
Abstract
Oral biofilms develop under a range of different conditions and different environments. This review will discuss emerging concepts in microbial ecology and how they relate to oral biofilm development and the treatment of oral diseases. Clues to how oral biofilms develop may lie in other complex systems, such as interactions between host and gut microbiota, and even in factors that affect biofilm development on leaf surfaces. Most of the conditions under which oral biofilms develop are tightly linked to the overall health and biology of the host. Advances in molecular techniques have led to a greater appreciation of the diversity of human microbiota, the extent of interactions with the human host, and how that relates to inter-individual variation. As a consequence, plaque development may no longer be thought of as a generic process, but rather as a highly individualized process, which has ramifications for the treatment of the diseases it causes.
THE HUMAN ORAL MICROBIOME
The human oral microbiome is comprised of hundreds of micro-organisms that colonize a variety of surfaces, many on tooth surfaces growing as a biofilm known as dental plaque. Dental plaque consists of at least 800 bacterial species (Paster et al., 2001, 2006; Becker et al., 2002; Aas et al., 2005, 2008; Preza et al., 2008), and this number is expected to rise into the thousands with the advances in mass sequencing techniques (Dethlefsen et al., 2007; Keijser et al., 2008).
MICROBIAL ECOLOGICAL FACTORS THAT AFFECT ORAL BIOFILM DEVELOPMENT
Oral biofilms develop under a range of different conditions and different environments. Factors that affect oral biofilm development, including inter-bacterial co-adhesion, pH, oxygen, and nutrients, have been well-reviewed recently in other articles (Burne and Marquis, 2000; Overman, 2000; Rosan and Lamont, 2000; Socransky and Haffajee, 2002; Marsh, 2005; ten Cate, 2006; Sissons et al., 2007). The aim of this review is to discuss emerging concepts in microbial ecology, particularly how they relate to oral biofilm development and how they are pertinent to the treatment of oral diseases.
Microbial ecology is the relationship of micro-organisms with one another, and with their environment (Konopka, 2006). A microbial ecosystem has been defined (Raes and Bork, 2008) as a system that consists of all the micro-organisms that live in a certain area or niche, which function together in the context of other biotic (plant and animals) and abiotic (temperature, chemical composition, and structure of the surroundings) factors of the niche.
Dental plaque is a dynamic and extremely complex oral biofilm ecosystem (Bowden, 2000; Marsh, 2003, 2006; Aas et al., 2005; Beighton, 2005; Jenkinson and Lamont, 2005; Socransky and Haffajee, 2005; Marsh and Percival, 2006; Haffajee et al., 2008). The microbial species in plaque form communities which establish an assortment of micro-niches, metabolic functions, and inter- and intra-species interactions (Marsh, 2003; Jenkinson and Lamont, 2005; Kolenbrander et al., 2006; Kuramitsu et al., 2007). The details of the complex web of microbial interactions are outside the scope of this review. For examples of specific cell-to-cell interactions and communication, the reader is directed to articles such as those by Kreth et al.(2008), Suntharalingam and Cvitovitch (2005), and Wen and Burne (2004).
Although dental plaque research spans 120 years, the view that dental plaque is a biofilm, and a microbial ecosystem, is relatively new. A definitive article that described dental plaque as a biofilm, and the diseases it causes, from an ecological perspective, was written in 1991 (Marsh, 1991). Marsh proposed the now-widely-accepted idea that a change in a key environmental factor (or factors) will trigger a shift in the balance of the resident plaque microflora to a disease-associated species composition—the ecological plaque hypothesis.
Although dental plaque is recognized as a complex microbial system, the experimental studies that investigated dental plaque from a microbial ecological perspective are substantially fewer than those that described single-species or selected bacterial consortia. To advance our understanding of dental plaque and the factors which influence its development, we must view plaque from a microbial ecological perspective. Clues to plaque development processes may lie in other complex systems, such as the interactions between the host and the gut microbiota, and even in factors that affect biofilm development on leaf surfaces (Monier and Lindow, 2005). Plaque development is often described as a fairly generic process. However, advances in molecular techniques across the whole field of microbial ecology, particularly mass sequencing techniques, indicate that generic descriptions of polymicrobial systems seriously underestimate their full complexity, especially those governed by multi-factorial processes.
THE ECOLOGY OF DENTAL PLAQUE IN HEALTH AND DISEASE
Many, if not all, conditions under which oral biofilms develop are tightly linked to the overall health and biology of the host, and vice versa. Because of this tight association, it is important that we first outline the ecology of dental plaque in health and disease, before discussing the ecological processes that affect oral biofilm development.
In health, there is an ecological balance between the human host and the indigenous micro-organisms (Jenkinson and Lamont, 2005; Marsh, 2006; Marsh and Percival, 2006). Under health conditions, dental plaque plays an essential role in natural host defense mechanisms; however, dental plaque is also the etiological agent associated with both dental caries and periodontitis (Jenkinson and Lamont, 2005; Socransky and Haffajee, 2005; Marsh, 2006; Marsh and Percival, 2006). Dental plaque bacteria are frequently faced with factors that challenge a health-compatible state, including exposure to high-sugar-content foods and tobacco smoke. Inefficient and insufficient oral hygiene practices, aging processes, genetic factors, and immune changes in the host also affect the oral environment and can generate conditions that encourage the plaque microbiota to a disease-associated state (Marsh, 2003; Jenkinson and Lamont, 2005). At another level, oral diseases are also affected by the individual’s socio-economic status (SES) and, particularly in the case of dental caries, by access to fluoridated water (Mignogna and Fedele, 2006).
The etiology of dental caries differs from that of periodontal disease, involving bacteria that have been described as metabolic and geographic opposites (Loesche, 2007). Dental caries occurs on teeth above the gum line (supragingivally), and periodontal disease occurs below (subgingivally), attacking the tooth-supporting tissues (Loesche, 2007). It is possible to have dental caries and periodontal diseases at the same time, and even on the same tooth. Due to the nature of the dynamics of the microbiota, it is also likely that treatment of one disease may affect the other (De Soete et al., 2005).
Much attention has been focused on identifying the bacteria which cause oral disease. It is of equal importance that bacteria associated with health also be identified, so that a microbiological goal for therapy can be established (Aas et al., 2005). However, it is extremely difficult to define a normal microflora, given the prevalence and complexity of these diseases, although recent research has indicated that there is a distinctive bacterial flora in the healthy oral cavity which is different from that of diseased oral cavities (Aas et al., 2005). Health appears to be associated with the lack of detection of certain species, such as Porphyromonas gingivalis, Treponema denticola, Streptococcus mutans, and Lactobacillus spp. (Eriksen and Dimitrov, 2003; Ledder et al., 2007), and elevated levels of other species, such as uncultivated phylotypes, Veillonella spp. oral clone X042 (Kumar et al., 2006), Deferribacteres clone W090, and clone BU063 from Bacteroidetes, Atopobium rimae, and Atopobium parvulum (Kumar et al., 2003).
Dental caries was once thought to be a simple disease. S. mutans was attributed as the sole etiological agent (Kleinberg, 2002), and caries research was dominated by, and treatments targeted at, this species. However, caries is unraveling as an extremely complex disease. Dental caries, measured clinically as demineralization of the tooth hard tissue, reflects the response of the tooth to a microbial challenge. The direct cause of dental caries is cariogenic plaque. Cariogenic plaques result when normally low populations of acido-genic and aciduric bacterial species, previously in balance with the oral environment and other plaque species, increase following high-frequency carbohydrate exposure (Marsh, 2003; Beighton, 2005). The metabolism of carbohydrate by these microbiota results in the acidification of plaque (pH < 5), and acid-induced demineralization of the enamel and dentin occurs, eventually resulting in cavitation. However, cavitation is a late event in the pathogenesis of decay (Loesche, 2007). Cariogenic plaques are comprised of numerous different microbial species, including S. mutans and other low-pH streptococci (Streptococcus oralis, Streptococcus mitis, Streptococcus anginosus), Rothia, Actinomyces, Lactobacilli and Bifidobacterium spp., and Candida albicans (Becker et al., 2002; Munson et al., 2004; Beighton, 2005; Aas et al., 2008; Preza et al., 2008).
Dietary patterns of carbohydrate consumption and a hierarchy of other genetic personal and social factors, such as oral hygiene habits, SES level, and ethnicity, also affect caries development (Gibson and Williams, 1999; Ramos-Gomez et al., 2002; Zero, 2004; Mignogna and Fedele, 2006). These strong genetic, personal, and social factors exert their effects by modifying the environment of the plaque biofilm sited on the tooth surface, its microbial communities, metabolism, and pH behavior, to shift the balance between health-compatible and pathogenic states (Bowden, 1991; Bradshaw and Marsh, 1998; Marsh, 2003).
Periodontal disease(s) reflects a cellular inflammatory response of the gingiva and surrounding connective tissue to the bacterial accumulations on teeth (Loesche, 2007; Kornman, 2008). These inflammatory responses are grouped into two clinical classifications, gingivitis and periodontitis. Gingivitis is measured clinically as inflamed gingiva and is extremely common (Loesche, 2007). Periodontitis, which is also very common, occurs when the plaque-induced inflammatory response in the gingival tissue results in the loss of collagen attachment between the tooth and the bone, and in bone loss (Loesche, 2007; Kornman, 2008). Gingivitis can enhance the progression to periodontitis (Sheiham, 1997). The etiology of periodontitis is also complex (Socransky et al., 1998; Paster et al., 2001, 2006; Socransky and Haffajee, 2005; Ledder et al., 2007). Research indicates that there are several key species that play a role in the disease process, and which have been grouped into microbial complexes based on clinical parameters, the disease stage, and the site in the oral cavity. These species include Porphyromonas gingivalis, Treponema denticola, Prevotella intermedia, Aggregatibacter actinomycetemcomitans, and Fusobacterium nucleatum. Advances in culture-independent approaches, such as quantitative ribosomal 16S cloning and sequencing techniques, have revealed other species which could play an important role in the disease process. These include Filifactor alocis, a Gram-positive anaerobe (Kumar et al., 2006), and new species, or phylotypes, including uncultivated clones from the Deferribacteres and Bacteroideetes phyla, Megasphaera clone BB166, and clone I025 from the TM7 phylum (Brinig et al., 2003; Kumar et al., 2003). It has been suggested that the TM7 phylum, in general, may play a role in the multifactorial process leading to periodontitis (Brinig et al., 2003; Kumar et al., 2003).
METAGENOMIC ANALYSIS OF THE ORAL MICROBIOME
Metagenomic approaches (also known as community genomics) used to explore the ecology of the oral microbiome have revealed new species and species associations with health and disease, as described above. However, it is important to note that although these culture-independent techniques are powerful and produce a wealth of information, they are not without their limits (Zoetendal et al., 2004; Sakamoto et al., 2005; Steward and Rappe, 2007). Many of the new species that have been described in the above metagenomic studies of dental plaque—e.g., Kumar et al.(2003), Aas et al.(2008), Preza et al.(2008)—have been identified with 16S Ribosomal RNA (rRNA) analysis. This is a technique which is based on the isolation of DNA from the target environment, polymerase chain-reaction (PCR) amplification of the rRNA gene, cloning the amplicons into Escherichia coli, and sequence analysis of the cloned 16S rRNA gene inserts. However, for some oral species, the 16S rRNA approach has been shown to have a limited capacity to discriminate between species of the same genus, particularly those from Veillonella (Beighton et al., 2008). Veillonella spp. are routinely isolated from the oral cavity and are found on all the mucosal and tooth surfaces; however, the representation of these species in some sequencing studies has been reported only at the genus level or limited to only three species: V. dispar, V. parvula, and V. atypica (Aas et al., 2005; Haraszthy et al., 2007). The limitation of species-level discernment could potentially be attributed to the type of primer that was used in the sequencing studies (Frank et al., 2008). In addition to potential primer limitations, 16S rRNA techniques are also subject to PCR bias, as are techniques such as PCR-denaturing gel gradient electrophoresis (PCR-DGGE) (Rasiah et al., 2005; Ledder et al., 2007).
Pyrosequencing, a next-generation sequencing technique, is a DNA sequencing technique which relies on the detection of pyrophosphate (PPi) released on the incorporation of nucleotides during DNA synthesis (Ronaghi, 2001). In a cascade of enzymatic reactions, visible light is generated that is proportional to the number of incorporated nucleotides. The cascade starts with a nucleic acid polymerization reaction in which inorganic PPi is released as a result of nucleotide incorporation by polymerase. The released PPi is subsequently converted to ATP by ATP sulfurylase, which provides the energy to luciferase to oxidize luciferin and generate light. Because the added nucleotide is known, the sequence of the template can be determined (Ronaghi, 2001). Pyrosequencing has been successfully applied to explore bacterial diversity in a range of ecosystems, from hydrothermal vents of a deep marine biosphere (Huber et al., 2007) to the differences in the vaginal microflora between HIV- and non-HIV-infected women (Spear et al., 2008). Recently, the first reported pyrosequencing analysis of the oral microbiome was published (Keijser et al., 2008). The findings of the pyrosequencing analysis of saliva and plaque from healthy adults indicated that diversity was one order of magnitude higher than has been previously described, and the authors estimated that there are at least 19,000 species-level phylotypes that contribute to the diversity of the oral microbiome (Keijser et al., 2008). Pyrosequencing techniques do not produce the full length of 16S rDNA sequences traditionally used in taxonomic studies, and they are expensive. However, as Keijser et al.(2008) stated, the approach that they used targeted the 16S rDNA hypervariable V6 region, which has the advantage of being flanked directly by a well-conserved region that can be used in PCR amplification and is an approach that has been validated in other studies (Huber et al., 2007; Sundquist et al., 2007). Keijser et al.(2008) also reported that, even using a conservative 6% difference in operational taxonomic units (OTUs), between 3600 and 6800 phylotypes were discernible. Even this more conservative interpretation indicates that the diversity of the oral microbiome is much greater than previously reported.
Cultivation techniques have largely been put to one side in favor of molecular analysis; however, they are beginning to receive more attention again, particularly in environmental ecosystems (Stott et al., 2008). The resurgence of cultivation analysis has come about since culture-independent techniques such as pyrosequencing do not provide information pertaining to the function and metabolic requirements of the bacteria that are identified. Learning how to grow as-yet-uncultivable bacteria is important in gaining a fuller understanding of the range of microbial taxonomic and metabolic biodiversity, which may also lead to important biotechnological discoveries (Stott et al., 2008). In the case of oral bacteria, cultivating as-yet-uncultivated species may identify their role in disease and allow for the development of specific antimicrobials.
INTER-INDIVIDUAL VARIATION IN THE ORAL MICROBIOME
Despite high levels of genetic similarity, humans respond differently to different stimuli, biological stressors, and environments (Relman, 2008). It is therefore not surprising that with the advances in molecular techniques (Steward and Rappe, 2007), the understanding of the human microbiome is receiving a fundamental re-evaluation, and with that, a greater appreciation of its diversity, genetic plasticity, and the extent of interactions with the human host (Dethlefsen et al., 2007; Bent and Forney, 2008; Ley et al., 2008). This has led to the recognition that although each microbial community contains micro-organisms from certain families and genera that are found in the same habitat in many or most individuals, at the species and strain levels, the microbiota of an individual may be as unique as a fingerprint (Dethlefsen et al., 2007).
In particular, studies of the gut microbiome have shown that the microbes co-evolved with their host, and are central in maintaining the host’s health (Hooper et al., 2002; Xu and Gordon, 2003; Bäckhed et al., 2005). This co-evolution is thought to be mutually beneficial—for example, gut microbiota are able to degrade complex polysaccharides as their food source and provide otherwise-inaccessible nutrients to the host (Hooper et al., 2002; Xu and Gordon, 2003; Bäckhed et al., 2005). Findings such as these are raising questions about whether attributes that have previously been assumed as human traits may in fact be microbial (Dethlefsen et al., 2007). Host-microbe interactions may vary according to the individual, thereby contributing to an individual’s unique microbial fingerprint. It has been proposed (Bäckhed et al., 2005) that the structure and composition of the gut microbiota reflect natural selection at two levels: at the microbial level, where lifestyle strategies (e.g., growth rate and substrate utilization patterns) affect the fitness of the individual bacteria in a competitive ensemble; and at the host level, where sub-optimal functionality of the microbial ensemble can reduce host fitness (Bäckhed et al., 2005). In terms of predicting health outcomes, the inter-individual variation of gut microbiota profiles observed in children has been proposed to represent a means of determining their potential weight problems as adults (Kalliomäki et al., 2008), and as a means of exploring the differences in healthy persons and those with celiac disease, a chronic inflammatory disorder of the small intestinal mucosa (Sanz et al., 2007). A similar health-disease predictive model is possible in oral communities. It is also possible that the oral microbiota may have a greater influence in regulating the environment in the oral cavity than is currently known. We are at the beginning of this area of research, which poses exciting and challenging avenues for the study of plaque development and oral microbial communities.
Analysis of data from our in vitro experiments, using dental plaque microcosms, indicates that the characteristics of the individual’s host environment are embedded in their plaque micro-biota, as evidenced by differences in plaque species composition and functional activity (Rasiah et al., 2005; Filoche et al., 2007, 2008). It is highly desirable to incorporate this level of inter-individual variation into laboratory models (Ledder et al., 2006), and necessary, so that responses of plaque microbiota to different therapies, and factors which drive disease, can be studied in depth. Although human studies are essential, they are constrained by the technical and ethical limitations to carrying out the kinds of experiments that are needed to elucidate the microbial ecology of dental plaque.
In a series of experiments, in vitro plaques were grown from the saliva of seven different individuals, in a chemically defined artificial saliva (DMM) (Wong and Sissons, 2001) with both 0.15% and 0.5% sucrose. The salivary and plaque microbial profiles were analyzed by checkerboard DNA-DNA hybridization (Filoche et al., 2007, 2008). Factor analysis of the data showed that the composition of the salivary microbiota differentiated according to either higher levels of periodontal, or to caries-associated pathogens (Filoche et al., 2008). In addition, the plaque microbiota responses, biomass yield, and pH response to sucrose supplementation were different for different individuals (Filoche et al., 2007, 2008). This suggests that the degree of responsiveness to sucrose, measured by the increase in the prevalence of caries-associated pathogens, is host-specific, which may translate into individual differences in the progression of caries, and in the treatment outcome.
A better understanding of inter-individual variation at the microbial level requires much more investigation, particularly for oral communities, which have received less “meta” attention compared with other microbial-human communities over the last few years.
INTER-INDIVIDUAL VARIATION IN PLAQUE DEVELOPMENT AND THE PROGRESSION OF DISEASE— THE ROLE OF MICROBIAL AGGREGATES
Oral biofilm formation is described as a highly ordered sequence of events involving the sequential attachment of particular species over time (Kolenbrander et al., 2006; Marsh, 2006), and it is the same for everyone. Distinct stages in plaque formation are described as: acquired pellicle formation; reversible adhesion involving weak long-range physicochemical interactions between the cell surface and the pellicle, which can lead to stronger adhesion-receptor-mediated attachment; co-adhesion resulting in attachment of secondary colonizers to already-attached cells; and multiplication and biofilm formation (including the synthesis of exopolysaccharides and, on occasion, detachment) (Marsh, 2006). This view on plaque formation has stemmed from the classic experiment by Ritz (1967) and has remained largely unchanged, and unchallenged, for 40 years. In Ritz’s investigation, samples of plaque were removed and plated periodically over a period of several days (Ritz, 1967). The results showed a progression of organisms, with the streptococci being the dominant pioneer species, followed by increasing proportions of actinomyces and, eventually, the conversion of the plaque to a mature community with high levels of Gram-negative anaerobic filamentous organisms (Ritz, 1967; Rosan and Lamont, 2000).
Given the evidence that shows the extent of inter-individual variation, plaque development must be person-specific. If how oral microbiota come together to grow and form plaque is in fact person-specific, the arrangement of species within plaque, and its resultant structure, will also be different, as will its function. How the very specific cell-cell interactions that have been identified over the years (Kolenbrander et al., 2006; Kuramitsu et al., 2007) are also played out should also be different in different individuals. Currently, the evidence to show that plaque development is person-specific is limited, but convincing. In a study which examined the initial plaque development in three humans wearing retrievable enamel chips, the initial plaque community, as measured by 16S rRNA gene sequencing and fluorescence in situ hybridization (FISH), of each person was shown to be unique in terms of diversity and composition (Diaz et al., 2006). The authors also proposed that, due to the repetitive and distinctive community composition within individuals, the spatiotemporal interactions and ecological shifts that accompany biofilm maturation could also occur in a person-dependent manner (Diaz et al., 2006), and this does appear to fit with Ritz’s model of plaque maturation.
In another investigation, initial biofilm formation was monitored in six persons who wore bovine buccal mounted enamel slabs (Hannig et al., 2007). Biofilm formation (cell attachment) was monitored after 3, 30, and 120 minutes and evaluated with 4′6-diamidino-2-phenylindole (DAPI), fluorescein diacetate and ethidium bromide (live/dead staining), and FISH of eubac-teria and streptococci. The results showed that the numbers of bacteria colonizing the enamel were different for each individual. The results of this study also showed that the bacteria were distributed randomly, in small aggregates.
The capacity of oral bacteria to form inter-specific interactions, in the form of co-aggregates, has been known for many years (Kolenbrander, 2000). Co-aggregation in non-oral bacteria has also been reported and has been proposed to be important in the role of biofilm development per se (Rickard et al., 2003a,b; Simões et al., 2008). We have also demonstrated that aggregates formed by three oral bacteria were central to surface attachment and growth, and to survivability after antimicrobial treatment (Filoche et al., 2004). Aggregate formation has been shown to facilitate the survival of both bacteria that immigrate onto leaf surfaces and those bacteria that are already resident (Monier and Lindow, 2005). Aggregates of bacteria have also been referred to as “planktonic biofilms,” enabling cells to function in a manner similar to multicellular organisms (Flemming and Wingender, 2001). It would appear that aggregates in saliva have the potential to contribute not only to the rapid accumulation of plaque on tooth surfaces, but also to the heterogeneity of plaque development, between individuals and within the oral cavity of an individual, and, as discussed in the preceding section, in biofilm proliferation (Figs. 1A, 1B).
It is possible that microbial aggregates present in saliva could also be related to the progression of oral diseases. Diabetes mellitus is a chronic metabolic disorder, characterized by hyperglycemia, and leading to significant systemic complications and increased morbidity and mortality in affected individuals (Lalla et al., 2007). Diabetes significantly increases the prevalence, severity, and rate of progression of periodontal disease, which is recognized as one of the complications of diabetes (Taylor, 2001). In this condition, microbial aggregates in saliva would present a greater challenge to the host’s immune system compared with single bacterial cells, and could potentially initiate periodontal disease, or facilitate its proliferation (Figs.1A, 1B). The involvement of aggregates in plaque development and disease propagation promises new avenues for the understanding, diagnosing, and controlling of dental diseases.
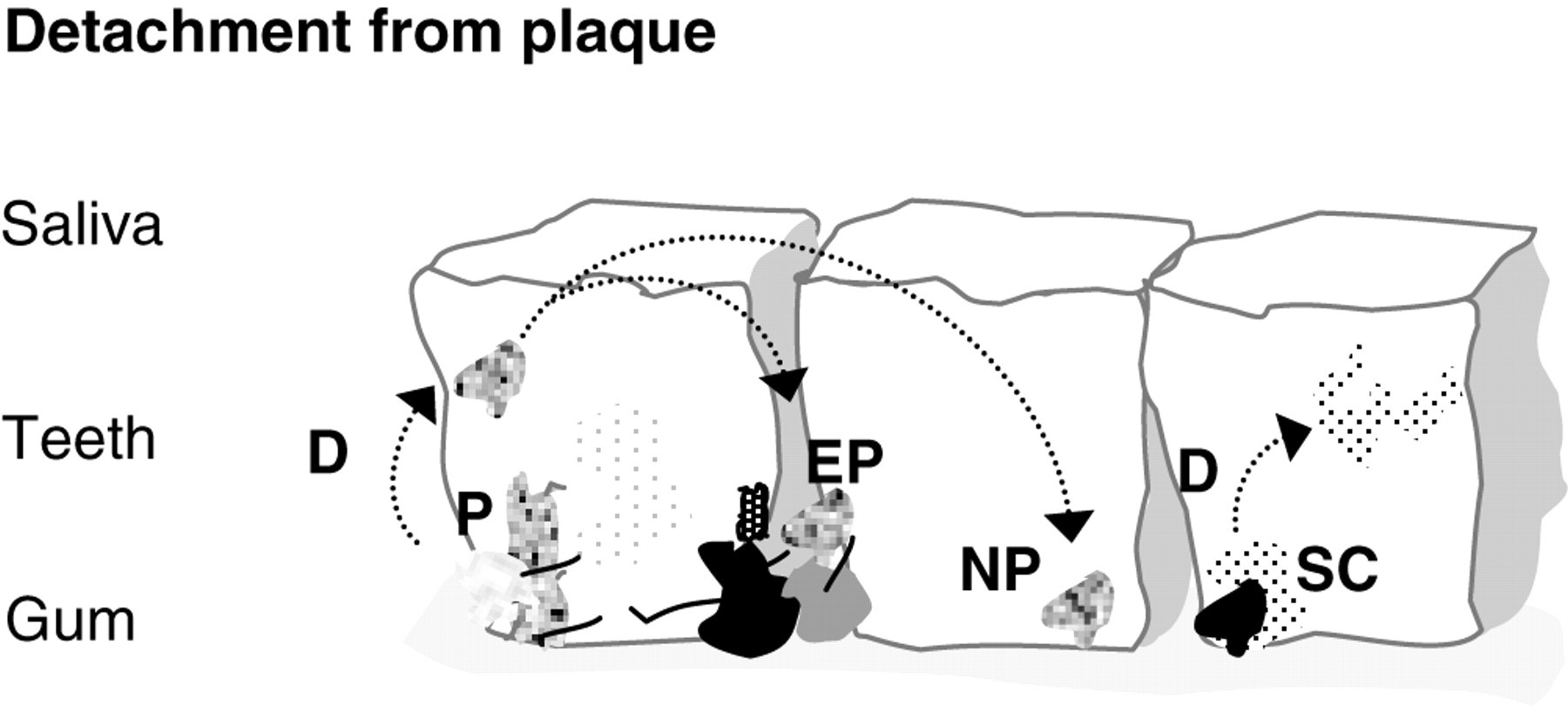
BIOFILM DETACHMENT AND DISPERSAL— ROLE OF MICROBIAL AGGREGATES
Considerable attention has been focused on the attachment of bacterial cells in the formation of a biofilm; however, the detachment of bacterial cells is also important in terms of the dissemination of infection and contamination of clinical and industrial systems (Stoodley et al., 2001). Little is known about the detachment of bacteria from the dental biofilm. In non-oral biofilm systems, the detachment of bacteria has been considered to be a passive process, which is influenced by fluid flow rates (shear stress) or starvation (Hall-Stoodley and Stoodley, 2005). However, detachment may also be a strategy by which bacteria proactively colonize new niches before space and nutrients become limited (Hall-Stoodley and Stoodley, 2005). It has been suggested (Hall-Stoodley and Stoodley, 2005) that there may be three types of dispersal strategies for biofilm bacteria—swarming dispersal, clumping dispersal, and surface dispersal—all of which would have a fluid (planktonic) phase and a surface-associated phase (Fig. 2). Findings from the above-described investigations indicate that oral bacteria would exhibit clumping (aggregate) dispersal strategies (Fig. 2), and that oral bacteria which are motile (e.g., T. denticola) would most closely fit swarming dispersal, which involves the release of single motile bacteria from the biofilm, as seen with Pseudomonas aeruginosa (Webb et al., 2003; Hall-Stoodley and Stoodley, 2005). Hall-Stoodley and Stoodley (2005) discussed how each strategy would have advantages and disadvantages over the other. For example, swimming or twitching motility, seen in swarming dispersal, has the advantage that the motile bacterium is self-propelled and directional (i.e., chemotactic). However, single cells do not have the protective advantage of being in a microbial aggregate. Conversely, cells in the microbial aggregate would have to rely on a non-directional, fluid-driven dispersal, but are in the protective confines of a microbial aggregate (Fig. 2). It is possible that oral biofilm detachment strategies could be a therapeutic target in the management of plaque-induced diseases, since they may limit the proliferation of oral biofilm development.
DEVELOPMENT OF HETEROGENEOUS PLAQUES—THE ROLES OF LOCATION AND SPATIAL PATTERNING
For microbial populations, geography is generally believed to have no impact whatsoever on distribution, a belief that is usually expressed as ‘everything is everywhere; the environment selects’ (O’Malley, 2008). However, it is now thought that microbial diversity is spatially structured by geographic location as well as by the environment (O’Malley, 2008). This may be important in our understanding of microbial ecological factors that influence oral diseases on two different scales. On a worldwide scale, the distribution of pathogens across the globe may be important (Rylev and Kilian, 2008). On the scale of the oral cavity, how spatial patterning of plaque development and diseased sites occurs is important (Mager et al., 2003; Dawes, 2008; Mineoka et al., 2008). Each surface in the mouth, even on the same tooth, presents a different environment, and as a result, the plaque that develops on these surfaces will be different (Mager et al., 2003; Dawes, 2008; Haffajee et al., 2009; Mineoka et al., 2008). Given the heterogeneous nature of the oral cavity, it is not surprising that location affects the development of plaque. Clinicians often observe the patchy distribution of diseased sites; however, there is a need to determine what drives these differences if treatment and their outcomes are to be improved.
In a recent review, it was highlighted that, for better understanding of biofilm processes and the development of a spatial dimension to biofilm, research is needed (Battin et al., 2007). Consequently, biofilm researchers were asked to view biofilms according to the tenets of landscape ecology (Battin et al., 2007), since it is a field of ecology which draws on many ecological principles, such as dispersion, ecosystem, and invasion ecology, and which focuses on the interactions between spatial heterogeneity and pattern—specifically, how to characterize it, where it comes from, why it matters, and how it changes through time. This approach is particularly relevant for oral biofilm formation and the progression of oral diseases, and it may prove useful in terms of analyzing the spatial patterning of dental plaque development.
In a study which looked at the effect of periodontal therapy on the composition of subgingival microbiota (Haffajee et al., 2006), a high degree of variation in the subgingival microbial profiles from site to site in the same person, and from person to person, was observed. However, it was also noted that many individuals showed common patterns of colonization. In another study, supragingival plaque samples were taken from the mesiobuccal aspect of each tooth in 187 persons (Haffajee et al., 2009). By checkerboard DNA-DNA hybridization analysis, the samples were analyzed for their content of 40 bacterial species, and significant differences in mean species counts and proportions were determined among tooth surfaces and six tooth-type categories: molars, bicuspids, and incisors/canines in the mandible and maxilla. The results showed that although some differences in plaque species composition could be accounted for by increased plaque biomass, it was the location of the tooth that influenced the proportions of certain species being present (Haffajee et al., 2009). Site-specific variations in plaque development could be linked to a series of competitive interactions between certain oral bacteria as the means of determining community structure, in relation to the geographical (site-specific) location on the teeth and within the oral cavity. If the spatial patterning of plaque development could be determined, including on an individual level, it may hold vital clues to the prediction of health and disease.
IMPLICATIONS FOR THE TREATMENT OF ORAL DISEASES— AN ECOLOGICAL APPROACH
Since the main drivers for deciphering factors that affect oral bio-film formation are a better understanding of the diseases dental plaque causes and improved treatments, it is appropriate to discuss how the above-described emerging principles and concepts in microbial ecology relate to the treatment of oral diseases.
The concept that caries and periodontal diseases arise as a result of environmental perturbations to the habitat has been encapsulated in the ecological plaque hypothesis, as discussed above (Marsh, 2003). Key features of the hypothesis are that the selection of pathogenic bacteria is directly coupled to changes in the environment, and that diseases need not have a specific etiology, since any species with relevant traits could contribute to the disease process (Marsh, 2003). As Marsh (2003) states, also implicit in this hypothesis is the concept that caries and periodontal diseases can be prevented, and managed, by direct inhibition of the putative pathogens, and by interference with the environmental factors that drive the selection and enrichment of these bacteria. Ultimately, this approach would lead to the re-establishment of a health-compatible plaque species composition. For this to be therapeutically achieved would mean re-adjusting ecological parameters to yield a stable, health-compatible condition, even after treatment has stopped.
It has recently been acknowledged that the goal for treating oral diseases, particularly dental caries, should be to reverse or halt disease progression by treating the underlying disease (Caufield et al., 2001; Guzmán-Armstrong and Warren, 2007; Selwitz et al., 2007). The medical model emphasizes the mechanical/chemical removal of plaque (oral hygiene), chemical (antimicrobial) modification of plaque, the use of fluorides, dietary modification, and salivary stimulation for better control of the underlying disease (Kidd and Fejerskov, 2003). This type of disease management would be minimally invasive and preserve the tooth. However, a global reduction in plaque biomass, particularly with chemotherapeutics, could result in deleterious effects (Caufield et al., 2001). For example, evidence suggests that after treatment with a strong antimicrobial such as chlorhexidine, a sub-population of bacteria could remain viable and able to proliferate (Gilbert et al., 2002; McBain et al., 2003). Given the resilience of these “persisters” to treatment, this situation could present an even greater antimicrobial challenge, and potentially a more pathogenic plaque, compared with the initial pre-treated plaque (Gilbert et al., 2002). Such a situation may also contribute to the recalcitrance of oral biofilms to treatment (Socransky and Haffajee, 2002) and the reported ineffectiveness of chlorhexidine as an anti-caries treatment (Fennis-le et al., 1998; Forgie et al., 2000; Sekino et al., 2004; Autio-Gold, 2008). For a health-compatible state to be restored chemothera-peutically, it is likely that a “nudge” approach would be more effective than a global eradication of plaque in modulating plaque species composition and therefore its function.
INTER-INDIVIDUAL VARIATION AND TREATMENT RESPONSES
Inter-individual variation represents a challenge for the treatment of polymicrobial diseases, such as dental caries and periodontitis. To treat these diseases effectively, one requires an understanding of normal healthy microflora so that the goal for antimicrobial therapy can be realized (Aas et al., 2005). The challenge lies in determining what is normal for that particular individual. Findings from studies that have looked at microbial associations in health and disease (Socransky et al., 1998; Becker et al., 2002; Kumar et al., 2006; Aas et al., 2008; Preza et al., 2008), microbial outcomes of antimicrobial treatment (Haffajee et al., 2006; Teles et al., 2006; Filoche et al., 2008), and factors which drive disease (Filoche et al., 2007; Sissons et al., 2007), suggest that different bacteria may be responsible for disease in different people, which suggests that disease progression is person-specific.
In a study which looked at the post-treatment effects of two oral antiseptics, in vitro plaques were grown from the saliva of five different individuals, in a chemically defined artificial saliva (DMM) (Wong and Sissons, 2001) with both 0.15% and 0.5% sucrose and treated with a single treatment of either chlorhexidine or Listerine™. Twenty-four hours post-treatment, factor analysis of the checkerboard DNA-DNA hybridization data revealed that the plaque microbiota composition was differentiated according to the originating saliva donors, and yielded a range of microbial responses to the same growth conditions and the same treatment (Filoche et al., 2008). The results from the investigation indicated that although similar microbiota responses to sucrose supplementation can occur with different individuals, this does not necessarily indicate that the responses to chemotherapeutic treatment will be the same. The results suggested that there are microbial factors additional to the growth environment—e.g., host specificity, bacterial strain variation, bacterial species arrangement in plaque—which could contribute to the outcome of chemotherapeutic treatment (Filoche et al., 2008).
In a study by Haffajee et al.(2006), 493 subjects were assessed in regard to 17 different treatments for periodontal disease. The data were obtained from a series of randomized clinical trials which had either been completed or were ongoing during the preceding 10 years at The Forsyth Institute (Boston, MA, USA). This approach provided an indication of the effects that periodontal therapy had on the subgingival microbiota overall, and a consideration of some of the effects that might be attributed to specific adjuncts. While the data analysis indicated that periodontal therapy, on average, provides a significant improvement in clinical parameters, there was a reduction in the levels of many subgingival species. The study participants also differed in the composition of their subgingival microbiota, because they were colonized by different species and by different levels of periodontal pathogens. It was also shown that the individuals responded quite differently to the same periodontal therapy. Similarly, in an evaluation of the microbiota associated with root caries, considerable inter-individual variation in both the levels and the types of particular species associated with the disease was shown (Preza et al., 2008).
In the case of antimicrobial treatment, inter-individual variation at the microbial level may be associated with prior anti- microbial exposure through medication, or use of different oral hygiene products. It may be that there are host-specific strains (Kilian et al., 2006, 2008; Rylev and Kilian, 2008) that yield different antimicrobial responses, and/or that different bacteria house the genes that are important in the functionality of disease (Ley et al., 2008; Zaneveld et al., 2008). It has been proposed (Haffajee et al., 2006) that differences in clinical responses to periodontal treatment are partly due to the host’s ability to cope with the infection, similarly indicated for dental caries (De Soet et al., 2008), and partly because of the nature of the colonizing subgingival species. Undoubtedly, factors such as host genetics, age, diet, place of residence, and perturbations (in the form of medication or drug use) affect the structure and behavior of all microbial-human communities.
BIOFILM STRUCTURE, ALTRUISTIC BEHAVIOR, AND ANTIMICROBIAL TREATMENT
There is an interplay between factors that affect biofilm development and the resultant biofilm structure. Factors that affect biofilm formation and development will also affect biofilm structure and architecture, and vice versa. The structure of the biofilm will influence the penetrability of antimicrobial compounds through surface cell layers and the extracellular matrix, potentially limiting their effectiveness (Stewart et al., 2001; Robinson et al., 2006). Reduced efficacy of topically applied agents has direct implications for the successful treatment outcomes of oral diseases.
In vivo dental plaques grown on enamel oral devices have been shown to consist of microbial “stacks” surrounded by voids and channels, exhibiting an open, fragmented architecture with a high surface-area:volume ratio in the outermost layers (Watson and Robinson, 2005). The void-and-channel system in plaque would serve as a route for the distribution of antimicrobial compounds. However, the presence and location of these delivery routes change with biofilm age, thickness, nutrient status, and external conditions (Characklis et al., 1990). In an electron microscopy study, oral biofilms on gingival epithelial cells demonstrated localized ultrastructural changes in response to chlorhexidine treatment (Vitkov et al., 2005). These ultra-structural changes could be sufficient to modify the channel fractions within the biofilm and delivery routes of subsequent antimicrobial treatment. Changes in the delivery routes potentially limit the distribution of antimicrobial compounds and their locus of action. This will affect how the biofilm develops and its resultant structure, leading to pockets of more sheltered bacteria within the biofilm which could continue to proliferate.
Bacterial interactions are also key in the resultant biofilm structure and subsequent resilience to antimicrobial treatment. When Veillonella parvula and Streptococcus mutans were grown as a dual-species biofilm, specific spatial arrangements were observed, which included the formation of clusters that formed within distances of 1.2 μm after treatment with chlorhexidine (Kara et al., 2007). As single-species biofilms, they were more susceptible to treatment with chlorhexidine. The authors proposed that this was a protective survival strategy against subsequent treatment with antimicrobials, similar to that observed from a biofilm consortium treated with a chlorinated alkaline solution, where one species conferred protection on another at its own expense (Leriche et al., 2003). This altruistic-like behavior of bacteria in biofilms has been proposed as a means of survival for the biofilm as a whole (Gilbert et al., 2002; Kreft, 2004). The degree to which altruistic behavior exists in dental plaque is not yet known.
NEW-GENERATION THERAPEUTICS
In a recent article that discussed host-bacterial co-evolution in relation to finding new drug targets, Zaneveld et al.(2008) stated that we have a ‘meta-genome’ composed of microbial and human components, and a ‘meta-metabolome’ that reflects metabolic activities carried out by both our microbial and human cells; and that this has implications for the definition of health, discernment of disease susceptibilities, and diagnosis of human pathologies (Zaneveld et al., 2008), as discussed above. Zaneveld et al.(2008) suggested that there is the potential for a new generation of therapeutics to be developed which affect the structure and function of our indigenous microbial communities.
Teams of investigators (Lujan et al., 2007; Potts et al., 2008) have been looking at ways of interrupting horizontal gene transfer in Escherichia coli. Their approach is based on inhibiting one of the key enzymes essential to the conjugative transfer of plasmid DNA (DNA relaxase), which has the potential to inhibit selectively the transfer of antibiotic resistance, and thus limit the generation of multidrug-resistant strains (Lujan et al., 2007). Although the impact of horizontal gene transfer within the oral microbiome has only recently been investigated, the oral cavity is believed to be an excellent environment in which horizontal gene transfer can occur (Roberts and Mullany, 2006). This has been attributed to the close and stable proximity of bacteria present in dental plaque, and the availability of exogenous DNA passing through the oral cavity (Roberts and Mullany, 2006).
Another therapeutic approach has aimed at optimizing the action of antimicrobial peptides, such as human beta-defensin-2 (hBD-2), psoriasin (PSO), and ribonuclease 7 (RNase 7), which play important roles in innate immunity (Eberhard et al., 2008). The hypothesis was that epithelial cells show a differential gene expression pattern of antimicrobial peptides (hBD-2, PSO, RNase 7) and inflammatory mediators such as interleukin-8 (IL-8) and 5-lipoxygenase (5-LO) in response to different stages of naturally formed biofilms. The findings showed that the expression of the messenger RNAs of hBD-2, RNase 7, and 5-LO was up-regulated as a result of the exposure to early biofilm stages, whereas the gene expression of IL-8 was increased in response to matured biofilms. Inter-individual differences in innate immune responses were also observed.
CONCLUDING REMARKS
Advances in molecular techniques have given rise to a much greater understanding of the diversity and complexity of human microbiota communities, including dental plaque. Plaque development may no longer be thought of in generic terms, but rather, as a highly individualized process. With that come new avenues for the understanding of plaque development in relation to its host, and therefore the potential for the development of more effective treatments. Oral diseases were once thought of as relatively simple diseases, with the etiological agents identified and treatments developed accordingly. This review has highlighted that the microbial ecology of dental plaque and the diseases it causes are far from simple, and far from being understood. As such, a new and exciting era of oral microbial ecology research awaits.
The role of microbial aggregates in plaque development. ( A conceptual representation of the potential role that microbial aggregates in saliva may play in plaque dispersal and proliferation. Microbial aggregates would detach (D) from the plaque (P), the surface-associated phase, into saliva, the planktonic-associated phase, and re-attach to pre-established plaque (EP) or establish new plaque (NP) elsewhere. Cells in the microbial aggregate would have to rely on a non-directional, fluid-driven dispersal, but would be in the protective confines of a microbial aggregate. Potentially, the detached microbial aggregates could have physiologies different from those of the original plaque as an adaptive response to a changing environment and/or diseased state, and would introduce these adaptations into the new or other established plaque. Single-cell (SC) detachment from plaque could take the form of swarming and would have the advantage over non-directional detachment, since the bacteria are self-propelled and directional.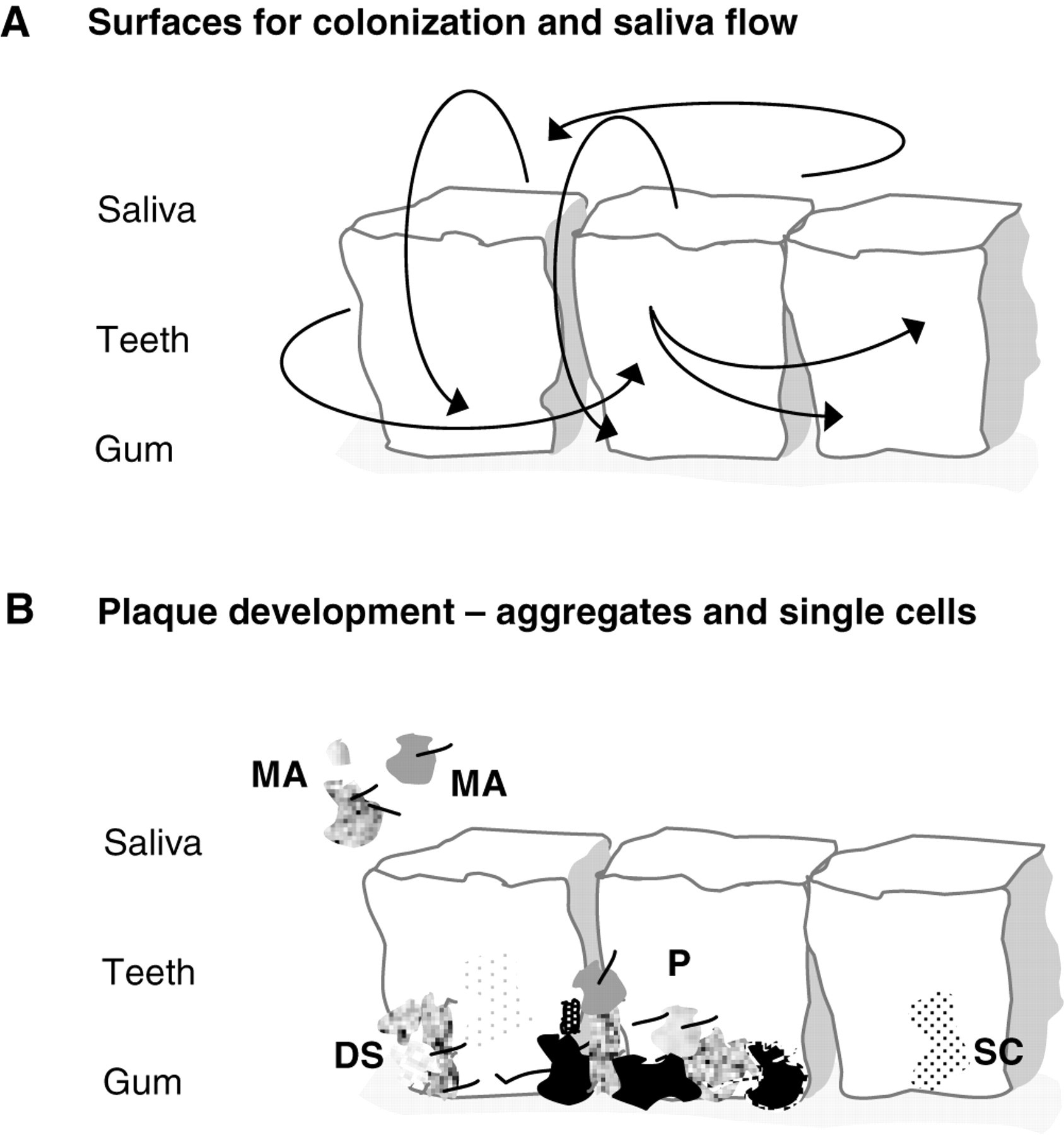
Footnotes
Acknowledgements
This manuscript is dedicated to the late Professor Peter Gilbert, who has made significant contributions to advancing our knowledge of biofilms and their recalcitrance to treatment, and who will be greatly missed. Funding for the research has been provided by the Health Research Council of New Zealand, the Wellington Medical Research Foundation, Lottery Health Research, and the University of Otago, New Zealand.