
Abstract
Since fibrosis is observed in smokers’ gingiva, it was hypothesized that fibrosis was caused by nicotine in the periodontium. Therefore, in this study, we investigated the effects of nicotine on the induction of a profibrotic molecule, connective tissue growth factor (CCN2/CTGF), in human gingival fibroblasts (HGFs) and periodontal ligament (PDL) cells. With 1 μg/mL nicotine, vacuolization and attenuated proliferation were observed. Interestingly, 1 μg/mL nicotine increased the production of CCN2/CTGF protein in both cells without increasing mRNA expression. Furthermore, type I collagen mRNA and protein were also increased and were significantly blocked by a CCN2/CTGF neutralizing antibody. This is the first report to describe a relationship between nicotine and CCN2/CTGF in periodontal tissue cells. Analysis of our data also indicated that nicotine was cytotoxic, while it increased CCN2/CTGF and, eventually, type I collagen production. These findings suggest that periodontal fibrosis can be promoted by nicotine from smoking via effects on CCN2/CTGF.
INTRODUCTION
Cigarettes, cigars, and some of their components, such as nicotine, have been found to have adverse effects on many types of cells, including gingival fibroblasts and cells of the immune system. In fact, smoking is a major risk factor for periodontitis (Calsina et al., 2002). Nicotine enters the bloodstream from the capillary vessels through lung epithelial cells, and is also promptly absorbed from oral mucosa and the skin surface during smoking. Direct effects of nicotine on the airway mucous membrane have been reported to cause chronic complications associated with smoking (Vial, 1986). Nicotine provokes inflammation and promotes scar formation in the recovery from damage to the bronchial wall (Wanner, 1985). The cytotoxic substance included in cigarettes also causes an inflammatory reaction, resulting in the migration of various cells and the secretion of cytokines and other factors (Kamma et al., 2004; Makino et al., 2008).
Connective tissue growth factor (CCN2/CTGF) is a multifunctional factor with a molecular weight of 38–40 kDa; it belongs to the Cyr61, CTGF, and Nov (CCN) family (Brigstock et al., 2003). Depending on the cell type studied, CCN2/CTGF exhibits diverse cellular functions, including extracellular matrix production (Takigawa et al., 2005), normal growth and development of certain tissues (Kubota and Takigawa, 2007), tissue regeneration, and cell migration and adhesion (Perbal, 2004). It has been shown that CCN2/CTGF is related to systemic scleroderma (SSc) (Leask, 2004), fibrosis (Ito et al., 1998), and scar formation (Blalock et al., 2003). Fibrosis is defined by the overgrowth, healing, and/or scarring of various tissues and is attributed to excess deposition of extracellular matrix components (Wynn, 2008). Among them, type I collagen is known to be a major molecule found in fibrotic organs (Alexakis et al., 2006). Clinically, it has been widely recognized that fibrosis is frequently observed in the smoker’s gingiva. However, the relationship between smoking-associated fibrosis and CCN2/CTGF has not been investigated. In this study, we investigated the relationship among nicotine, CCN2/CTGF, and gingival fibrosis by using human gingival fibroblasts (HGFs) and periodontal ligament (PDL) cells in vitro. In addition, we evaluated the hypothesis that nicotine promotes periodontal fibrosis by CCN2/CTGF-mediated induction of type I collagen.
MATERIALS & METHODS
Cell Culture and Nicotine
HGFs were obtained from human normal gingiva during extraction of impacted third molars (n = 4, one male and three female donors, 23–34 yrs of age). PDL cells were obtained from normal human periodontal tissue attached to permanent lower premolar teeth that were extracted during orthodontic therapy (n = 3, three female donors, different from those donating HGF, 23–34 yrs of age). Cells from each donor were independently cultured and analyzed. This study was approved by the Institutional Ethical Review Committee of The Nippon Dental University. Informed consent was obtained from each person prior to the study. All donors were healthy adults who had not used antibiotics and were non-smokers. Tissues were obtained according to established procedures (Sato et al., 2005). HGFs and PDL cells were cultured with 20% FBS DMEM/F-12 (Invitrogen Corp., Carlsbad, CA, USA). Cells were used between the 4th and 8th passages for all experiments. Nicotine stock solution was prepared at 10 μg/mL in 0.5% FBS DMEM/F-12 with a nicotine purity of > 97% (#148-01212; Wako, Osaka, Japan).
Reverse-transcriptase Polymerase Chain-reaction (RT-PCR) Analysis
HGFs and PDL cells were plated at 2 × 105 cells per 100-mm tissue culture plates containing 0.5% FBS DMEM/F-12. Cells were maintained in serum-deprived conditions for 24 hrs, followed by treatment with 0.5% FBS DMEM/F-12 added with nicotine at a final concentration of 0, 0.1, 1, or 10 μg/mL for 12, 24, and 48 hrs. Total RNA was isolated, and the concentration and purity of RNA were determined by spectrophotometric absorption at wavelengths of 260/280 nm. The expression of CCN2/CTGF and type I collagen mRNA transcripts was semi-quantitatively analyzed by RT-PCR (annealing temperature: 59°C for CCN2/CTGF, 56°C for type I collagen), and was standardized to the expression level of GAPDH housekeeping gene transcripts. RT-PCR was performed with Ready-To-Go PCR beads (Amersham Biosciences, Buckinghamshire, UK). The primers used are shown in the Table. PCR products were separated on a 2% agarose gel. After visualization by ethidium bromide staining, the luminescence density of each band was calculated with a densitograph lane analyzer.
Enzyme-linked Immunosorbent Assay (ELISA)
Cell culture supernatants of HGFs and PDL cells were sampled at indicated timepoints after the addition of nicotine. At the same time, the cell lysates were prepared with a lysis buffer (Epitomics, Inc., Burlingame, CA, USA). In a few experiments, a neutralizing mouse anti-CTGF antibody (Ikawa et al., 2008) was also added during nicotine treatment at a final concentration of 50 μg/mL. The levels of CCN2/CTGF and type I collagen in the cell culture supernatants or cell lysates were evaluated with sandwich ELISA systems (CCN2/CTGF provided by Nichirei Corporation, Tokyo, Japan; type I collagen purchased from ACBio, Kanagawa, Japan). Signals were developed with tetramethylbenzidine (TMB) and quantified as optical densities at a wavelength of 450 nm, as described previously (Takeuchi et al., 2008).
Immunofluorescence
HGFs and PDL cells were plated at 1 × 103 cells per well in 8-well culture chamber slides and incubated at 37°C in humidified air containing 5% CO2 until subconfluence. Following incubation under each condition, cells were washed 3 times in Dulbecco’s phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde for 30 min. Thereafter, the cells were blocked with 10% normal goat serum for 30 min. The cells were incubated for 30 min with an anti-human CTGF antibody (SC25440, Santa Cruz Biotechnologies, Santa Cruz, CA, USA) diluted at 1:200, followed by incubation with Alexa Fluor 488 goat anti-rabbit IgG (H+L) (A11008, Invitrogen Corp.) at 1:200 for 2 hrs. After being washed 3 times with 0.01M PBS (−), cells were incubated with 4,6-diamidino-2-phenylindole (DAPI) (Cambrex Science, Walkersville, MD, USA) and mounted with Vectashield (Vector Laboratories, Inc., Burlingame, CA, USA), and then observed by epifluorescent microscopy. Cells treated without primary antibody were used as negative controls.
Cell Growth Assay and Morphological Analysis
Both cell types were plated at a density of 1×106 cells each in 35-mm dishes. The cells were observed by phase-contrast microscopy with an inverted microscope. After observation and image capture, the cells were detached by trypsinization, and viable cells were counted by the dye-exclusion test with a 0.4% trypan blue solution. These analyses were performed in triplicate.
Statistical Analysis
All data are presented as mean ± SD. Analysis of variance and Fisher’s Protected Least Significant Difference were used for the evaluation of significant differences. Unless otherwise specified by brackets, statistical significance was evaluated between each group with nicotine at the specified dose and a corresponding control group without nicotine. A p value < 0.05 was considered significant. Significance was evaluated with STATVIEW (Hulinks, Inc., Tokyo, Japan) software. All experiments were repeated at least twice, yielding comparable results.
RESULTS
Effect of Nicotine on the Gene Expression and Production of CCN2/CTGF in Human Periodontal Tissue Cells
Initially, we evaluated the effect of nicotine at the mRNA level. As a result, expression of CCN2/CTGF mRNA in HGFs was significantly increased by 0.1 μg/mL nicotine from 24 to 48 hrs during the treatment (p < 0.05). In contrast, PDL cells showed significant decreases in CCN2/CTGF mRNA levels at 12 and 48 hrs (p < 0.01). At 1 and 10 μg/mL, nicotine decreased CCN2/CTGF gene expression in both cells (p < 0.05). In particular, almost no CCN2/CTGF mRNA could be detected with 10 μg/mL nicotine (Figs. 1A, 1B).
However, surprisingly, the levels of CCN2/CTGF protein in cell lysates were significantly increased after 12 or 24 hrs of treatment by 1 μg/mL nicotine, both in HGFs (p < 0.05) and in PDL cells (not statistically significant). In both cell lysates, the peak of the protein accumulation occurred at 12 hrs, and subsequently, the protein levels showed a constant decrease over time, probably due to secretion. In supernatants after 12 to 48 hrs of nicotine treatment, CCN2/CTGF concentrations were significantly increased in both cell types (p < 0.05). In contrast to that in cell lysates, the peak of the protein level occurred at 24 hrs in the supernatants (Figs. 1C, 1D).
Consistent with the above findings, immunofluorescence analysis revealed no change in CCN2/CTGF protein levels with 0.1 μg/mL of nicotine (data not shown), but indicated higher protein levels at 1 μg/mL of nicotine in both cell types than in controls, which was most evident after 12 hrs of treatment (Appendix Fig. 1).
Effect of Nicotine on Type I Collagen in Human Periodontal Tissue
Next, we investigated whether nicotine could promote type I collagen deposition through the induction of CCN2/CTGF. Expression of type I collagen mRNA was significantly increased time-dependently by 0.1 and 1 μg/mL nicotine treatment in HGFs (p < 0.05) and PDL cells (p < 0.01), compared with controls. With 10 μg/mL nicotine, gene expression could be detected in both cell types, which indicated the cytotoxic effect of nicotine at this dose (Figs. 2A, 2B). Furthermore, the levels of type I collagen protein were significantly increased in cell fractions of both cells (p < 0.05) after nicotine treatment at 1 μg/mL for 24 hrs (Figs. 2C, 2D), which was effectively blocked by an anti-CCN2/CTGF neutralizing antibody (Figs. 2E, 2F).
Effect of Nicotine on the Growth and Morphology of the Cells
To assess the cytotoxic effects, we monitored cell growth in the presence or absence of 0.1–10 μg/mL of nicotine. Viable cell numbers were significantly increased in the presence of 0.1 μg/mL of nicotine in both cells (p < 0.05).
However, cell numbers significantly decreased compared with controls (p < 0.05) on the addition of 1 μg/mL nicotine. At a dose of 10 μg/mL, almost no viable cells were observed (Figs. 3A, 3B).
By microscopic morphological analysis, vacuolization was evidently observed in 1 μg/mL nicotine- treated HGFs, with obvious morphological changes (Appendix Fig. 2). With 10 μg/mL nicotine, only almost-desquamated cells were observed (Fig. 3C).
DISCUSSION
This is the first study to clarify that nicotine promotes CCN2/CTGF production from HGFs and PDL cells, which eventually induces the production of type I collagen, a major component of the fibrotic extracellular matrix. In clinical cases, gingival fibrosis is often observed among smokers. However, no report has shown a distinct relation between the thickening/fibrosis of periodontal tissue and habitual smoking. Nicotine associated with smoking can cause many complications throughout the body, whereas CCN2/CTGF is a key player in establishing fibrotic legions in various organs (Abdel Wahab and Mason, 2004; Gressner and Gressner, 2008). Therefore, our present results provide a novel insight into the pathogenesis of periodontal disorders in smokers. Generally, nicotine is regarded as a highly toxic molecule, as observed here after 12 hrs of stimulation. Disturbance in cell orientation with vacuolization of the cells was observed with 1 μg/mL nicotine. At 10 μg/mL, most of the cells died and were detached, presenting a spherical appearance. Under such conditions, the number of cells was significantly decreased by nicotine in a dose- and time-dependent manner. Expression of CCN2/CTGF mRNA was also significantly decreased at 1 or 10 μg/mL; however, intriguing results were obtained in terms of CCN2/CTGF production. In cell lysates, the production levels of CCN2/CTGF protein were significantly increased in HGFs and PDL cells by 1 μg/mL of nicotine within 12 hrs of the treatment. After 12 hrs of treatment, the cell-associated CCN2/CTGF protein was decreased in a time-dependent manner in both cell types, whereas accumulated CCN2/CTGF was increased in both supernatants after 24 hrs of treatment. Taken together, these results indicate that CCN2/CTGF overproduced within 12 hrs of nicotine treatment was mostly secreted in to the culture supernatant by 24 hrs after the beginning of the treatment. Overall, the levels of production of CCN2/CTGF protein were significantly increased compared with those in control groups for all times evaluated in both cell lysates and supernatants of either cell type. Subsequently, we examined the effect of nicotine on type I collagen gene expression and production, which has been reported to have a positive relationship to CCN2/CTGF (Heng et al., 2006). We found that expression of type I collagen mRNA was significantly induced by nicotine at 24 and 48 hrs in HGFs and over the time evaluated in PDL cells. Moreover, the levels of type I collagen protein were significantly increased by 1 μg/mL nicotine in the lysates of both cells after 24 or 48 hrs of treatment. Importantly, this effect was neutralized by a CCN2/CTGF neutralizing antibody, indicating that it was indeed mediated by CCN2/CTGF. Analysis of our data, taken together, indicates that 1 μg/mL of nicotine was cytotoxic. It prominently increased CCN2/CTGF protein levels without a significant increase in mRNA, which yielded an increase in type I collagen gene expression. The observed increase of CCN2/CTGF protein without an increase in corresponding mRNA suggests a translational regulation of CCN2/CTGF gene expression by nicotine, the mechanism of which remains to be clarified.
CCN2/CTGF promotes extracellular matrix production, and it has been reported that CCN2/CTGF is strongly associated with fibrosis in general. A relationship between CCN2/CTGF and fibrosis has been reported in several organs (Cheng et al., 2006), including the lung (Lasky et al., 1998). It should be noted that the lungs and oral cavity are the only organs that have direct contact with smoke during smoking, and that nicotine can be absorbed from the mucosa. Once the lungs become accustomed to smoke, stimulation of inflammation is continually repeated, which eventually leads to chronic irreversible lung damage. In such circumstances, it has been reported that a large amount of collagen causes a decrease in the contraction force, and such pathological changes cannot be reversed. Of note, involvement of CCN2/CTGF in pulmonary fibrosis has been strongly indicated as well. Compared with the lungs, the oral cavity is exposed to much higher concentrations of nicotine during smoking. Therefore, it is strongly suspected that induction of a profibrotic molecule, CCN2/CTGF, by nicotine is a major promoter of periodontal fibrosis caused by smoking. However, until this study, the effect of continual stimulation with nicotine for a long period, which mimics the events occurring in smokers’ oral cavity, had not been assessed. Animal studies are needed to confirm our findings in vivo. Subsequent clinical research may clarify the nature of these findings and may develop a novel clinical application.
Primer Sequences Used for RT-PCR
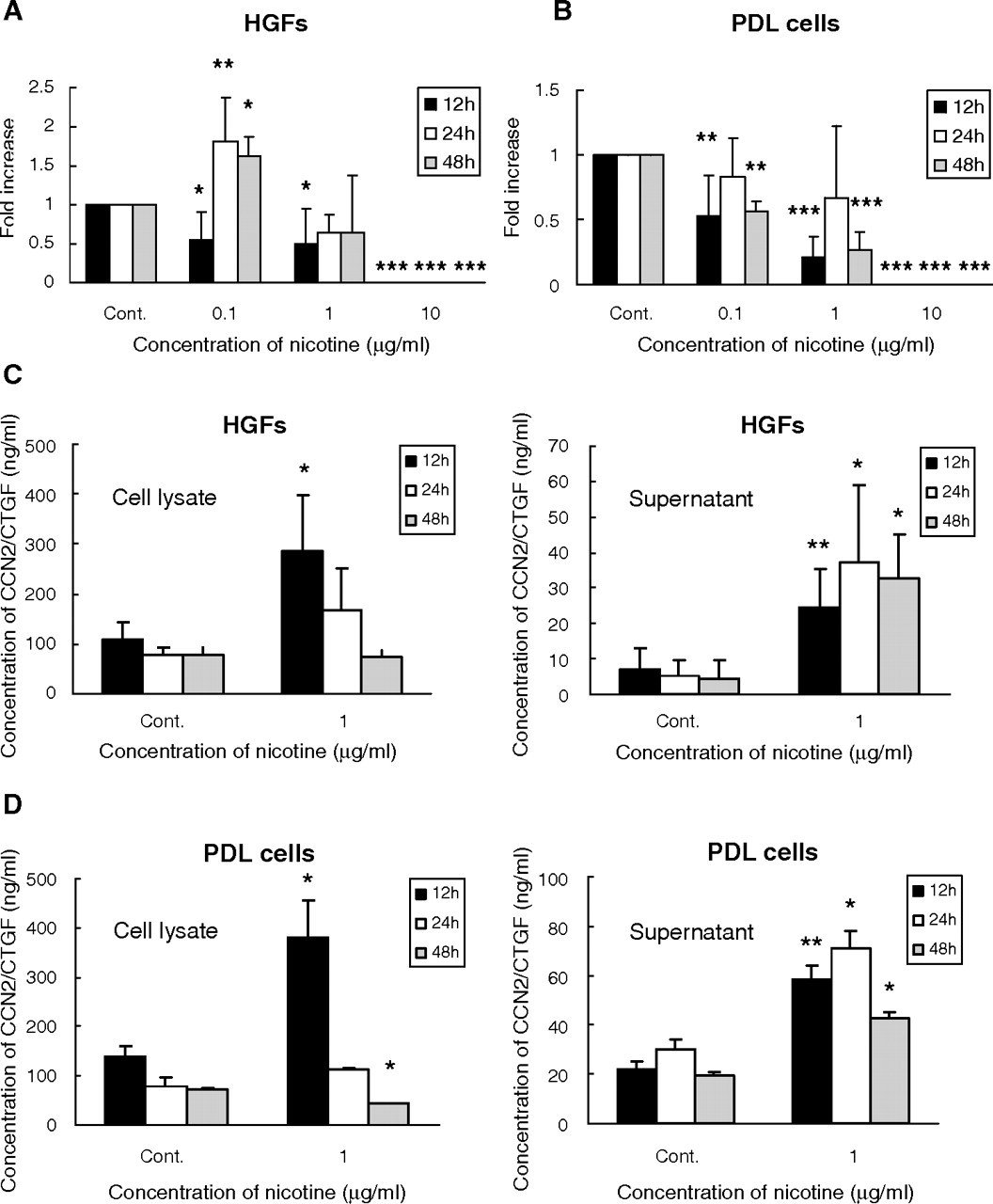
Quantitative analysis of connective tissue growth factor (CCN2/CTGF) mRNA induced by nicotine in human gingival fibroblasts (HGFs)
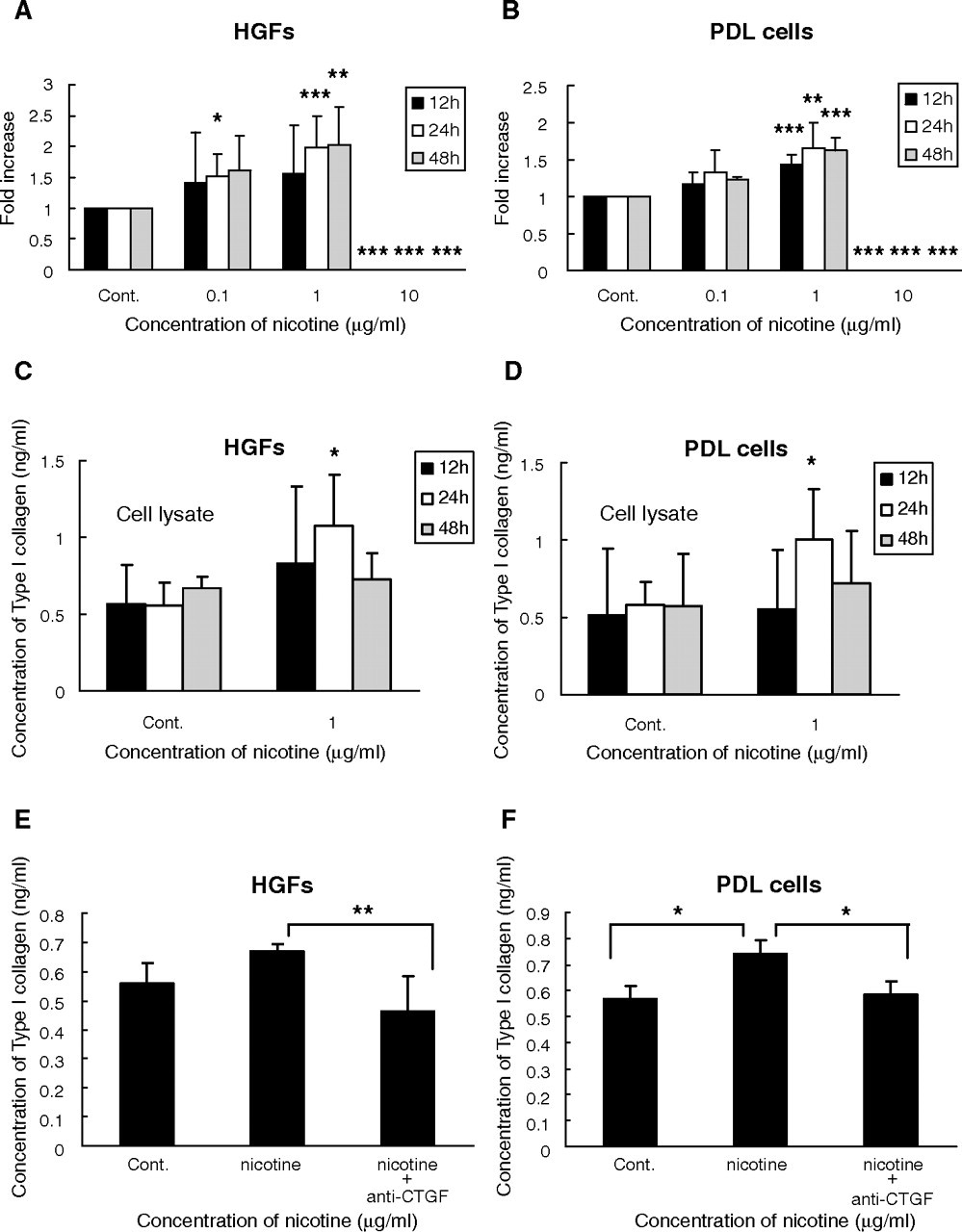
Quantitative analysis of type I collagen mRNA induced by nicotine in human gingival fibroblasts (HGFs)
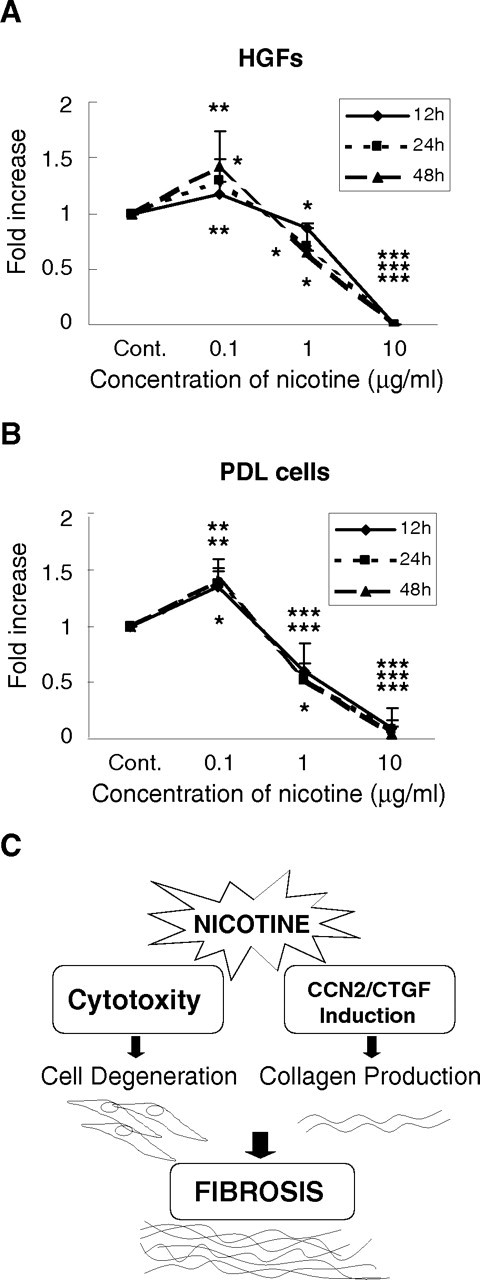
Effect of nicotine on cell growth. Relative cell numbers were counted and computed against those of corresponding controls at each timepoint. Addition of 0.1 μg/mL of nicotine enhanced cell growth, whereas nicotine of higher doses repressed it in both cell types.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.