
Abstract
Although callous-unemotional (CU) traits have been associated with amygdala hypoactivity, it is unclear whether such traits are associated with amygdala structure. This study examines the relationship between amygdala structure and CU traits and considers sex differences in hemisphere-related asymmetries in this relationship. In this brain imaging study of children (n = 272), a significant sex × hemisphere × CU traits interaction was observed. Females with high CU traits and males with low CU exhibited a rightward exaggerated amygdala volume asymmetry, whereas females with low CU traits and males with high CU traits did not. Findings may reflect sex-related influences related to CU traits and amygdala volume and are broadly consistent with a neurodevelopment perspective on psychopathic-like traits in children.
Introduction
Callous-unemotional (CU) traits encompass characteristics in children that reflect interpersonal and emotional insensitivity, including low empathy, shallow affect, and a lack of moral emotions such as guilt or remorse (Frick & White, 2008; Frick et al., 2003). In fact, a recent meta-analysis found that CU traits are strongly negatively correlated with empathy, prosociality, and guilt (Waller et al., 2019). Lending credence to the importance of understanding CU traits, CU traits were added as a specifier for the diagnosis of conduct disorder in the Diagnostic and Statistical Manual of Mental Disorders (5th ed.; DSM-5; American Psychiatric Association, 2013). CU traits in children have been conceptualized as a precursor to the affective features of adult psychopathy (Barry et al., 2000; Waller et al., 2019), and children who exhibit CU traits typically engage in more severe and chronic antisocial behaviors, and with an earlier onset, than their counterparts without CU traits (Bolhuis et al., 2019; Frick et al., 2003, 2014). The presence of CU traits has been useful in identifying a subgroup of antisocial individuals who show distinct cognitive, emotional, temperament, and personality characteristics compared with those who do not exhibit these traits (Frick et al., 2014). Developmentally, CU traits can be observed in in preschool children as young as 2 years old and appear to be relatively stable (Frick et al., 2014; Waller et al., 2012; Waller & Hyde, 2018).
A key neural structure that has been examined in relation to CU traits is the amygdala. This brain region, considered the “emotional center” of the brain, continues to develop throughout childhood and adolescence (Kaczkurkin et al., 2016; Tottenham & Galván, 2016). Some of the change in amygdala volume corresponds to pubertal development (Goddings et al., 2014) and observed sex differences in puberty-related maturation of the amygdala have been thought to contribute to sex-related social and cognitive differences during adolescent development and into adulthood (Bramen et al., 2011; Satterthwaite et al., 2014). Although there has been robust evidence that CU traits are associated with aberrant amygdala functioning (Schwenck et al., 2017; Sebastian et al., 2014; Szabó et al., 2017; Viding et al., 2012), there is a dearth of research on whether amygdala structure is associated with CU traits.
Of the limited prior studies that have examined amygdala structure in relation to CU traits, most report null findings (Bolhuis et al., 2019; De Brito et al., 2009; (Fairchild et al., 2011; Fairchild, Hagan, et al., 2013; Sebastian et al., 2016; Wallace et al., 2014). Three studies reported bilateral amygdala volume reductions associated with CU traits (Caldwell et al., 2019; Cardinale et al., 2019; Rogers & De Brito, 2016); however, one of these acknowledged that this finding differed based on sex, age, and brain hemisphere (Cardinale et al., 2019). However, no study to date has directly adjusted for pubertal development when examining the relationship between CU traits and amygdala structure, despite utilizing samples that span a wide age range with individuals at different stages of pubertal maturity. For example, Caldwell et al. (2019) found that CU traits were associated with reduced amygdala volume in a sample ranging in age from 13 to 20 years. Cardinale et al. (2019) found that CU traits were associated with bilateral amygdala volume reductions in a sample of 10- to 18-year-olds. A meta-analysis by Rogers and De Brito (2016) found reduced amygdala volume when comparing conduct-disordered youth aged 8 to 21 years to typically developing youths aged 8 to 21 years and noted that this finding was stronger in conduct-disordered youth with CU traits. Although age has often been controlled for in these prior studies, the effects of age and pubertal stages on subcortical brain development may differ (Blakemore et al., 2010; Goddings et al., 2014). Differences in sex hormone receptor concentration in the amygdala may account for different sex-related patterns of amygdala development during puberty, and there is evidence that pubertal development has an independent influence on amygdala volume from that of chronological age (Goddings et al., 2014). Thus, the primary aim of the current study was to determine the relationship between amygdala structure and CU traits while controlling for multiple covariates, including both age and pubertal development.
In addition to examining whether there is a general relationship between CU traits and amygdala volume, there is preliminary evidence that the association between the amygdala and CU traits may vary based on sex and brain hemisphere. In a study by Cardinale et al. (2019), there were sex and brain hemisphere differences observed in the relationship between amygdala volume and CU traits. In particular, they found that although CU traits predicted bilateral amygdala volume in males across all age groups, CU traits predicted only left amygdala volume in younger females. This finding may suggest divergent neurodevelopmental profiles for males and females who exhibit CU traits. The overall findings fit more broadly into an emerging literature suggesting that clinical and antisocial populations exhibit structural hemispheric asymmetries (Raine et al., 1997, 2004).
While prior studies of CU traits have often not explicitly studied asymmetries, Visser et al. (2014) suggest that previous structural findings in one hemisphere but not another may reflect hemispheric asymmetries. Such findings have been reported in various antisocial individuals. For example, Sterzer et al. (2007) reported reduced left, but not right, amygdala volume in male adolescents with conduct disorder compared with age-, sex- and IQ-matched controls. In antisocial adults, left amygdala connectivity with paralimbic regions has been found to be increased in violent offenders and decreased in nonoffender controls, but this pattern was not found for right amygdala connectivity (Siep et al., 2019). Moreover, connectivity between the left amygdala and medial prefrontal cortex decreased in violent offenders compared with nonoffender controls, but this was not seem for right amygdala connectivity (Siep et al., 2019). Raine et al. (1998) reported increased subcortical glucose metabolism for the right, but not left, hemisphere in affective and predatory murderers compared with controls. In normally developing individuals, males exhibit right-greater-than-left (R > L) asymmetry in the amygdala, whereas females show no amygdala asymmetry (Uematsu et al., 2012). Overall, given that there may be asymmetries to consider, a secondary aim is to assess whether sex and hemispheric differences exist in regard to the relationship between amygdala structure and CU traits.
The current study examines the relationship between CU traits and amygdala in a relatively large sample of male and female youth. In addition, given emerging evidence of sex differences and brain asymmetries in association with antisocial individuals, sex- and brain hemisphere-related effects are considered. Importantly, the current study also overcomes a key limitation of prior studies, which consider age-related effects but have often failed to account for the effect of puberty, by utilizing a narrower age range and accounting for pubertal development. Moreover, aggression, impulsivity, and narcissism were controlled for to ensure that results were a function of the level of CU traits rather than severity of other conduct issues.
Method
Participants
Data from 454 children aged 11 to 13 years were collected (M = 11.92, SD = 0.60, 223 males). Recruitment details and inclusion criteria are detailed elsewhere (Choy et al., 2015; Liu et al., 2013; Richmond et al., 2013). The racial composition of the sample was 78.2% African American, 11.9% Caucasian, 4.8% multiracial, 0.9% other, 0.7% Hispanic, 0.4% Asian/Pacific Islander, and 0.4% Native American (the remaining 2.6% did not provide a response). The caregiver of each participant was also recruited as an informant for the child’s behavior. Caregiver participants were primarily the biological mother (87.7%). Written informed consent and assent were obtained from both parents and children. The Institutional Review Board at the University of Pennsylvania and the Philadelphia Department of Health approved the protocols of this study.
CU Traits
CU traits were measured using the Antisocial Process Screening Device (APSD; Frick & Hare, 2001). This 20-item scale comprised three subscales: CU traits (six items), narcissism (seven items), and impulsivity (five items) (two items are not included in the subscales). Items are rated on a 3-point scale, ranging from 0 = not at all true to 2 = definitely true. Similar to prior work (Kahn et al., 2013), child self-reports were combined with parent report scales to create a composite measure of each item based on the highest rating of each symptom (Frick & Hare, 2001; Piacentini et al., 1992). Item scores were summed to create an overall score for each subscale. Similar to previous studies (Martin-Key et al., 2017; Schwenck et al., 2012), individuals were classified as “low CU” or “high CU” based on a median-split of the total composite score.
Amygdala Volume
Magnetic resonance imaging (MRI) acquisition
Brain images were obtained on 313 subjects. Structural MRI (effective voxel resolution of 0.9375 × 0.9375 × 1 mm) was acquired on a Siemens Trio 3 T (Erlangen, Germany) system using a 5-min magnetization-prepared, rapid acquisition gradient echo (MPRAGE) T1-weighted sequence with the following parameters: repetition time (TR) = 1,630 ms, echo time (TE) = 3.87 ms, inversion time (TI) = 1,100 ms, flip angle = 15, field of view (FOV) = 256 × 192.
There were no significant differences in sex, race, age, CU traits, pubertal development stage, narcissism, or aggression between individuals with brain imaging data and those without (ps > 05; see Supplemental Table S1). However, there were higher impulsivity scores for those with brain imaging data (M = 5.45, SD = 1.86) than those without (M = 5.02, SD = 1.83), t(429) = 2.163, p = .031.
Image processing
To obtain volumetric measures, T1-weighted raw data were imported into FreeSurfer version 5.0 (https://surfer.nmr.mgh.harvard.edu/). FreeSurfer’s surface-based image processing pipeline is described extensively elsewhere (Fischl, 2012). Briefly, for each subject, recon-all processing stream was used, which includes motion correction, skull-stripping, registration, segmentation, smoothing, and parcellation mapping. Each voxel in the normalized brain volume was then assigned to one of 40 labels, using a probabilistic atlas obtained from a manually labeled training set (Fischl et al., 2002). The labels used for further analysis in the current study were left and right amygdala. Left and right amygdala volume values were summed to create a total amygdala volume value for each participant. The average Euler, a measure of structural image quality, was also extracted using FreeSurfer and used to determine the level of noise for each participant’s scan. Participants whose average Euler value was 3 SDs or greater were excluded from analyses (n = 9). Among the participants who had brain imaging data, there were no significant differences in sex, race, age, CU traits, pubertal development stage, narcissism, or aggression between individuals who were excluded from analyses based on their Euler value and those who were included. However, those who were excluded from analyses because of their Euler value had more impulsivity (M = 6.67, SD = 1.87) than those who were included (M = 5.41, SD = 1.85), t(305) = 2.007, p = .046.
Covariates
All analyses were adjusted for covariates, including age, race, total intracranial volume, and average Euler. Unlike prior studies that only controlled for chronological age, the current study controls for pubertal development as assessed by Tanner stages (Tanner, 1962). Using 5-point scales, boys were rated for genital development and pubic hair growth, whereas girls were rated for breast development and pubic hair growth (see Supplemental Materials for exploratory robustness analyses regarding pubertal development). To assess the relationship between amygdala volume and CU traits specifically, aggression based on the self-reported reactive-proactive questionnaire (Raine et al., 2006) as well as composite impulsivity and narcissism subscales of the APSD was also included as covariates.
Statistical Analyses
An ordinary least squares regression (OLS) regression was used to examine whether CU traits were associated with bilateral amygdala volume. To examine sex and brain hemisphere differences, a repeated measures analysis of covariance (ANCOVA) was used. Post hoc t tests were run for any significant interaction effects observed as part of the second aim. Analyses were only run on participants who had complete data on all study measures (n = 272; see Supplemental Table S2). Unstandardized coefficients are reported in the results. To determine whether any significant findings were specific to the amygdala, the hippocampus was identified a priori as a comparison region to be used in a specificity analysis because prior research (Fish et al., 2020) has suggested that the amygdala and hippocampus are both implicated in sex-related, developmentally emergent psychopathology. The hippocampus is a subcortical region adjacent to and closely connected with the amygdala. Moreover, asymmetries in the hippocampus have previously been identified in antisocial individuals (Raine et al., 1998, 2004).
Results
Table 1 displays the descriptive statistics of the study variables. In this sample, males had larger left and right amygdala volumes and larger intercranial volumes than females. Males also exhibited higher levels of CU traits, narcissism, and impulsivity than females. Females were more pubertally advanced than males. No other significant sex differences emerged.
Descriptive Statistics of Study Variables.
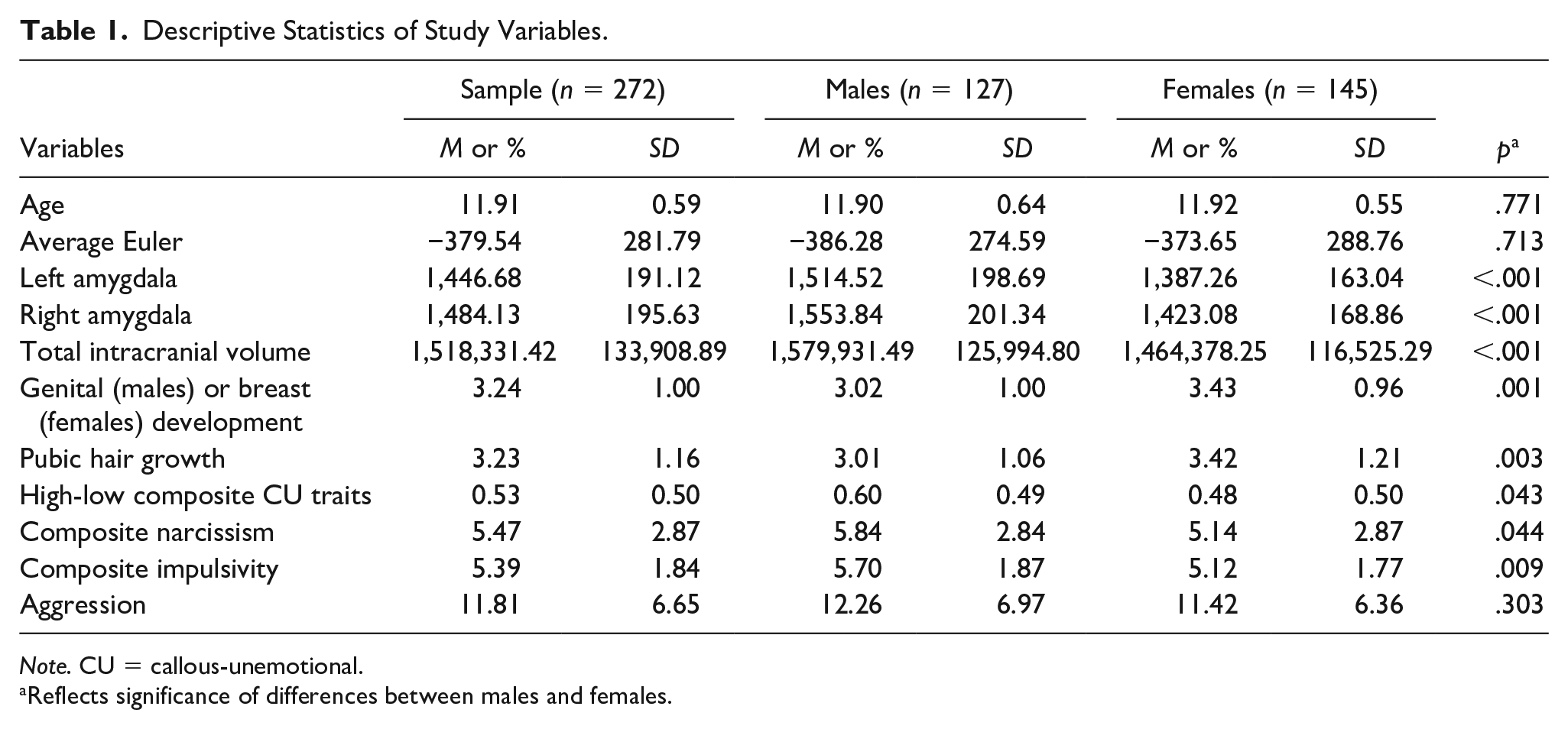
Note. CU = callous-unemotional.
Reflects significance of differences between males and females.
To address the primary research question, a hierarchical OLS regression was conducted. Model 1 included only covariates, whereas Model 2 included the covariates and the high-low composite CU traits measure. As shown in Table 2, CU traits were not associated with total amygdala volume (b = 14.94, SE = 39.31, p = .704).
OLS Regression Models for Effect of Covariates and CU traits on Total Amygdala Volume.
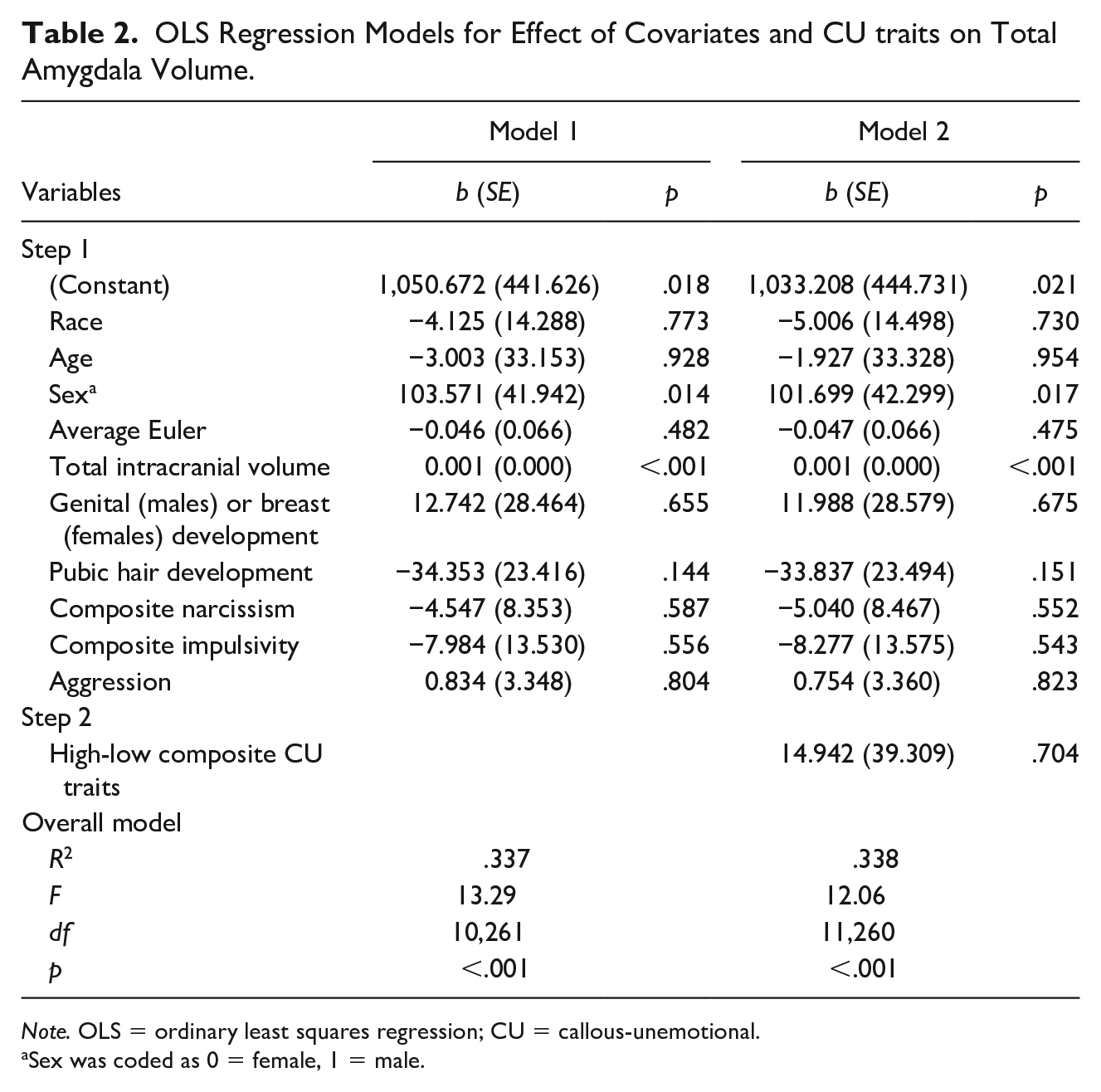
Note. OLS = ordinary least squares regression; CU = callous-unemotional.
Sex was coded as 0 = female, 1 = male.
To investigate sex and brain hemisphere differences, a repeated measures ANCOVA was conducted. Age, race, average Euler, total intracranial volume, both measures of pubertal development, composite narcissism, composite impulsivity, and aggression were entered as covariates. Left and right amygdala volumes were entered as the within-subject factor, whereas sex and the high-low composite CU traits measure were entered as between-subject factors. As shown in Figure 1, this analysis revealed a significant three-way interaction, F(1, 260) = 7.07, p = .008. Post hoc analyses were conducted to examine this interaction. These analyses revealed that the right amygdala was significantly larger than the left amygdala in females with high CU traits (p < .001), but there were no differences between left and right amygdala in females with low CU traits (p = .307). The opposite pattern was observed in males, with the right amygdala being significantly larger than the left amygdala in males with low CU traits (p = .002), but no differences between left and right amygdala in males with high CU traits (p = .065). No other differences emerged.
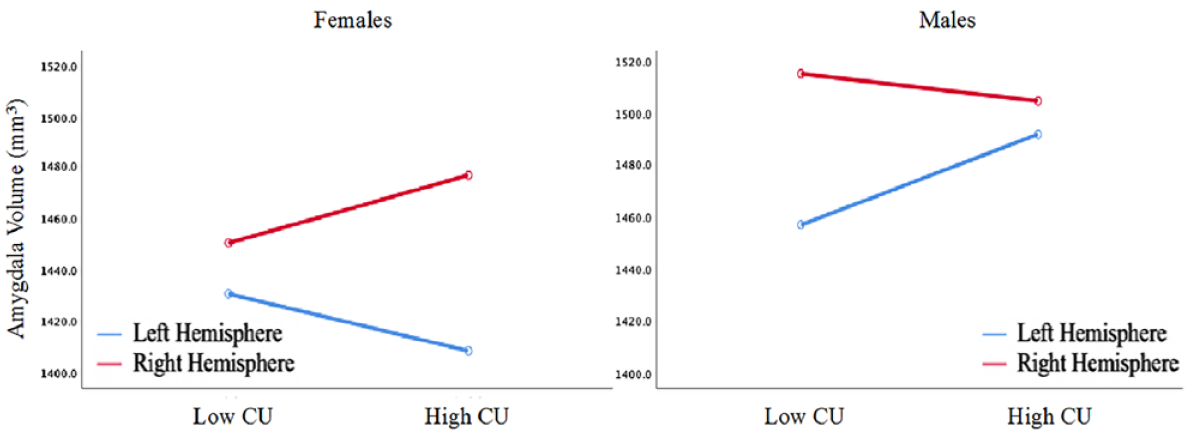
Illustration of amygdala asymmetries in high and low CU groups for male and female youth.
Specificity Check
A repeated measures ANCOVA was conducted with the hippocampus as the brain region instead of the amygdala. Age, race, average Euler, total intracranial volume, both measures of pubertal development, composite narcissism, composite impulsivity, and aggression were entered as covariates. Left and right hippocampal volumes were entered as the within-subject factor, whereas sex and the high-low composite CU traits measure were entered as between-subject factors. This analysis yielded no main or interaction effects (see Supplemental Table S3 and Supplemental Figure S1).
Discussion
This study aimed to examine the relationship between amygdala volume and CU traits in children aged 11 to 13 years. In addition to examining the direct relationship between amygdala structure and CU traits, sex and hemisphere asymmetries were explored to determine whether the relationship between CU traits and amygdala volume might differ as a function of brain or demographic characteristics. CU traits were not associated with amygdala volume in the overall sample. However, a significant sex × CU traits × brain hemisphere interaction was found. This interaction indicated that females with high CU traits show an exaggerated R > L asymmetry in amygdala volume, whereas females with low CU traits did not exhibit an asymmetry between the left and right amygdala; for males, the opposite pattern emerged such that a larger R > L amygdala was found in males with low CU traits but there was no amygdala hemisphere asymmetry in males with high CU traits. Moreover, this finding was not a function of age, pubertal development, nor differences in aggression, narcissism, or impulsivity. Specificity to the amygdala was documented as no such significant effects were found for the hippocampus. Findings are the first to the authors’ knowledge to document divergent sex-related amygdala asymmetries in relation to CU traits and suggest a neurodevelopmental basis to these psychopathic-like characteristics in children.
The pattern observed in males with low CU traits (R > L) and females with low CU traits (no asymmetry) are consistent with the asymmetry findings reported by Uematsu et al. (2012), who reported rightward amygdala asymmetry in the amygdala for normally developing males and no amygdala asymmetry for normally developing females. In the current study, the opposite pattern of hemispheric asymmetries between males and females with high versus low CU may have resulted in a canceling effect and could explain why there were null main effects. The divergent pattern of asymmetry found in the current study consisting of youth with high CU (R > L in females and lack of asymmetry in males) may reflect neurodevelopmental differences that are important to consider, particularly because they may reflect disrupted biological processes rather than environmental influences that may be more likely to result in bilateral hemispheric effects (Raine et al., 2004; Visser et al., 2014). Relatedly, another potential interpretation is that, assuming high levels of CU traits characterize males while low levels characterize females, “sex-consistent” levels of CU are characterized by equivalent left and right amygdala volumes while “sex-inconsistent” levels of CU are characterized by rightward amygdala asymmetry (see Figure 1). Future studies should investigate these differences to determine whether the current findings reflect developmental (e.g., time-specific growth differences) or fundamental (e.g., differences present across development) differences between sexes, as suggested elsewhere (Raschle et al., 2018).
Findings provide some broad support for a neurodevelopmental perspective of CU traits. In recent years, there has been increasing interest in identifying the neurodevelopmental basis of various clinical conditions including adult antisocial personality and has been extended to childhood conditions such as disruptive behavior and conduct disorder (Fairchild, Van Goozen, et al., 2013; McDonough-Caplan & Beauchaine, 2018; Raine, 2018; Wakschlag et al., 2018). Adopting a neurodevelopmental framework for understanding behavioral phenotypes that emerge early in life, such as CU traits, aligns with the theoretical perspective initially proposed by Moffitt (1993), who suggests that neuropsychological abnormalities may underlie early-onset behavioral problems that are more likely to persist throughout development and into adulthood. Indeed, CU traits have been suggested to be a precursor to the affective features of adult psychopathy, a condition that has been argued to be a neurodevelopmental disorder (Glenn, 2019).
In addition to supporting a neurodevelopmental perspective, the current findings contribute to two other bodies of the emerging literature: one reflecting sex-related asymmetries in antisocial individuals and another reflecting hemisphere-related subcortical asymmetries in antisocial individuals. There is emerging evidence that sex-related asymmetries can be observed in relation to various antisocial outcomes. For example, social and emotional impairments in males are associated with right ventromedial prefrontal cortex (VMPFC) damage but not left VMPFC damage; however, these deficits are observed in females with left VMPFC damage but not right VMPFC damage (Tranel et al., 2005). Moreover, these differences are not limited to cortical brain structures—they can also be observed in subcortical structures. Visser et al. (2014), for example, found that total hippocampal volume was associated with aggression only for females. In addition, as referred to earlier, hemispheric differences have been found in antisocial individuals. Unsuccessful male psychopaths exhibit larger right than left hippocampal volume (Raine et al., 2004) and murderers have greater right than left amygdala activity compared with age- and sex-matched controls (Raine et al., 1997). However, studies examining both sex- and hemisphere-related asymmetries are scarce. The findings of the current study need to be replicated, and the mechanisms to explain how these asymmetries influence the CU traits—amygdala volume relationship needs to be elucidated.
The current study utilized a sample that offered three key advantages to overcome the limitations of prior work: (a) while previous studies typically utilized male-only samples, the current study involved male and female youth; (b) prior studies were often limited by small sample sizes, whereas the current study yielded a larger sample size than any prior study (see Bolhuis et al. (2019), who utilized a younger sample with a wider age range, and Caldwell et al. (2019), who utilized a larger sample of incarcerated youth for exceptions); and (c) unlike prior studies, the current study utilized both a restricted age range and accounted for differences in pubertal development. Given that this study had approximately equal numbers of males and females, it was possible to examine sex differences, thus contributing to a growing body of work answering calls for increased research on sex differences in biological functioning and behavior (National Institute of Mental Health, 2011; National Institutes of Health, 2015). Importantly, sexual dimorphisms emerge more prominently during early adolescence, when individuals begin to undergo pubertal development (Goddings et al., 2014; Neufang et al., 2009). Amygdala volume in particular changes in a sex-dependent manner and as a function of pubertal stage (Neufang et al., 2009). Unlike previous studies, age, pubertal stage, and sex were adjusted for when examining how CU traits were associated with amygdala volume in the current study. Our secondary analyses revealed sex differences that did not seem to be a consequence of age or pubertal stage. Although these findings suggest sex and brain hemisphere are important to consider when examining the relationship between CU traits and amygdala volume, future studies should consider the mechanisms by which these factors influence this relationship.
There are some limitations to acknowledge. First, this was a cross-sectional study. A longitudinal design would allow examining whether the reported asymmetries continue to persist through adolescence and into adulthood. Second, the current sample consists of both males and females aged 11 to 13 years. However, the onset of puberty and related changes in brain development generally occur earlier in females than in males (Giedd et al., 2006; Goddings et al., 2014; Uematsu et al., 2012). The current study capitalizes on its sample’s narrow age range and controlled for two measures of pubertal development to account for potential influences of age and puberty, but there may remain important developmental changes that are not adequately captured by these measures—such as changes in hormone levels—which could be important to consider. In addition, the use of child- and parent-reported APSD data may be a limitation as they rely on the informants to accurately report on behavior. However, the current study utilizes a combined-informant scoring method for the APSD, which is thought to provide a more accurate depiction of behavior (White et al., 2009).
Despite these limitations, the current study provides potentially important preliminary insight into the relationship between CU traits and amygdala volume. In this sample, females with high CU traits show an exaggerated amygdala volume asymmetry (right > left) compared with their counterparts with low CU traits, whereas males with high CU traits fail to show an amygdala volume asymmetry (right = left) compared with their counterparts with low CU traits. This finding was not a function of age, pubertal development, nor differences in aggression, narcissism, or impulsivity. These divergent findings may reflect neurodevelopmental differences, which may suggest a neurodevelopmental basis—in part—to CU traits, but also warrant replication and further investigation.
Supplemental Material
SupplementalMaterials – Supplemental material for Divergent Amygdala Volume Asymmetries for Male and Female Youth With High Versus Low Callous-Unemotional Traits
Supplemental material, SupplementalMaterials for Divergent Amygdala Volume Asymmetries for Male and Female Youth With High Versus Low Callous-Unemotional Traits by Shichun Ling, Adrian Raine, Rebecca E. Waller, Kosha Ruparel, James Loughead and Ruben C. Gur in Crime & Delinquency
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.