
Abstract
Objectives
Laryngopharyngeal reflux (LPR) is associated with inflammatory and neoplastic airway diseases. Gastric pepsin internalized by airway epithelial cells during reflux contributes to oxidative stress, inflammation, and carcinogenesis. Several plant extracts and compounds inhibit digestive enzymes and inflammatory or neoplastic changes to the esophagus in models of gastroesophageal reflux. This study examined the potential of chemoprotective phytochemicals to inhibit peptic activity and mitigate pepsin-mediated damage of airway epithelial cells.
Methods
Cultured human laryngeal and hypopharyngeal epithelial cells were pretreated with curcumin (10 μmol/L), ecabet sodium (125 μg/mL), and anthocyanin-enriched black-raspberry extract (100 μg/mL) 30 minutes before treatment with pepsin (0.1 mg/mL; 1 hour; pH 7). Controls were treated with media pH 7 or pepsin pH 7 without phytochemicals. Cell damage and proliferative changes were assessed by electron microscopy, cell count, thymidine analog incorporation, and real-time polymerase chain reaction array. Pepsin inhibition was determined by in vitro kinetic assay.
Results
Micromolar concentrations of curcumin, ecabet sodium, and black-raspberry extract inhibited peptic activity and pepsin-induced mitochondrial damage and hyperproliferation. Curcumin abrogated pepsin-mediated depression of tumor suppressor gene expression and altered the subcellular localization of pepsin following endocytosis.
Conclusions
Several phytochemicals inhibit the pepsin-mediated cell damage underlying inflammatory or neoplastic manifestations of LPR. Dietary supplementation or adjunctive therapy with phytochemicals may represent novel preventive or therapeutic strategies for LPR-attributed disease.
Introduction
Laryngopharyngeal reflux (LPR), the backflow of gastric contents into the laryngopharynx, is associated with a diverse spectrum of clinical manifestations, including laryngitis, chronic rhinosinusitis, and otitis media, as well as life-threatening conditions such as sleep apnea and laryngospasm. A high incidence of LPR has been observed in patients with laryngeal cancer, 1 which is a cancer that has a significant impact on voice, swallowing, and quality of life, a low survival rate (40% 5-year survival), and a rising incidence. A case-control study of 96 laryngeal cancer patients and 192 controls determined that reflux increases the risk of laryngeal cancer by twofold. 2 Analyses of neoplastic development in patients with prior gastrectomy suggest that nonacid reflux promotes laryngeal carcinogenesis, as well. 3
The digestive enzyme pepsin is one of the primary noxious elements of refluxate. Studies performed on human laryngeal biopsies and cultured hypopharyngeal and laryngeal cells show that pepsin in a nonacid environment (eg, nonacid reflux) is taken up by the cell via receptor-mediated endocytosis and retained in intracellular vesicles of acidic pH. 4 Deleterious cell and molecular biological changes were detected in airway epithelial cells after pepsin exposure,4–8 including mitochondrial damage, induction of a proinflammatory cytokine expression profile similar to that of gastroesophageal reflux disease, reduced expression of protective mucosal barrier genes or proteins and tumor suppressors, mis-expression of microRNAs associated with head and neck cancer, hyperproliferation, resistance to apoptosis, and anchorage-independent cell growth (unpublished observation). Pepsin increased the tumor volume in a hamster buccal pouch model of mucosal carcinogenesis, 9 and analyses of clinical specimens confirmed that pepsin was present in laryngeal biopsy specimens obtained from patients with laryngeal cancer and was absent in those from patients without. 4 These findings corroborate the clinical association of weak or nonacid LPR with carcinogenesis in the airways and implicate pepsin in the disease process.
In accordance with evidence that LPR disease is primarily attributed to pepsin rather than acid, proton-pump inhibitors exhibit minimal efficacy in resolution of LPR signs and symptoms. 10 Although antireflux surgery has been shown to resolve symptoms in up to 85% of LPR patients, a less invasive treatment modality has yet to be proven. Several dietary phytochemicals and fruit and vegetable extracts have recently been identified that directly inhibit peptic activity and attenuate inflammation, lesions, and carcinogenesis in animal models of gastroesophageal reflux. Such compounds hold promise as a clinical option for reflux patients.
Curcumin, a yellow polyphenol present in the household spice turmeric (Curcuma longa), has a long history of use in traditional medicine as a potent anti-inflammatory. Recent research has demonstrated its direct interaction with as many as 33 human inflammatory signaling molecules, its modulation of expression of an extensive list of markers of cell proliferation and apoptosis, and its inhibition of tumor development in animal models of carcinogenesis of the oral cavity, skin, forestomach, duodenum, and colon.11–13 In a phase II clinical trial, curcumin (600 mg 5 times per day) resolved peptic ulcer in 48% of subjects in 4 weeks and 78% of subjects in 12 weeks. 14 A pilot study of patients with Barrett's esophagus, the premalignant intestinal metaplasia of esophageal epithelium attributed to reflux, demonstrated that 500 mg curcumin supplementation per day for just 7 days had antineoplastic effects in Barrett's tissues (increased apoptosis and potentially reduced activity of NF-χB [nuclear factor kappa B]). 15
Anthocyanins are potent antioxidants that confer bright red, blue, and purple colors to foods such as berries, grapes, apples, purple cabbage, and corn. They are among the most abundant polyphenols in the human diet, consumed at levels tenfold higher than those of other common dietary phenols. Anthocyanins alter the expression of genes and proteins that regulate inflammation, proliferation, angiogenesis, and cell migration and inhibit the development of esophageal and colon cancers in animal models of disease.16,17 An anthocyanin-enriched freeze-dried extract of black-raspberry powder (BRB; 4% to 5% anthocyanin by dry weight) decreased rectal polyp numbers in patients with familial adenomatous polyposis and reduced markers of inflammation, angiogenesis, oxidative stress, and/or neoplasia in patients with oral dysplasia and Barrett's esophagus.18–20
Ecabet sodium (ES), a phenolic dehydroabietic acid derivative of pine resin, is a well-researched pepsin-inhibitory and mucoprotective topical agent used widely for the treatment of gastritis and gastric ulcer. It precipitates and inactivates pepsin, exhibits high affinity for the mucosal epithelium, stimulates re-epithelialization after mucosal damage, and has been shown to inhibit mucosal dysplasia in experimental animals.21–23 As demonstrated in a rat model of gastroesophageal reflux disease, inactivation of pepsin by ES provides complete protection against esophageal lesions despite continued exposure of esophageal epithelium to gastric acid. 24
In spite of the poor efficacy of acid-targeting therapeutics for LPR disease and evidence that pepsin (independent of the pH of the refluxate) is the primary deleterious agent of refluxate during LPR, proton pump inhibitors continue to be the most frequently prescribed therapeutic for LPR. No medical therapies have yet been designed to specifically or directly target pepsin. Dietary phytochemicals capable of inhibiting pepsin and/or its inflammatory and neoplastic effects have the potential to alleviate a myriad of symptoms and conditions attributed to LPR. The aim of the current study was to investigate the potential of curcumin, BRB, and ES to inhibit pepsin and protect against pepsin-mediated damage of laryngeal and hypopharyngeal epithelial cells via in vitro kinetic assay, electron microscopy, cell count assay, thymidine analog incorporation, and real-time polymerase chain reaction (PCR) array.
Methods
Kinetic Assay
Synthetic pepsin substrate containing a paranitrophenylalanine chromophore at the site of pepsin cleavage (Lys-Pro-Ala-Glu-Phe-PNP-Arg-Leu-COOH; synthesized by Mark Lively, Wake Forest University) was reconstituted to 20 mmol/L in methanol, and curcumin (Sigma-Aldrich Corp, St Louis, Missouri) to 50 mmol/L in 0.5N sodium hydroxide. Anthocyanin-enriched BRB was prepared as previously described. 16 Polyphenols were assayed at, above, and below effective concentrations cited in the literature (10 μmol/L curcumin, 100 μg/mL BRB, and 125 μg/mL ES). In triplicate reactions, 1 μg/mL pepsin 3b (purified from human gastric juice; Medical College of Wisconsin Institutional Review Board protocol No. PR4759) or 1μg/ mL pH-inactivated pepsin 3b control (inactivated at pH 8.5; 37°C; 15 minutes) was assayed in 0 to 1 mg/mL bovine serum albumin (negative control), 0 to 80 μmol/L curcumin, 0 to 0.4 mg/mL BRB, or 0 to 1 mg/mL ES (Molcan Corp, Toronto, Canada) plus 70 μmol/L synthetic pepsin substrate in 0.1 mol/L glycine pH 3.0. Absorbance at 300 nm was recorded every 12 seconds for 20 minutes. The mean initial linear hydrolysis rate (±SD; n = 3) was plotted over time, and the significance of change in the rate of hydrolysis was determined by Student's t-test (2-sample, paired t-test for means).
Cell Culture
Human hypopharyngeal epithelial FaDu cells (ATCC, Manassas, Virginia) were grown in Eagle's minimum essential medium supplemented with 10% fetal bovine serum (ATCC) and 1x Antibiotic-Antimycotic (Ab-Am; Life Technologies, Grand Island, New York). Gastric adenocarcinoma AGS cells (ATCC) were cultured in F-12K medium (ATCC) supplemented with 10% fetal bovine serum and 1x Ab-Am. A primary human laryngeal epithelial cell culture from the posterior cricoid cartilage of control subjects who had no reflux, inflammatory disease, or neoplasia and were undergoing surgery for structural or functional reasons (Medical College of Wisconsin Institutional Review Board protocol No. PRO0000-4777) was established and characterized as described previously. 8
Normal primary laryngeal and hypopharyngeal FaDu cells were grown to 30% confluence, pretreated with or without curcumin (10 μmol/L), ES (125 μg/mL), or BRB (100 μg/mL) for 30 minutes, rinsed with phosphate-buffered saline solution (PBS), and treated with porcine pepsin (0.1 mg/mL; Sigma-Aldrich) pH 7.0 for 1 hour at 37°C in 5% carbon dioxide; control cells were treated with media pH 7.0 for 1 hour at 37°C in 5% carbon dioxide. The cells were rinsed with PBS and cultured in normal growth media for 24 hours (approximately 65% final confluence). To determine whether phytochemicals affected pepsin-induced hyperproliferation, we harvested cells by trypsinization and counted them or exposed them to a thymidine analog and processed them for flow cytometry (Click-iT EdU Flow Cytometry Assay; Life Technologies); the statistical significance of a change in cell number or percentage of cells in S-phase was determined by t-test (2-sample, paired t-test for means). To determine whether the phytochemicals affected mitochondrial damage by pepsin, we processed cells for routine transmission electron microscopy. To measure the change in cell cycle regulatory gene expression, we harvested curcumin- and/or pepsin-treated cells and controls and subjected them to real-time PCR analysis. To investigate the effect of curcumin on receptor-mediated endocytosis of pepsin by immunoelectron microscopy, we pretreated FaDu cells at 70% confluence with or without curcumin (10 μmol/L in culture medium; 30 minutes; 37°C; 5% cardon dioxide), washed them briefly in PBS, and then held them at 4°C for 30 minutes to inhibit endocytosis. The cells were then incubated with or without human pepsin 3b (200 ng/mL; 20 minutes) at either 37°C or 4°C. The cells were washed in cold PBS and fixed in 0.1% glutaraldehyde containing 4% paraformaldehyde in 100 mmol/L Sörensen's buffer pH 7.4 for 1 hour on ice.
Transmission Electron Microscopy
After fixation in 2.5% glutaraldehyde (100 mmol/L sodium cacodylate buffer pH 7.2), FaDu and AGS cells were post-fixed in 1% osmium tetroxide on ice for 1 hour and then dehydrated in a graded methanol series and embedded in EPON 812 (Electron Microscopy Sciences, Hatfield, Pennsylvania). We cut 60-nm sections, stained them with uranyl acetate and Reynolds lead citrate, and examined them in a Japanese Electron Optics Ltd (JEOL) 2100 Transmission Electron Microscope (JEOL USA Ltd, Peabody, Massachusetts).
Human Cancer PathwayFinder Super Array
RNA from 3 biological replicates was isolated with the SuperArray RT 2 -qPCR-Grade RNA Isolation Kit (Qiagen, Valencia, California). RNA was reverse-transcribed with the SuperArray RT 2 First Strand Kit (Qiagen). Complementary DNA and RT 2 SYBR Green/Fluorescein qPCR Master Mix (Qiagen) were applied to the Human Cancer PathwayFinder PCR Array (Qiagen). Real-time PCR was performed in the iCycleriQ Multicolor Real Time PCR Detection System (Bio-Rad Life Sciences, Hercules, California) under the reaction conditions specified by the array manufacturer. Reactions exhibiting more than 1 peak within the melt curve or a threshold cycle (Ct) greater than 35 were excluded from analysis. The means of housekeeping genes for each replicate were subtracted from each gene value to normalize the data. Differential expression was evaluated by fitting a mixed model to the entire data set with fixed gene and gene-treatment interaction effects, and random intercept and treatment effects for each biological replicate. We adjusted the p values for the gene-specific treatment effects using the Benjamin-Hochberg procedure with a 5% cutoff. The synergistic or antagonistic effects of pepsin and curcumin were identified by comparing log (base 2) expression ratios from the pepsin-treated condition to those of the pepsin-and-curcumin–treated condition (both normalized to the untreated condition). Statistical analysis of the divergence between the actual and expected (additive) effects was performed as for differential expression data.
Immunoelectron Microscopy
After fixation, cells were washed (3 × 5 minutes) in Sörensen's buffer, scraped from the culture dishes, and gently pelleted at 1,000g for 10 minutes. Cell pellets and tissue specimens were processed into Lowicryl K4M resin by the “progressively lower temperature” method of Berryman and Rodewald 25 following the protocol of Burleigh et al. 26 After polymerization, ultrathin (60-nm) sections were cut and mounted on Formvar and carbon–coated copper grids, and immunoelectron microscopy was performed on the sections by incubating them sequentially on 25-μL droplets in a humidified chamber following the protocol of Burleigh et al. 26 Pepsin was detected with a mouse monoclonal anti-human pepsin antibody (Promab, Richmond, California) synthesized against a previously described peptide. 5 Mouse anti-human pepsin was revealed with goat anti-mouse 10-nm conjugated gold colloidal reagent (Electron Microscopy Sciences). The sections were examined with a JEOL 2100 transmission electron microscope.
Results
Kinetic Assay
Curcumin (at least 25 μmol/L, or 9.2 μg/mL), ES (at least 100 μg/mL), and BRB (at least 50 μg/mL) inhibited the initial or log-phase rate of substrate hydrolysis by pepsin (p < 0.05; Fig 1) and reduced the percentage of substrate hydrolyzed at the end of assay (p < 0.05; Table 1). Bovine serum albumin did not significantly inhibit the rate of hydrolysis until at least 1 mg/mL. The end-point percentage of substrate hydrolyzed was reduced from 12% (without phytochemical) to less than 4% by 80 μmol/L (29.5 μg/mL) curcumin, 200 μg/mL BRB, and 100 μg/mL ES. Ecabet sodium achieved near-complete inhibition of hydrolysis at concentrations of 500 μg/mL or more (0.02% ± 1.07%; p = 0.0017). The end-point percentage of substrate hydrolyzed was not significantly reduced by 1 mg/mL bovine serum albumin (t20 = 11.07% ± 1.12%; p = 0.0628) or pH-inactivated pepsin (t20 = 0.15% ±0.91%; p = 0.0040; data not shown).
End-Point Percentage of Substrate Hydrolyzed by Pepsin in Presence of Polyphenols
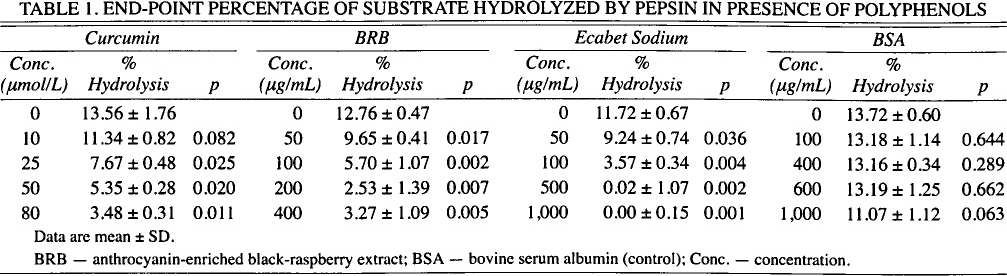
Data are mean ± SD.
BRB — anthrocyanin-enriched black-raspberry extract; BSA — bovine serum albumin (control); Conc. — concentration.
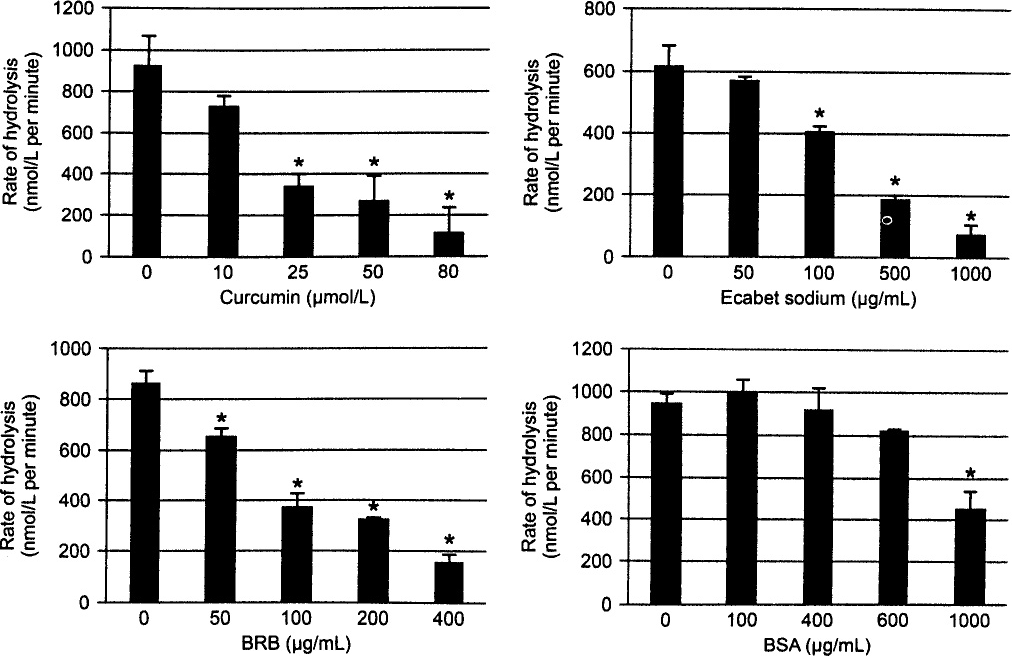
Concentration-dependent inhibition of initial rate of pepsin substrate hydrolysis by curcumin, anthocyanin-enriched black-raspberry extract (BRB), and ecabet sodium (ES) in vitro. Initial (log-phase) rate of hydrolysis of synthetic peptide substrate (70 μmol/L) by pepsin (0.1 μg/mL; 0.1 mol/L; glycine pH 3.0) was determined for increasing concentrations of curcumin, ES, BRB, and bovine serum albumin (BSA) control. Asterisk indicates statistically significant change in rate of hydrolysis relative to rate of hydrolysis without curcumin, BRB, or ES (p < 0.05).
Transmission Electron Microscopy
Pepsin treatment of human hypopharyngeal cells induced mitochondrial damage as described previously 10 (ie, swollen mitochondria with degraded cristae); the control cells (medium pH 7; no pepsin) were unaffected (Fig 2). Curcumin, BRB, or ES treatment prior to pepsin exposure inhibited these mitochondrial changes. Pepsin treatment of gastric cells yielded no detectable damage (Fig 2).
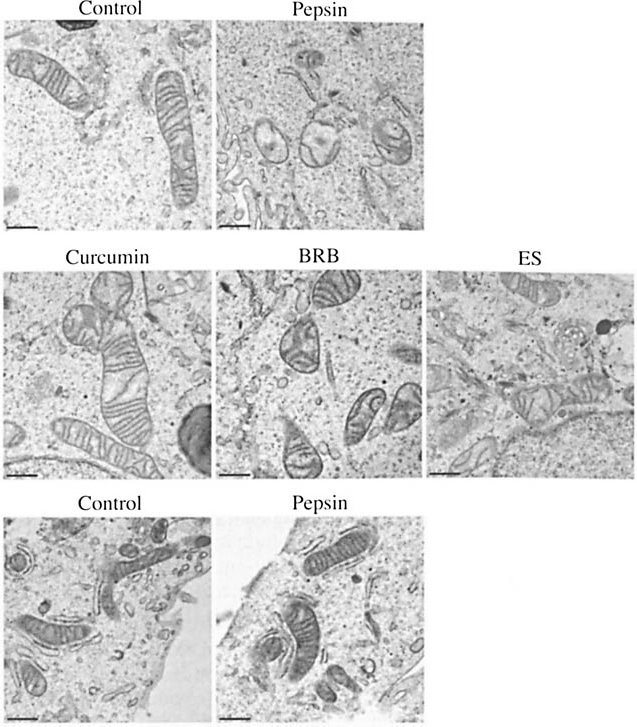
Curcumin, BRB, and ES pretreatment prevented pepsin-induced mitochondrial damage in hypopharyngeal cells, whereas pepsin had no effect on gastric cell mitochondria in vitro. Mitochondrial damage was observed in pepsin-treated hypopharyngeal cells, but not in hypopharyngeal cells treated with media pH 7 alone (top row); mitochondria in pepsin-treated cells (0.1 mg/mL; pH 7; 37°C; 1 hour) appear swollen with degraded cristae relative to those of control cells (media pH 7 alone; 37°C; 1 hour). Pretreatment with 10 μmol/L curcumin, 100 μg/mL BRB, or 100 μg/mL ES (pH 7.4; 37°C; 30 minutes) prior to pepsin treatment (0.1 mg/mL; pH 7; 37°C; 1 hour) prevented change in mitochondria (middle row). Human gastric cell mitochondria (bottom row) were unaffected by pepsin (0.1 mg/mL; pH 7; 37°C; 1 hour) relative to control (pH 7; 37°C; 1 hour). Bars - 0.5 μm.
Cell Proliferation Assays
Pepsin treatment significantly increased the number of human hypopharyngeal and laryngeal, but not gastric cells counted relative to control (p < 0.05; Fig 3A). Pretreatment of hypopharyngeal and laryngeal cells with curcumin, BRB, or ES prior to pepsin exposure significantly reduced the number of cells counted relative to pepsin-treated conditions. Pepsin treatment significantly increased the percentage of human hypopharyngeal cells in S-phase (p < 0.05; Fig 3B); curcumin (10 μmol/L) pretreatment prior to pepsin treatment significantly reduced the percentage of cells in S-phase relative to pepsin treatment alone.
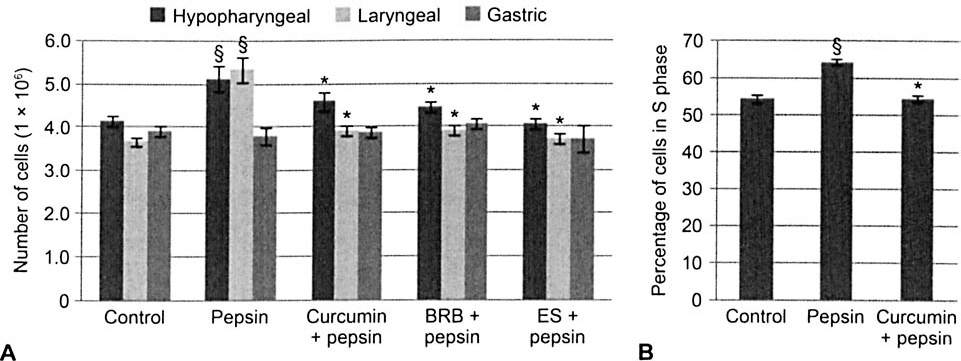
Pepsin increased proliferation of upper airway, but not gastric, cells in vitro, and curcumin, BRB, and ES abrogated pepsin-induced cell proliferation. A) Pepsin treatment (0.1 mg/mL; pH 7; 37°C; 1 hour) increased number of hypopharyngeal FaDu and laryngeal cells counted 24 hours after treatment. Pretreatment (pH 7.4; 37°C; 30 minutes) with curcumin (10 μmol/L), BRB (100 μg/mL), or ES (125 μg/mL) prior to pepsin treatment significantly reduced cell number relative to pepsin-treated condition. Gastric cell number was not significantly affected by pepsin treatment or phytochemical pretreatment followed by pepsin treatment. B) Measurement of DNA synthesis by detection of incorporated thymidine analog confirmed that curcumin pretreatment (10 μmol/L; 30 minutes) abrogated pepsin-induced hyperproliferation. Section sign indicates statistically significant change in cell count relative to control (p < 0.05); asterisk denotes statistically significant change in cell count relative to pepsin-treated condition (p < 0.05).
Human Cancer PathwayFinder Super Array
Relative to untreated controls, pepsin significantly reduced the expression of 28 of 84 assayed genes and increased the expression of 3 genes (more than 1.5-fold; p < 0.05; Table 2). Pretreatment with curcumin prior to pepsin treatment reversed the pepsin-mediated effect on 26 of the 28 genes whose expression was reduced by pepsin, restoring expression of all genes but 1 to normal or untreated levels of expression or higher, and similarly reversed the pepsin-me-diated effect on 2 of the 3 genes whose expression was increased by pepsin. Comparison of log (base 2) ratios indicate statistically significant antagonism between pepsin and curcumin in modulation of the 9 genes most significantly dysregulated by pepsin.
Effect of Curcumin on Pepsin-Mediated Gene Expression Change in Hypopharyngeal Cells
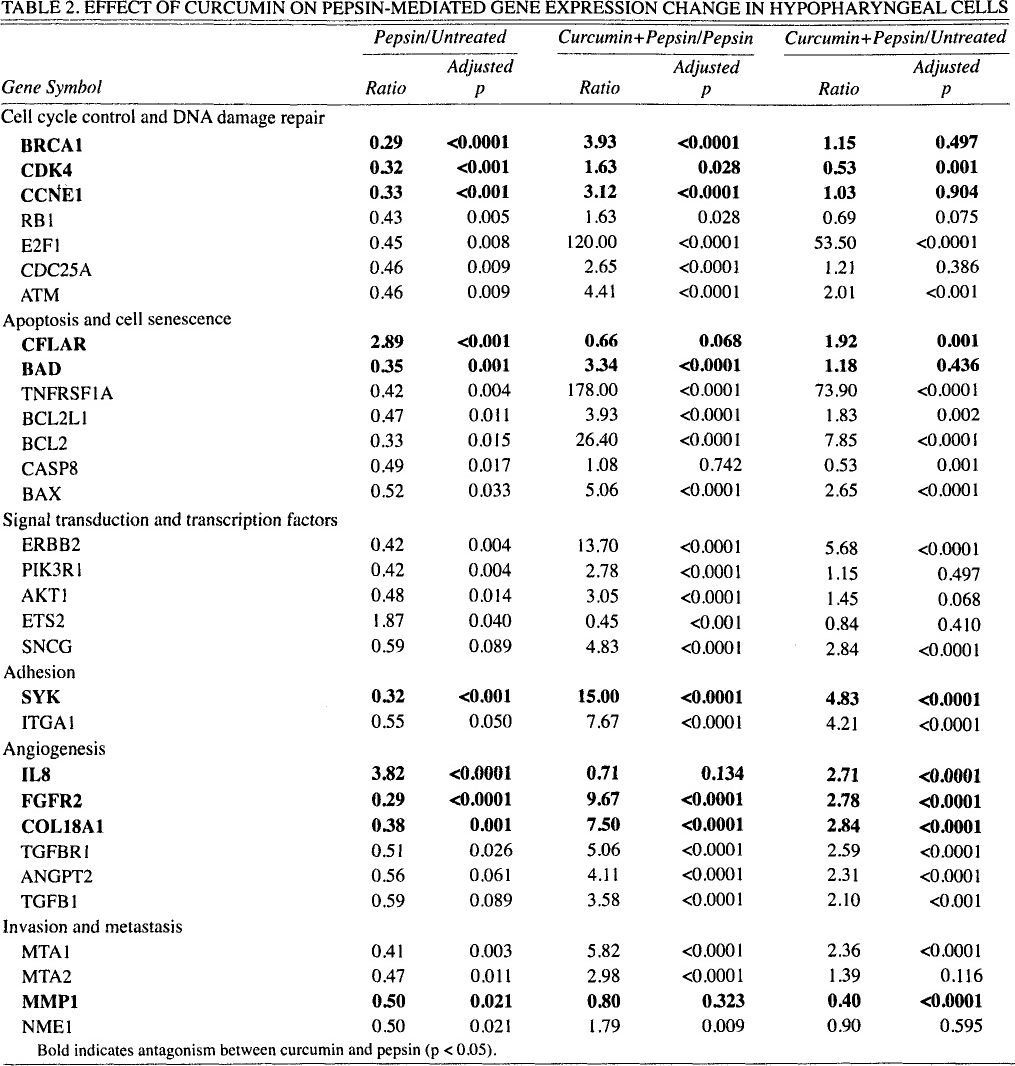
Bold indicates antagonism between curcumin and pepsin (p < 0.05).
Immunoelectron Microscopy
As described previously, 4 the pepsin in hypopharyngeal FaDu cells following incubation at 37°C was primarily restricted to intracellular vesicles (Fig 4); the pepsin remained at the surface of control cells exposed to pepsin at 4°C. In cells pretreated with curcumin, pepsin localization was not restricted to intracellular vesicles, but was frequently scattered throughout the cytosol. Pepsin labeling within curcumin-pretreated cells appeared to be reduced relative to that in cells that received no pretreatment; however, the difference was not directly measured.
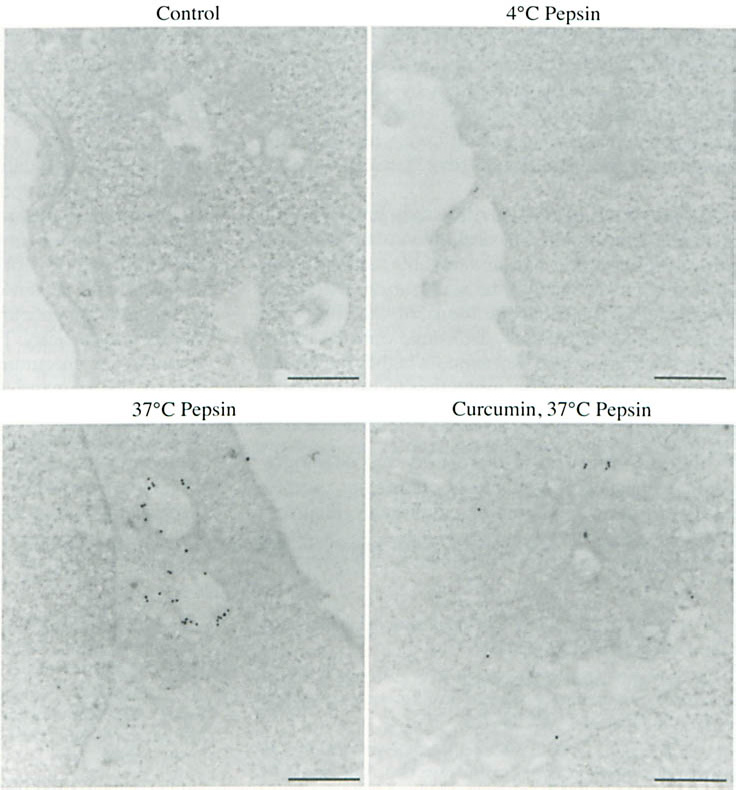
Curcumin altered subcellular localization of pepsin following endocytosis in hypopharyngeal cells. Hypopharyngeal cells were treated with and without curcumin (10 μmol/L; 30 minutes) and human pepsin 3b (200 ng/mL; pH 7.0; 20 minutes) at 4°C and 37°C and subjected to pepsin immunolabeling and transmission electron microscopy. Cells exposed to medium pH 7.0 alone (control) demonstrated absence of staining. Pepsin remained at surface of hypopharyngeal cells after incubation at 4°C. Upon incubation at 37°C, pepsin was endocytosed and retained in intracellular vesicles. In curcumin-pretreated cells, pepsin was found primarily in cytosol. Bars - 0.5 μm.
Discussion
According to the World Health Organization, approximately 90% of deaths in developed countries are attributed to noncommunicable chronic diseases caused, to a large extent, by poor diet, sedentary lifestyle, and tobacco use. Multiple case-control studies have demonstrated an inverse correlation between a fruit- and vegetable-rich diet and cancers of the oral cavity and pharynx,27–29 and aside from smoking, fruit intake is the strongest predictor of survival of head and neck squamous cell carcinoma. 30 In addition to chemoprotective benefits, a fruit-and vegetable-rich diet may resolve noncancerous manifestations of LPR, as well; a diet that is high in whole grains and low-acid fruit and vegetables and low in fat, red meat, and processed and acidic foods was shown to reduce signs and symptoms of LPR that prior proton pump inhibitor therapy had failed to mitigate. 31 Whole animal model and clinical research indicates that the anti-inflammatory and anti-neoplastic activity of dietary phytochemicals is most significant in tissues with which they come into direct contact, so cancers of the aerodigestive tract are a prime target for their use. Dietary chemopreventive and therapeutic compounds exhibit low toxicity, selective uptake and activity in tumors or rapidly proliferating cells, and simultaneous targeting of multiple pathways dysregulated during inflammatory disease and carcinogenesis without the increased potential for side effects that accompanies combined use of single-target drugs.
The pepsin inhibitory potential of anthocyanin and curcumin has not been described in the literature to date. As described herein, curcumin and an anthocyanin-enriched BRB extract inhibited peptic activity at concentrations lower than that of the known pepsin inhibitor and ulcer therapeutic ES. All 3 compounds or extracts protected laryngeal and hypopharyngeal cells against the mitochondrial damage and hyperproliferation caused by pepsin. Analysis of the expression of 84 genes representative of the 6 biological pathways involved in transformation and tumorigenesis revealed that 90% of changes induced by pepsin are abrogated by curcumin pretreatment. A tool for future studies, a suspected pepsin-tolerant cell population, was confirmed as well; human gastric cells displayed insensitivity to pepsin, as indicated by a normal mitochondrial morphology and proliferation rate following treatment with pepsin.
Enzymatic inhibition may have contributed to the observed protective effects of BRB and ES; concentrations that inhibited mitochondrial damage and hyperproliferation also significantly reduced peptic activity (approximately 55% and more than 70%, respectively). The concentration of curcumin that inhibited mitochondrial damage and hyperproliferation in these experiments did not significantly alter peptic activity. Curcumin is a hydrophobic molecule that avidly accumulates in biological membranes; although our data suggest that curcumin-mediated enzyme inhibition is unlikely to have contributed to protection against cell damage by pepsin, we cannot exclude the potential accumulation of curcumin at subcellular locations (eg, cell membrane, endocytic vesicles) in which it might achieve a concentration high enough to inhibit pepsin. Future examination of the capacity of these compounds to protect against pepsin-mediated hyperproliferation over a range of concentrations, their phytochemical-enzyme binding kinetics, and the reversibility of their inhibition of pepsin will provide more information about the role of enzymatic inhibition in their disease-preventive capacities.
Curcumin and anthocyanin are known inhibitors of NF-χB, a central component of signaling pathways that link inflammation and cancer, and many of the anti-inflammatory and chemoprotective effects of these molecules have been found to correlate with changes in the expression of genes regulated by NF-χB. The finding that several NF-χB–regulated genes (eg, matrix metalloproteases, bax, caspases, and bcl-2) are dysregulated upon exposure to pepsin and that curcumin-mediated reversal or reduction of these aberrances coincides with protection against cell damage implicates NF-χB in the airway epithelial cell damage caused by pepsin. Taken together with the established role of NF-χB in gastroesophageal reflux disease and our recent unpublished observation that pepsin induces oxidative stress, known to activate NF-χB, in airway cells, these data encourage further exploration of the role of NF-χB in pepsin-mediated cell damage and the potentiating effect of pretreatment with curcumin. In future studies we hope to better define the contribution of NF-χB to pepsin-mediated damage through the use of an NF-χB reporter assay and an NF-χB super-repressor combined with functional assays.
Given its interaction with the cell membrane and its capacity to alter lipid bilayer properties, transmembrane protein lifetimes, and endosomal trafficking,32–33 we further suspected that curcumin might protect cells against pepsin by altering pepsin uptake. Indeed, pepsin endocytosis appeared to be reduced in curcumin-pretreated cells; however, further experiments will be required to measure the significance of this change. Of greatest interest, curcumin altered the subcellular localization of endocytosed pepsin such that it was no longer restricted to intracellular vesicles, but was scattered throughout the cytosol. Intracellular reactivation of nonacid pepsin is necessary for its downstream deleterious effects; this reactivation is evidenced to occur within intracellular vesicles of low pH in which pepsin is normally retained upon receptor-mediated endocytosis. 4 Perturbing the intracellular localization of pepsin to acidic vesicles would presumably inhibit its reactivation and subsequent damage to airway epithelium. These findings suggest a novel mechanism of LPR disease prevention by curcumin. We plan to further characterize the effect of curcumin on pepsin endocytosis and trafficking, including the duration and reversibility of this effect.
Although the current in vitro study proved useful in demonstrating the cell and molecular biological effects of curcumin and anthocyanin, animal studies and clinical trials will be necessary to determine whether effective concentrations of the drugs are achievable at target sites, determine optimal routes of administration, and determine whether the effects observed confer disease protection to the individual.
Multitargeting anti-inflammatory and chemopreventive plant compounds hold significant promise as therapeutics for complex, multigenic disorders such as chronic inflammatory diseases and cancer. This study is the first to assess the effect of dietary phytochemicals in the context of LPR. Our findings include two novel mechanisms of curcumin-mediated chemoprotection that have not been described in the literature to date and are particular to the context of reflux — specifically, inhibition of peptic activity and altered pepsin uptake or trafficking. The capacity of curcumin to mitigate the carcinogenic effects of pepsin through modulation of gene expression and inhibition of peptic activity by either direct interaction with pepsin or disruption of its intracellular reactivation encourages further exploration of its therapeutic capacity. This study indicates significant potential for curcumin and anthocyanin in prevention of cell damage and neoplasia due to nonacid pepsin and supports the use of dietary modification or supplementation with anti-inflammatory or chemopreventive phytochemicals as a novel pepsin-protective preventive strategy for LPR-attributed diseases, including laryngeal cancer.
Footnotes
Acknowledgment
We acknowledge Aniko Szabo, PhD, Department of Biostatistics, Medical College of Wisconsin, for assistance with statistical analysis.