
Abstract
During orthodontic tooth movement, the activation of the vascular system in the compressed periodontal ligament (PDL) is an indispensable process in tissue remodeling. We hypothesized that compressive force would induce angiogenesis of PDL through the production of vascular endothelial growth factor (VEGF). We examined the localization of VEGF in rat periodontal tissues during experimental tooth movement in vivo, and the effects of continuous compressive force on VEGF production and angiogenic activity in human PDL cells in vitro. PDL cells adjacent to hyalinized tissue and alveolar bone on the compressive side showed marked VEGF immunoreactivity. VEGF mRNA expression and production in PDL cells increased, and conditioned medium stimulated tube formation. These results indicate that continuous compressive force enhances VEGF production and angiogenic activity in PDL cells, which may contribute to periodontal remodeling, including angiogenesis, during orthodontic tooth movement.
INTRODUCTION
During orthodontic tooth movement, periodontal remodeling takes place in response to various mechanical stresses, such as compressive and tension forces. On the compressive side, the application of excessive orthodontic force induces circulatory failure, local ischemia, tissue hyalinization, and cell death in the periodontal ligament (PDL) (Reitan and Kvam, 1971; Rygh, 1974; Nakamura, 1982). Hyalinized tissue is then eliminated by scavenger cells such as multinucleated giant cells and macrophages (Kvam, 1972). Alveolar bone adjacent to the hyalinized tissue is also eliminated via the undermining of resorption by the adjacent bone marrow (Proffit, 2000). Finally, the compressed PDL returns to its original width, and connective tissue cells invade the degenerated tissues (Reitan and Rygh, 1994). Newly formed blood vessels in the degenerated tissue serve many functions, such as the recruitment of hematopoietic stem cells and mesenchymal cells through blood flow (Zaki and Vanhuysen, 1963) and the supply of nourishment and oxygen to the re-established tissue. Therefore, the activation of the vascular system in the compressed PDL is an indispensable process in periodontal remodeling. However, the role in angiogenesis of PDL cells subjected to compressive force remains unclear.
Vascular endothelial growth factor (VEGF) is the primary mediator of angiogenesis (Leung et al., 1989) and serves various biological functions, such as increasing vascular permeability (Senger et al., 1983) and promoting chemotaxis in human monocytes (Barleon et al., 1996; Clauss et al., 1996). VEGF is also involved in bone resorption (Niida et al., 1999; Nakagawa et al., 2000; Aldridge et al., 2005a) and formation (Mayr-Wohlfart et al., 2002; Street et al., 2002). Therefore, VEGF may play an important role in periodontal remodeling during orthodontic tooth movement.
Our objective was to clarify the involvement of VEGF in periodontal tissue remodeling on the pressure side during orthodontic tooth movement. The localization of VEGF in rat periodontal tissues during experimental tooth movement was examined in vivo. Moreover, the effects of continuous compressive force on VEGF production and angiogenic activity in human PDL cells were examined in vitro.
MATERIALS & METHODS
Experimental Tooth Movement
The study was approved by the Animal Care Committee of Tohoku University. The animals used were 6-week-old male Wistar rats. A uniform standardized compressive spring, made of 0.012-inch nickel-titanium wire (3M Unitek Dental Products, Monrovia, CA, USA), was placed in each animal’s mouth between the right and left upper first molars (Kanzaki et al., 2006), and compressive force was applied at 150 mN. We examined a total of 30 teeth in 15 rats. Experimental tooth movement was continued for 1 (n = 4), 3 (n = 4), 5 (n = 4), 7 (n = 4), 14 (n = 4), or 21 (n = 4) days. Teeth without movement were examined on days 0 (n = 2) and 21 (n = 4).
Immunohistochemical Staining
The experimental animals were killed under pentobarbital anesthesia and perfused. The maxillary bone, including the molars, was removed, fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 24 hrs at 4°C, and then rinsed in PBS. After samples were decalcified in 10% EDTA-PBS and embedded in paraffin, 5-μm-thick serial horizontal sections were cut from the root furcation level toward the root apex, within the range of 150 μm. VEGF expression in periodontal tissues of the mesiopalatal root of the maxillary first molars was examined. Sections were blocked and then immunostained with a 1:50 dilution of primary mouse anti-rat VEGF monoclonal antibody (Upstate Biotechnology, Lake Placid, NY, USA), followed by anti-mouse secondary antibody with streptavidin-conjugated horseradish peroxidase (Cell and Tissue Staining Kit, R&D Systems, Inc., Minneapolis, MN, USA; Chen et al., 2004). To visualize the immunoreactivity, we incubated the sections in a solution of diaminobenzidine. The sections were counterstained with hematoxylin for light microscopy and then dehydrated and mounted. Sections incubated without primary antibody were used as a negative control.
Primary Human Periodontal Ligament Cells
The protocol for this experiment was reviewed and approved by the Tohoku University of Dentistry Research Ethics Committee, and informed consent was obtained from all participants. Four teeth from four individuals (one female 18 yrs old, and three males 12, 16, and 22 yrs old) were used in this study. Pieces of PDL were obtained from the middle of the root of premolars extracted for orthodontic treatment. PDL tissues were cultured until confluence at 37°C under an atmosphere of 5% CO2 in α-minimum essential medium (α-MEM) supplemented with 20% fetal bovine serum (FBS) and antibiotics. PDL cells that proliferated were passaged from 4 to 8 times and used in all experiments.
Application of Compressive Force
We applied force to confluent cells by inserting glass wells (20 mm in diameter) containing lead pellets directly into the wells (35 mm in diameter) of a six-well plate containing PDL cells in 10% FBS α-MEM (Kanzaki et al., 2002; Goga et al., 2006). We adjusted the amount of compressive force by increasing or decreasing the number of pellets. PDL cells were subjected to loads of 0, 1.02 × 10−4, 2.04 × 10−4, 4.08 × 10−4, or 6.12 × 10−4 N/cm2 for 24 hrs. The experiment was repeated 4 times.
Enzyme-linked Immunosorbent Assay (ELISA)
The amount of VEGF in conditioned medium drawn from PDL cell cultures was determined with the use of a human VEGF ELISA kit (QuantikineTM, Human VEGF, R&D Systems, Inc., Minneapolis, MN, USA), according to the manufacturer’s instructions.
Real-time Reverse-transcriptase-Polymerase Chain-reaction (RT-PCR)
Total RNA was isolated from PDL cells by means of an RNeasy Mini kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. First-strand cDNA synthesis was performed with Ready-To-Go You-Prime First-Strand Beads (Amersham Pharmacia Biotech, Buckinghamshire, UK) and Oligo(dT)15 primer (Promega, Madison, WI, USA). This cDNA was used as a template in the subsequent real-time PCR analysis, performed with LightCycler® FastStart DNA Master SYBER Green I and a LightCycler system (Roche Diagnostics, Mannheim, Germany). The VEGF sense primer was 5′-cga agt ggt gaa gtt cat g-3′ (corresponding to nucleotides +170 to +192 of the VEGF coding region), and the antisense primer was 5′-ttc tgt atc agt ctt tcc tgg tga g-3′ (corresponding to nucleotides +681 to +705) (Janota et al., 2003). VEGF gene expression was normalized against GAPDH expression, which was determined with the sense primer 5′-ctt gtc atc aac ggg aag c-3′ (corresponding to nucleotides +239 to +257 of the GADPH coding region) and the antisense primer 5′-caa agt tgt cat gga tga cc-3′ (corresponding to nucleotides +545 to +526).
In vitro Angiogenic Activity
Capillary-like tube formation was assessed by means of an angiogenesis kit (Kurabo, Osaka, Japan), according to the manufacturer’s instructions. Briefly, human umbilical vein endothelial cells (HUVECs), co-cultured with normal human dermal fibroblasts (NHDFs) in 24-well plates, were incubated for 11 days with 50% of the culture supernatant from PDL cell culture. To confirm whether VEGF produced in compressed PDL cells enhances angiogenesis, we tested the conditioned medium alone or in combination with a neutralizing antibody against VEGF (Immuno-Biological Laboratories, Gunma, Japan), using methods previously described (Yoshino et al., 2003). After incubation, the cells were fixed in 70% cold ethanol and incubated with mouse anti-human CD31 antibody, followed by incubation with an alkaline-phosphatase-conjugated goat anti-mouse antibody. We then stained the cells with 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium to visualize the tube network. In each assay, 5 randomly selected fields in each well were photographed with a digital camera (Nikon, Tokyo, Japan) under a light microscope (Nikon, Tokyo, Japan). We analyzed tube formation by measuring the lengths of tubes per well, using an analytical software package (Kurabo, Osaka, Japan).
Statistical Analysis
Data are expressed as the mean ± standard deviation (SD). Statistical analyses were carried out by one-way analysis of variance (ANOVA) and the Bonferroni/Dunn post hoc test.
RESULTS
Immunolocalization of VEGF in Periodontal Tissues during Experimental Tooth Movement
VEGF immunoreactivity was observed in vascular endothelial cells, osteoblasts, and fibroblasts in periodontal tissues from both control and experimental animals (Fig. 1). In the control animals, PDL cells showed uniform VEGF expression on days 0 (Fig. 1A) and 21 (see Appendix Figure). In the experimental animals, highly compressed areas of the periodontal ligament and locally necrotic areas, so called “hyalinized tissues”, were observed 1–7 days after the initiation of tooth movement (Figs. 1B, 1C). Relatively strong VEGF immunoreactivity was observed in fibroblasts adjacent to hyalinized tissue and alveolar bone in the compressive area (Figs. 1B, 1C). On day 1, VEGF expression was also observed in mononuclear cells and fibroblasts (Fig. 1E). VEGF-positive osteoclasts in resorption lacunae and multinucleated giant cells along hyalinized tissues increased in number on day 7, when undermining resorption of the alveolar surface became apparent (Fig. 1F). VEGF expression returned to control levels by the end of the experimental period (Fig. 1D).
VEGF Production in Periodontal Ligament Cells in Response to Compressive Force
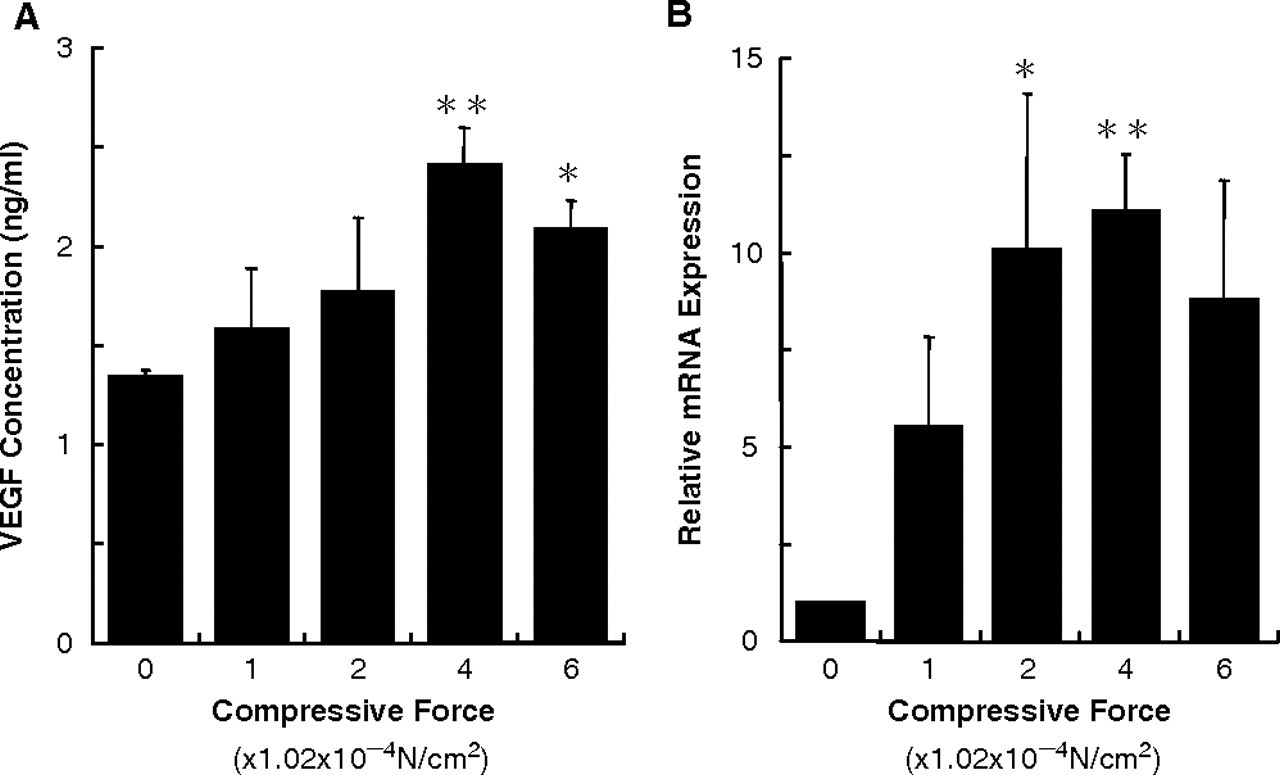
Continuous compressive force enhanced VEGF production in PDL cells (Fig. 2A). VEGF production increased in a load- dependent manner for loads up to 4.08 × 10−4 N/cm2, but decreased at the load of 6.02 × 10−4 N/cm2.
VEGF mRNA Expression in Periodontal Ligament Cells
Continuous compressive force up-regulated VEGF mRNA expression in PDL cells (Fig. 2B). The expression of VEGF increased in a load-dependent manner for loads up to 4.08 × 10−4 N/cm2, but decreased at the load of 6.02 × 10−4 N/cm2.
In vitro Angiogenic Activity of Conditioned Medium from Compressed Periodontal Ligament Cell Culture
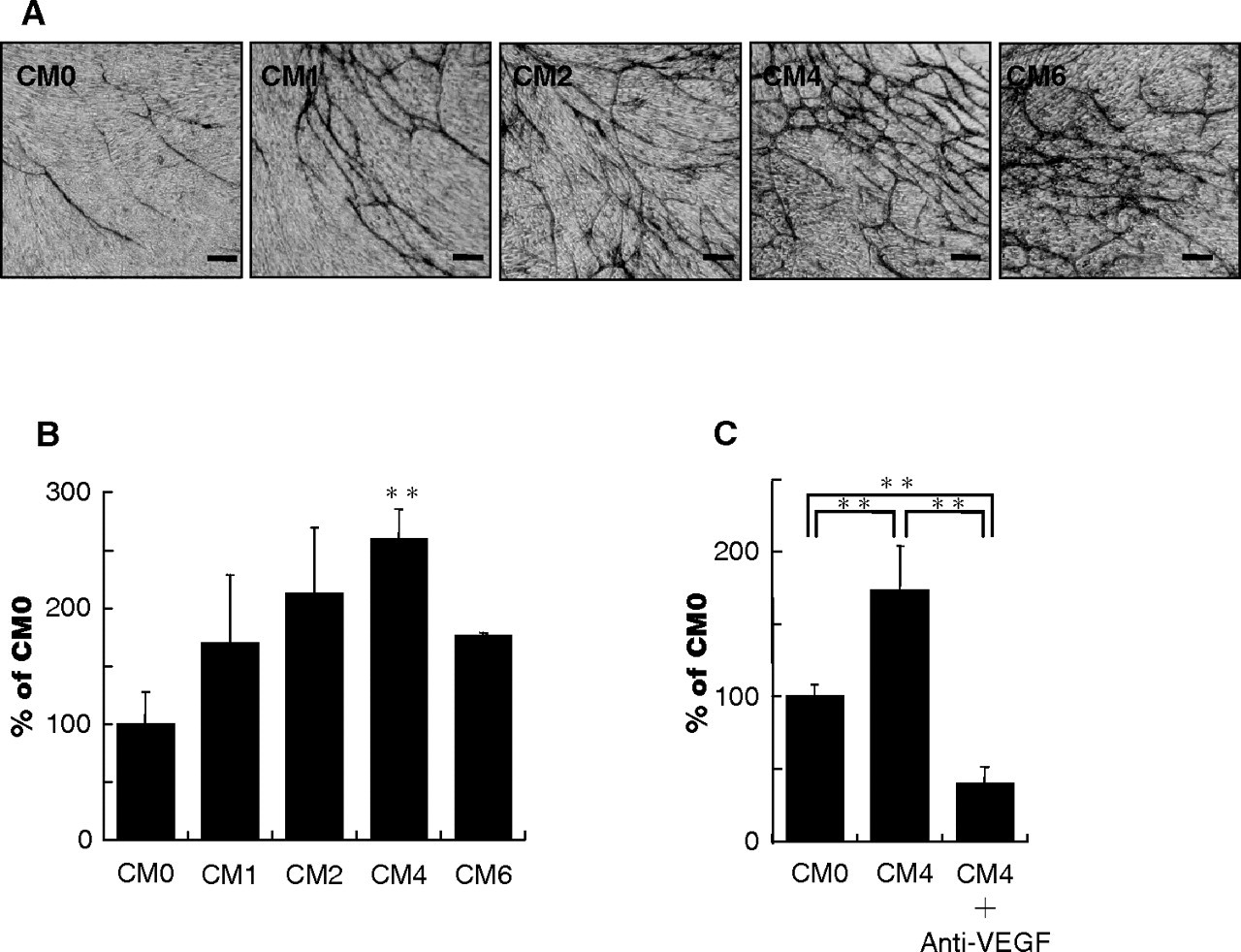
Continuous compressive force enhanced the in vitro angiogenic activity of conditioned medium from PDL cell culture that had been compressed for 24 hrs (Figs. 3A, 3B). The tube-forming activity increased in a load-dependent manner for loads up to 4.08 × 10−4 N/cm2, but decreased at the load of 6.02 × 10−4 N/cm2. This effect was blocked by the addition of anti-VEGF antibody (Fig. 3C; p < 0.01).
DISCUSSION
This is the first demonstration of VEGF localization on the compressive side of rat periodontal tissues during experimental tooth movement in vivo. PDL cells adjacent to hyalinized tissues and alveolar bone on the compressive side were strongly positive for VEGF at 1 wk after the start of tooth movement, suggesting that VEGF may be involved in the early stages of periodontal remodeling during orthodontic tooth movement, during which rapid changes in local blood circulation occur. However, it is unknown whether the applied orthodontic force directly induced VEGF production in PDL cells. Other local conditions, such as low oxygen, low nourishment, or inflammation, may have influenced VEGF production in PDL cells in the compressed area during tooth movement. To clarify this point, we performed an in vitro study to investigate the effects of continuous compressive force on VEGF production in cultured PDL cells.
VEGF mRNA expression and synthesis increased in a load- dependent manner for loads up to 4.08 × 10−4 N/cm2, but then decreased at the load of 6.02 × 10−4 N/cm2. These results indicate that there is an optimal compressive force for VEGF production in PDL cells, and that excessive force results in decreased VEGF production. Similar to its effect on VEGF mRNA expression and production, compressive force increased the angiogenic activity of conditioned medium from PDL cell culture. Furthermore, this increase was completely blocked by the addition of the neutralizing antibody. These results indicate that VEGF produced in compressed PDL cells may play a major role in angiogenesis, although other angiogenic factors, such as fibroblast growth factor-2, tumor necrosis factor -α, and/or transforming growth factor-β, may also be involved (Maruotti et al., 2006). Therefore, it is reasonable to assume that VEGF produced in compressed PDL cells could promote angiogenesis in hyalinized tissues and adjacent areas. Moreover, through the biological functions of VEGF, such as increasing vascular permeability (Senger et al., 1983) and promoting chemotaxis (Barleon et al., 1996; Clauss et al., 1996), blood vessels adjacent to hyalinized tissues may supply many additional cell types, such as fibroblasts, mesenchymal cells, macrophages, and multinuclear giant cells, to the degenerated tissues.
VEGF expression was observed in osteoclasts in resorption lacunae and in multinucleated giant cells appearing along hyalinized tissues on day 7, at which point undermining bone resorption was under way. There are two possible explanations for this observation. First, VEGF may have bound VEGF receptors (Flt-1, Flk-1) expressed on the cell surfaces of osteoclasts (Tombran-Tink and Barnstable, 2004; Aldridge et al., 2005b) and monocyte-macrophage lineage cells (Sawano et al., 2001). Alternatively, these cells may have produced VEGF. VEGF is known to salvage osteoclast formation mediated by Flt-1 in M-CSF-absent op/op mice (Niida et al., 1999) and induces osteoclast differentiation through Flt-1 (Aldridge et al., 2005a). VEGF is essential for osteoclast recruitment into the growth plate in embryos (Engsig et al., 2000) and directly enhances osteoclastic bone resorption and survival in mature osteoclasts (Nakagawa et al., 2000). The local administration of recombinant human VEGF increases the number of osteoclasts induced by experimental tooth movement (Kaku et al., 2001). These reports suggest that VEGF expression in compressed periodontal tissue may play an important role in bone resorption, as well as angiogenesis, on the compressive side.
Moderate VEGF expression was also evident in PDL cells and osteoblasts on the tension side. This is consistent with the demonstration of VEGF expression in osteoblasts on the tension side of mouse incisors (Kohno et al., 2003). Cyclic tensile force induces VEGF production in PDL cells (Yoshino et al., 2003) and osteoblasts (Motokawa et al., 2005). These results may explain the observation of constitutive VEGF expression in PDL cells, which are subject to intermittent chewing forces, regardless of tooth movement. Constitutive VEGF expression may contribute to PDL homeostasis by regulating local blood circulation and bone metabolism. Other studies have shown that VEGF stimulates bone repair by promoting angiogenesis and bone turnover (Street et al., 2002) and chemotactic migration of human osteoblasts (Mayr-Wohlfart et al., 2002). Taken together, these findings suggest that VEGF plays a pivotal role in periodontal remodeling during orthodontic tooth movement by acting directly on bone resorption and formation and indirectly on angiogenesis.
Immunohistochemical staining for vascular endothelial growth factor (VEGF) in periodontal tissues of the mesiopalatal root of maxillary first molars in rats. Expression in control animals on days 0 ( VEGF protein levels in supernatant collected from compressed PDL cells (
In vitro angiogenic activity of conditioned medium from cultured PDL cells. Conditioned medium from PDL cell culture subjected to loads of 0 (CM0), 1.02 × 10−4 (CM1), 2.04 × 10−4 (CM2), 4.08 × 10−4 (CM4), or 6.12× 10−4 (CM6) N/cm2 for 24 hrs was analyzed. CM0 is a control. Representative immunostaining for human umbilical vein endothelial cells (HUVECs) with anti-CD31 antibody to visualize the tube network. Magnification, 40×. Bar = 100 μm (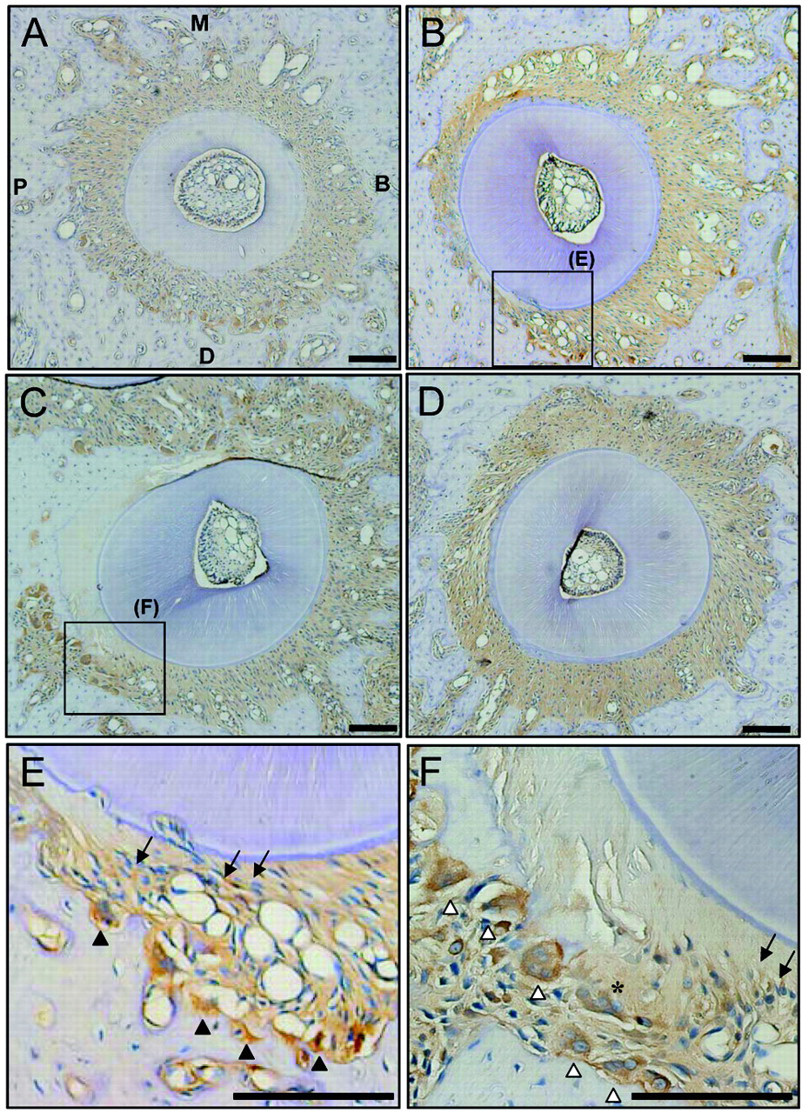
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.