL-6, TNFα, stem cell factor, and IL-1β.2,13 Despite no difference in BMI between trauma patients and THR, we found differences in circulating levels in 3 of the 4 adipokines tested after trauma. Ample evidence exists to support the roles of visfatin and resistin in the proinflammatory responses to sepsis and burns. 3 Human studies have repeatedly linked increased plasma resistin with release of proinflammatory and erythroid suppressive cytokines IL-6, TNFα, and IL-1β and worse outcomes.3-5 These findings correlate with what was found in our trauma patients. On the other hand, increased adiponectin is generally seen as a favorable indicator of improved outcomes and survival.6-8 Exogenous adiponectin administration led to reduced IL-6 and TNFα and the damage-associated high mobility group box protein-1 (HMGB-1) production and release as well as improved mortality in murine sepsis. 14 The low adiponectin levels we saw in our trauma patients may be a response to increased circulating inflammatory cytokines like TNFα, which is known to suppress adiponectin production from human adipocytes in vitro. 15 There is evidence to suggest that low adiponectin levels which rise over time may be more beneficial in patients with sepsis.7,15 Initial low adiponectin levels may be an early finding after injury and inflammation, and there may be a temporal component to its production.
In contrast to peripheral adipose tissue, the unique role of bone marrow adipocytes and impact of adipokines within the bone marrow is less well-studied. Evidence is mounting to suggest a role for bone marrow adipose tissue in the paracrine/endocrine regulation of bone marrow activities, including skeletal remodeling and hematopoiesis (Figure 6).1,16 Our study identified an increase in bone marrow transcription of visfatin alone after trauma. Bone marrow visfatin expression has been shown to influence bone growth and mineralization in the setting of widespread inflammation.
17
The lack of transcriptional changes in the bone marrow of other factors may imply differing functions between peripheral and bone marrow adipose tissue. Overview of the role of peripheral and bone marrow adipose tissue in trauma. IFNγ, interferon-gamma; TNFα, tumor necrosis factor-alpha; IL-1β, interleukin-1 beta; IL-6, interleukin-6; SCF, stem cell factor.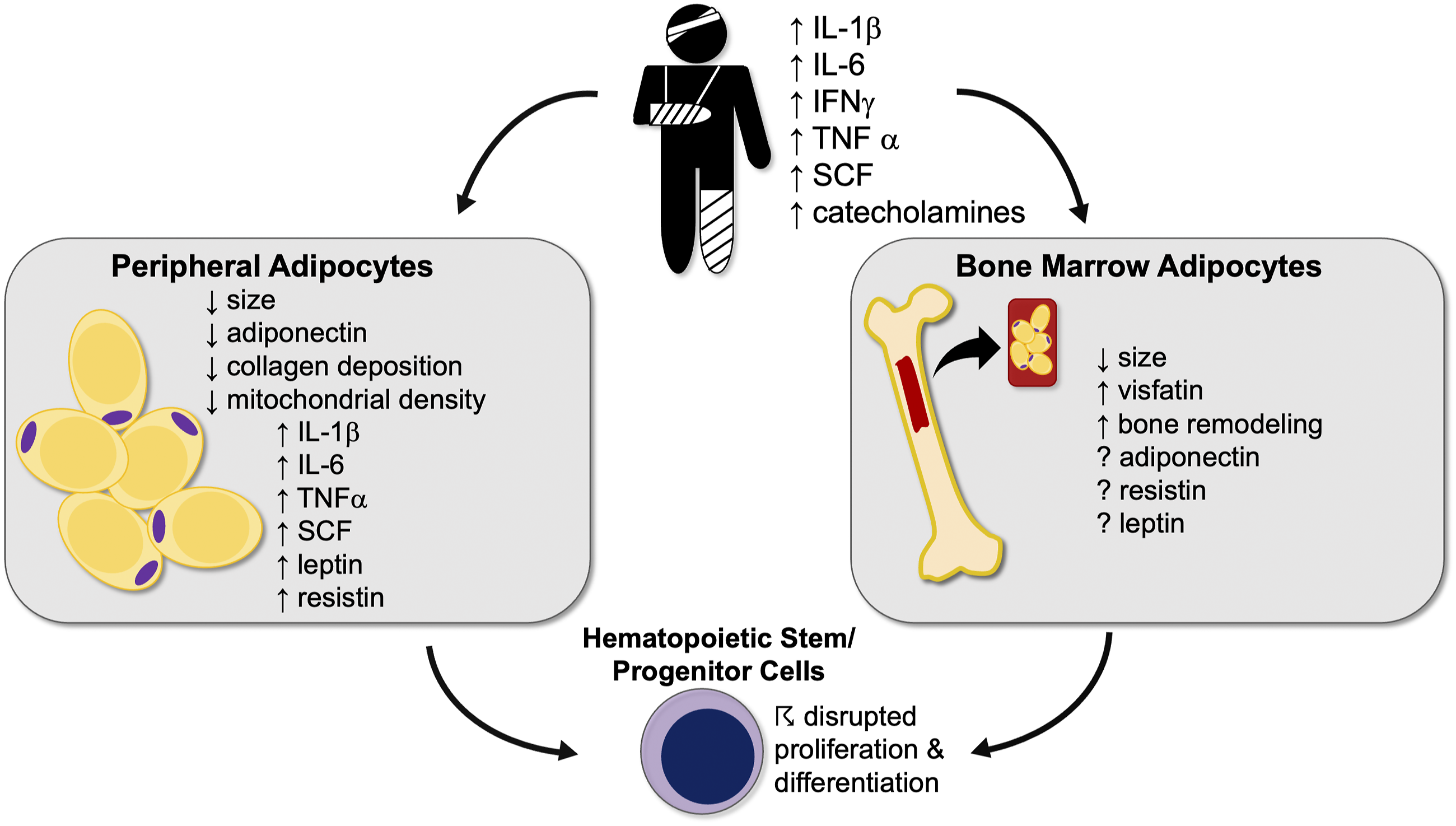
The reduction in erythroid progenitor growth in the trauma cohort is consistent with prior animal and human studies.9,18 This study identified correlations between adipokine transcription and erythroid progenitor growth, particularly CFU-E. Boyd et al 19 demonstrated the dependence of both erythropoiesis and myelopoiesis on the bone marrow adipocyte niche. Bone marrow niche adipokines may exhibit a unique local effect that is distinct from their systemic effects, and that this local influence is not completely understood.
Using elective THR patients as our operative control group inherently consists of older, female-predominant patients as compared to our younger male trauma patients. Our subgroup analysis revealed that resistin and leptin exhibited female-specific differences between trauma patients and THR. Among those over age 65, leptin differences were more prominent while differences in adiponectin were blunted. Aging is associated with a decline in plasma leptin and adiponectin and this is particularly notable in females.20 This may amplify the trauma-induced increase in leptin and dampen the reduction in adiponectin after trauma. Furthermore, resistin levels are generally higher among females, which may prime females for a more pronounced increase after injury.
One murine study found that adiponectin levels drop in hemorrhagic shock and can be partially restored with fresh frozen plasma (FFP) transfusions, and that some of FFP’s protective effects are mediated by adiponectin.21 In our study, hemorrhagic shock was an inclusion criterion for the trauma cohort, and most received transfusions. Following trauma, plasma levels of resistin and leptin both weakly correlated with longer ICU length of stay, and plasma resistin correlated with development of complications, but there were no correlations identified between resistin, leptin, adiponectin, or visfatin and packed red blood cell administration or injury severity.
This study has several limitations. Correlating decreased CFU-E growth with reticulocyte counts would have allowed for better assessment of terminal erythropoiesis. It is possible that the variability in anatomic aspirate site might be associated with differences in volume of bone marrow adipocytes at each site. Similarly, there may be considerable heterogeneity in adipokine release and temporal changes may be more significant than a single timepoint.
Adipokines are differentially released into the circulation after trauma, with some degree of variability depending on age and sex. Additionally, their adipokine expression in the bone marrow may play a role in erythroid progenitor growth suppression, particularly CFU-E, after injury. Further investigation of bone marrow adipose tissue and the impact of local adipokines in the bone marrow microenvironment are needed to define their precise hematopoietic impact following severe injury.
Footnotes
Acknowledgments
The authors would like to thank Jillianne Brakenridge, Ruth Davis, Ashley McCray, Jennifer Lanz, and Iverson Williams for their clinical research support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health (NIGMS R01 GM105893, R01 GM113945, T32 GM-008721).