
Abstract
Individuals receiving clozapine treatment for schizophrenia complain of drooling. Reports on salivary flow measurements are contradictory in humans and lacking in animals. Clozapine has affinity for several different receptor types and may, hypothetically, both stimulate and inhibit salivary secretion. In rats, intravenous clozapine evoked a long-lasting secretion, being more prominent from submandibular than from parotid glands. Chronic denervation enhanced the responses. Clozapine acted on muscarinic (M1-) receptors of acinar cells, independent of central nervous mechanisms, pre-synaptic intraglandular events, or circulating catecholamines. A fraction of the methacholine- and parasympathetic-nerve-evoked secretion was abolished by clozapine at doses below those evoking secretion. Sympathetic-nerve-evoked secretion was partially reduced by clozapine, due to antagonistic action on α-adrenoceptors; the β-adrenoceptor-mediated response persisted. Subsecretory doses of clozapine enhanced secretion induced by the β-adrenoceptor agonist isoprenaline. The overall actions of clozapine suggest that, in clozapine-treated humans, salivation is increased during sleep and at rest, but is decreased during meals.
INTRODUCTION
Clozapine is used to treat schizophrenic patients resistant to therapy with traditional antipsychotics. About one-third of the clozapine-treated individuals complain of drooling during the daytime and of a wet pillow following a night’s sleep (McEvoy et al., 2006; Praharaj et al., 2006). The situation can be so troublesome that the clozapine treatment is discontinued. Objective measurements of salivary flow in humans are few and, in fact, inconclusive (Ben-Aryeh et al., 1996; Rabinowitz et al., 1996; Sockalingam et al., 2007). Animal experiments directly recording salivary secretion are lacking. Indeed, some investigators suggest that salivation in response to clozapine reflects an inhibitory effect by the drug on the swallowing reflex, rather than a stimulatory effect on the salivary glands (McCarthy and Terkelsen, 1994; Pearlman, 1994).
The present study was designed to define the influence of clozapine on salivary secretion, with the rat parotid and submandibular glands as models. Both glands are supplied with muscarinic acetylcholine receptors, α1- and β1-adrenergic receptors, and non-adrenergic, non-cholinergic receptors, which, upon stimulation, evoke the secretion of saliva to various degrees (Ekström, 1999a). A mixed population of muscarinic M3/M1-receptor subtypes mediates salivation (Tobin et al., 2009). Assays on transfected cell lines show the antagonist profile of clozapine to dominate on various muscarinic receptor subtypes, even though it exerts a weak agonist activity on subtypes M1, M2, and M4; in addition, clozapine is antagonistic to the α1-adrenergic receptor (Weiner et al., 2004; Lameh et al., 2007). Due to the mixed properties of clozapine, both excitatory and inhibitory secretory effects were expected. Therefore, we recorded the effects of clozapine on non-secreting as well as on secreting glands.
MATERIALS & METHODS
Animal procedures were as humane as possible and were approved by the Ethics Committee for Animal Experiments, Göteborg, Sweden. Fifty-seven adult female Sprague-Dawley rats (Charles River, Sulzfeld, Germany), each weighing 284 ± 8 g (mean ± SEM), were used. For the acute experiment, the animals were anesthetized with pentobarbitone (55 mg/kg, i.p., followed by supplementary intravenous doses as required). The body temperature of the anesthetized animal, measured with a rectal probe, was maintained at 38°C by means of a thermostatically controlled blanket. Eventually, the animals, still under anesthesia, were killed by exsanguination, and the parotid and submandibular glands were removed, cleaned, and weighed, the weight (of non-denervated glands) being 187 ± 7 mg (n = 34) and 190 ± 6 mg (n = 35), respectively.
The animals were fitted with a femoral venous polyethylene catheter, which served as a conduit for injections, and a tracheal cannula. The parotid duct was exposed externally, close to the mouth and the submandibular duct under the mylohyoid muscle. In the same rat, only one gland of each type was examined, except for those rats subjected to chronic denervation, where also the contralateral gland was examined. The ducts were cannulated with fine polyethylene tubes. The parasympathetic auriculo-temporal nerve and the sympathetic trunk were exposed and cut at the base of the skull and in the neck, respectively. The peripheral end of each nerve was placed in a bipolar ring electrode. The submandibular gland was chosen for studies on sympathetic secretion, since the response is greater from this gland than from the parotids.
We performed preliminary surgery 2–4 wks before the terminal experiment under pentobarbitone anesthesia (25 mg/kg, i.p.) combined with ketamine (50 mg/kg, i.m.), to cause chronic gland denervation. Combined parasympathetic and sympathetic post-ganglionic denervation of the parotid glands (7 rats) was achieved by avulsion of the auriculo-temporal nerve (Khosravani et al., 2006) and the superior cervical ganglion (Alm and Ekström, 1977), respectively. We deprived the submandibular glands of parasympathetic pre-ganglionic innervation by cutting the chorda-lingual nerve (5 rats). Surgery was performed unilaterally; the contralateral gland served as control.
The nerves were electrically stimulated (6 V, 2 ms) using a Grass S48 stimulator and Grass SIU 5A isolation unit (Grass Technologies, Astr-Med, Inc., West Warwick, RI, USA). Clozapine (30 mg/mL or 10 mg/mL) was dissolved in ethanol, dissolved in 80–70% (v/v) aqueous ethanol, and then diluted further in saline to reach lower concentrations. The vehicle, in various ethanol concentrations, had no effect on secretion. When the intravenous effect of clozapine per se was tested, the interval between subsecretory doses was usually 10–15 min. Due to a drug half-life of about 1½ hrs, a dose accumulation was likely to have occurred (Baldessarini et al., 1993; Sun and Lau, 2000). At secretory doses, the interval was extended until the secretion disappeared or reached a steady level. When the effect of clozapine was tested on methacholine- or nerve-evoked responses, the interval between two doses was usually 15–20 min.
Since clozapine induced salivary secretion, at the higher doses tested, pre-existing secretion was a complicating factor when the effect of clozapine on methacholine- or nerve-evoked secretion was to be determined. The immediate preceding response was subtracted from the response subsequent to the respective mode of stimulation. Comparisons were based on 5-(methacholine injection), 2- (sympathetic stimulation), and 1-minute periods (parasympathetic stimulation).
The secreted saliva was collected in pre-weighed vials or on pre-weighed filter paper to enable the flow to be estimated gravimetrically (assuming the density of saliva to be 1.0 g/mL). To monitor mean aortic blood pressure, we inserted a cannula, connected to a pressure transducer, into the femoral artery.
Drugs
Atropine sulphate, isoprenaline hydrochloride, methacholine chloride, pirenzepine hydrochloride (“M1-selective antagonist”), propranolol hydrochloride, and substance P were obtained from Sigma Chemicals (St. Louis, MO, USA). Clozapine and phentolamine mesylate were obtained from Novartis Pharma AG (Basel, Switzerland). The drugs were administered intravenously.
Statistics
The statistical significance of differences was calculated either by Student’s t test for paired values or by one-way analysis of variance (ANOVA), followed by Fisher’s protected least-significant difference based on raw data. P < 0.05 was considered significant. Values are means ± SEM.
RESULTS
Effects of Clozapine per se in Normal and Denervated Glands
There was no resting secretion. All submandibular glands secreted at a dose of 3 mg/kg (Fig. 1). Saliva appeared at 1–2 min upon clozapine administration. The flow peaked within 5–15 min and, depending on dose, continued at a fairly steady rate for 1–2 hrs. Combined α- and β-adrenoceptor blockade (by phentolamine and propranolol, respectively) was without effect on clozapine-induced secretion (before, 0.67 ± 0.03 μL/5 min; after, 0.60 ± 0.06 μL/5 min, n = 4). Few parotid glands responded to the highest dose of clozapine (30 mg/kg). Combined chronic sympathetic and parasympathetic post-ganglionic denervation of the parotid gland and chronic parasympathetic pre-ganglionic denervation of the submandibular gland sensitized the secretory cells of the atrophied glands, not only to methacholine, but also to clozapine. Pirenzepine completely abolished the ongoing secretion to clozapine, 3 mg/kg, from non-operated submandibular glands (0.37 ± 0.08 μL/5 min, n = 7; mean dose of pirenzepine 0.19 ± 0.05 mg/kg, n = 7) and completely or almost completely abolished the secretion to 10 mg/kg (0.66 ± 0.07 μL/5 min vs. 0.06 ± 0.03 μL/5 min, n = 7; mean dose of pirenzepine 0.3 ± 0.03 mg/kg, n = 7). The ongoing secretion from the denervated submandibular (5 ± 1 μL/5 min, n = 7) and parotid glands (0.8 ± 0.2 μL/5 min, n = 5) upon clozapine administration, 3 mg/kg, was unaffected by propranolol (0.5 mg/kg), but was completely abolished by pirenzepine (mean dose 0.3 ± 0.1 mg/kg, n = 7, and 0.15 ± 0.05 mg/kg, n = 5, respectively). Atropine (0.2 mg/kg) completely abolished the responses.
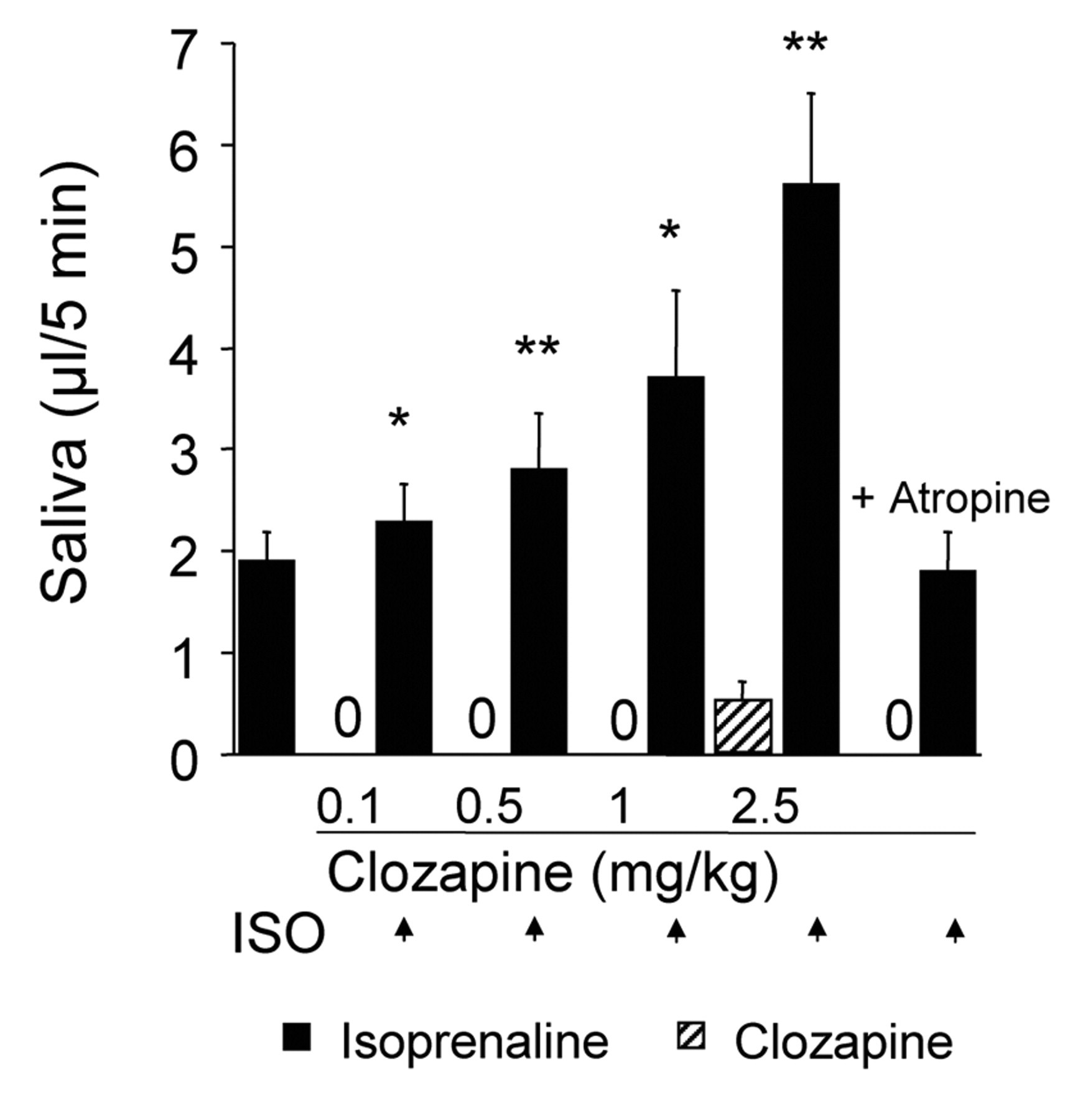
Synergistic Interactions
Both sub- and suprathreshold doses of clozapine enhanced the submandibular flow of saliva evoked by a standard dose of the β-adrenoceptor agonist isoprenaline (Fig. 2). Atropine (0.2 mg/kg) abolished the positive interaction.
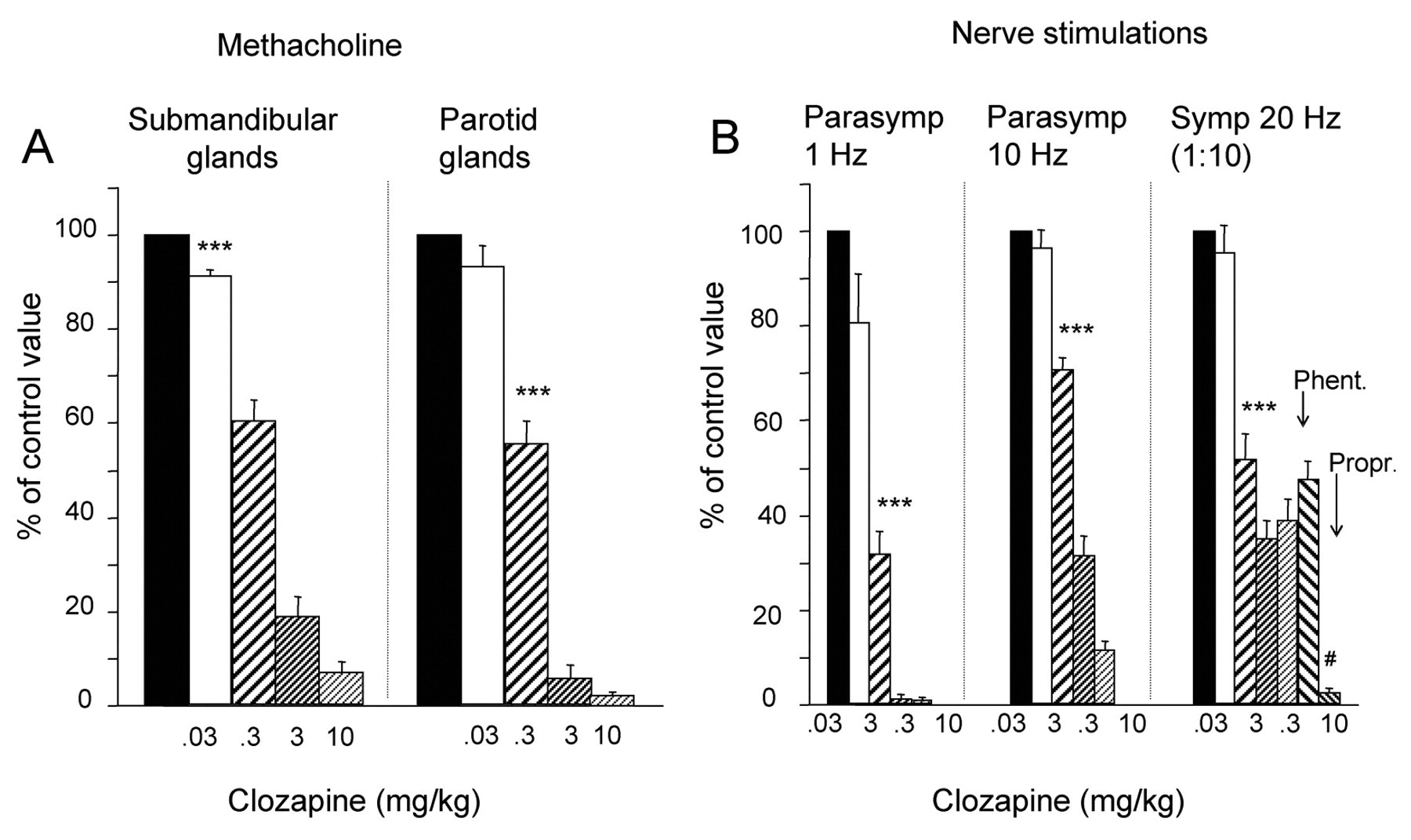
Effects on the Methacholine-evoked Response
After clozapine, 0.3 mg/kg, the responses of the 2 glands were reduced by 40–44% (Fig. 3). Upon a dose of 10 mg/kg, the persisting response was 7–2%, which disappeared after pirenzepine (0.2–0.5 mg/kg). Secretion could still be elicited, however, since, when tested in the submandibular glands (and after atropine, 1–2 mg/kg), the glands responded (27 ± 5 μL, n = 5) to substance P (0.5 μg/kg) with a volume in the same range as in the absence of clozapine and atropine (32 ± 6 μL, n = 5).
Effects on the Parasympathetic-nerve-evoked Response
Clozapine, 0.3 mg/kg, reduced the parotid response by 68% (1 Hz) and 29% (10 Hz) (Fig. 3). The persisting response (12%, 10 Hz) in the presence of clozapine, 10 mg/kg, was abolished by atropine (0.2 mg/kg).
In the rats treated with clozapine and subjected to atropine (1–2 mg/kg), stimulation at 10 Hz for a period longer than the standard protocol of 1 min invariably evoked a small flow of parotid saliva starting about 30 sec later. The “delayed” secretion is attributed to non-adrenergic, non-cholinergic transmission mechanisms (Ekström, 1999a).
Effects on the Sympathetic-nerve-evoked Response
The response was halved by 0.3 mg/kg of clozapine (Fig. 3). A steady secretory level was reached at 3 mg/kg, the reduction being 65%. In the presence of clozapine, 10 mg/kg, phentolamine (2 mg/kg) caused no further reduction. A subsequent injection of propranolol (1 mg/kg) almost abolished the response. In 3 rats not subjected to clozapine, phentolamine (2 mg/kg) reduced the sympathetic secretion (at 20 Hz) by 76 ± 3 % (P < 0.05).
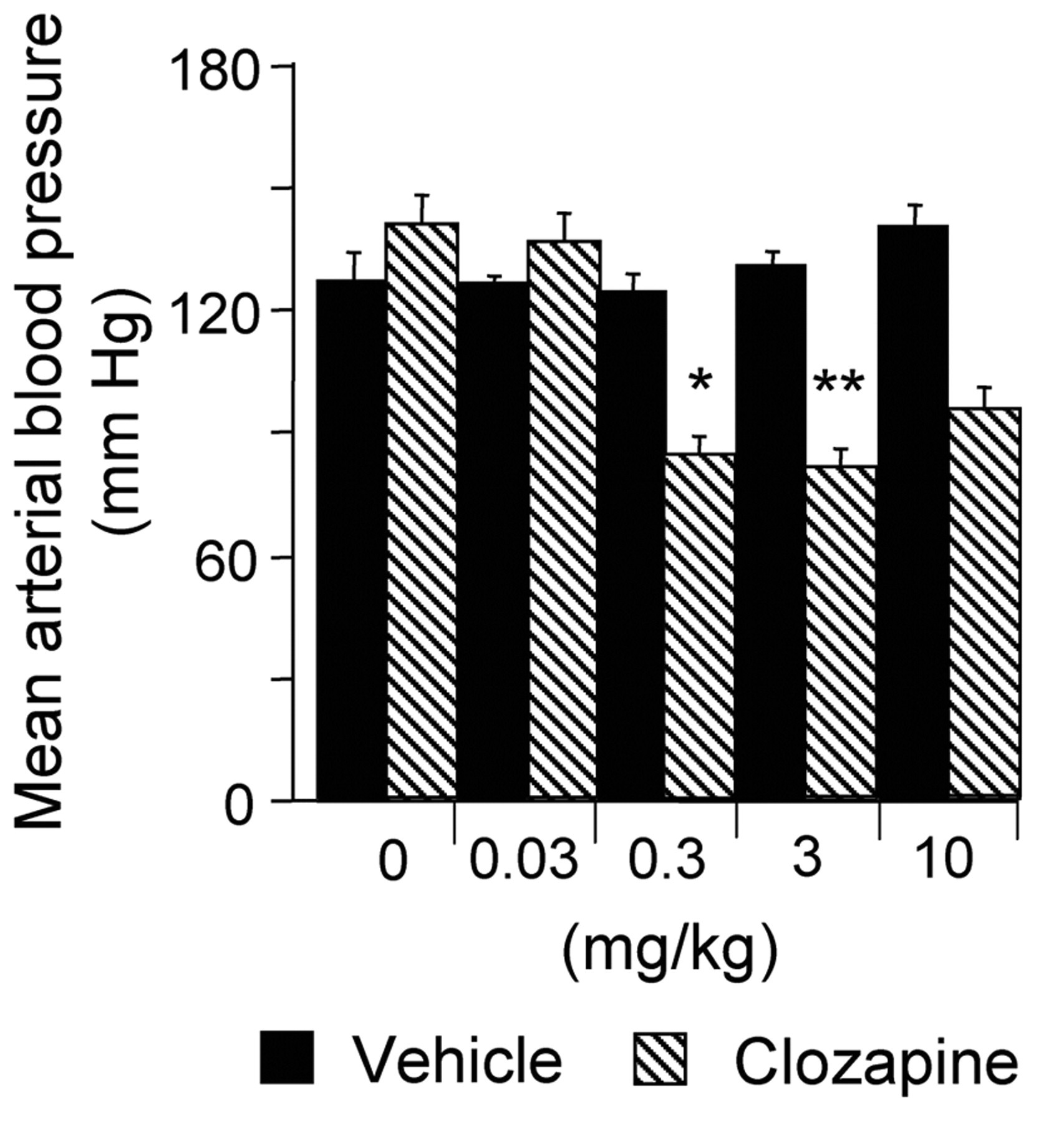
Blood Pressure
Vehicles were without effect. Increasing doses of clozapine gradually lowered the blood pressure (Fig. 4).
DISCUSSION
The present animal model supports clinical reports on clozapine-induced sialorrhea (McEvoy et al., 2006; Praharaj et al., 2006). Clozapine caused secretion by a direct action on muscarinic (M1-) receptors of the acinar cells, a response independent of central nervous mechanisms, pre-synaptic intraglandular events, or circulating catecholamines. The larger responses to clozapine from submandibular than from parotid glands agree with previous findings of an acinar immunoreactivity for M1-receptors that is stronger in submandibular than in parotid glands (Ryberg et al., 2008).
No support for a secretagogue effect of clozapine was found in a study on isolated rat submandibular acinar cells, with intracellular Ca2+ to indicate secretion (Pochet et al., 2003). However, human submandibular gland tissue exposed to clozapine in vitro responded with ultrastructural changes reflecting secretory activity (Riva et al., 2003; Testa Riva et al., 2006).
Interestingly, the present study indicates two mechanisms that may enlarge the volume of saliva secreted in the presence of clozapine under clinical conditions: positive interactions, as shown for isoprenaline, and denervation supersensitivity. In salivary acinar cells, muscarinic receptor agonists mobilize the Ca2+-pathway, while β-adrenoceptor agonists mobilize the cAMP-pathway. Synergistic interactions between agonists using the two pathways occur with respect to secretion, e.g., involving the parasympathetic cotransmitter vasoactive intestinal peptide, activating the cAMP-signaling mechanism (Ekström, 1999a). Since non-adrenergic, non-cholinergic transmitter mechanisms may be preserved in the presence of clozapine, clozapine may interact synergistically not only with cAMP-activating sympathetic noradrenaline and circulating adrenaline, but also with non-adrenergic, non-cholinergic mechanisms. Supersensitivity to secretagogues in salivary glands following surgical or pharmacological (pre-/post-ganglionic) denervation, reducing or abolishing the transmitter bombardment of the gland receptors, is a well-known phenomenon (Emmelin, 1961; Ekström, 1980; Ekström and Malmberg, 1981). In the denervated glands, clozapine showed increased excitatory efficacy. It is suggested that, in humans under clozapine treatment, prolonged blockade of the non-M1-receptor fraction of the muscarinic receptor population by clozapine, combined with a blockade of α1-adrenoceptors (see below), increases the sensitivity of the non-blocked muscarinic receptor population to agonistic actions, and thus enlarges the secretory response mediated by these non-blocked receptors (Ekström, 1999b). That clozapine exerted strong inhibitory actions in ongoing secretion was shown in the present study, and this occurred at dose levels of clozapine below those evoking overt secretion to clozapine alone.
The methacholine-evoked secretion from submandibular and parotid glands was almost abolished by clozapine, as was the secretion from parotid glands upon parasympathetic stimulation, provided the stimulation period was short, thus avoiding an overt secretory contribution from non-adrenergic, non-cholinergic transmitter mechanisms (Ekström, 1999a). Likewise, ultrastructural changes evoked by carbachol in human submandibular tissue, indicating secretory activity, were partially reduced by clozapine (Testa Riva et al., 2006).
The submandibular secretion to sympathetic stimulation was more than halved by clozapine. The finding that the α-adrenoceptor antagonist phentolamine diminished the sympathetic response in the absence of clozapine, but not in its presence, suggests that, in the clozapine-treated rats, the α1-adrenoceptors of the gland were already under blockade. In contrast, the β-adrenoceptor antagonist propranolol inhibited the sympathetic-nerve-evoked response under clozapine, suggesting that clozapine lacked antagonistic action on the β1-adrenoceptors of the gland, an observation in agreement with unaffected isoprenaline responses of atropinized rats in the presence of clozapine.
Clozapine lowered arterial blood pressure, which may be ascribed to loss of sympathetic vasoconstrictor tone mediated by α1-adrenoceptors (Lameh et al., 2007). A decrease in pressure of the present magnitude is unlikely to influence the methacholine-or nerve-evoked secretory responses presently under study (Lung, 1990), since they were of short duration.
Clozapine is subjected to extensive hepatic metabolism (Eiermann et al., 1997). N-desmethylclozapine, a major metabolite, has been suggested to be responsible for the positive results in response to clozapine treatment of schizophrenia (Lameh et al., 2007). The rapid response to clozapine presently found favors the opinion that clozapine exerts the observed pharmacodynamic effects primarily in a non-metabolized form.
Though comparisons should be made with caution, it may be mentioned that the clozapine doses presently tested were in a dose range clinically used (Taylor et al., 2005). The experimental model suggests that, in humans, the contribution of clozapine-induced secretion becomes prominent when superimposed upon the small spontaneous secretion during sleep or between meals (in addition, maintained by a weak reflex; Hector and Linden, 1999), explaining “the wet pillow” in the morning and the drooling during the daytime. It also suggests that the volume produced in response to a meal or to citric acid (eliciting strong reflexes) will probably be reduced. Moreover, the demonstration of a clozapine agonistic action on M1-muscarinic glandular receptors and of a clozapine synergistic interaction with agonists acting on β1-adrenergic glandular receptors may open new perspectives in the search for a therapeutic agent able to control clozapine’s side-effects on salivation.
Secretion from normal and chronically denervated glands. Synergistic interactions in submandibular glands. Isoprenaline (2 μg/kg) was given alone or repeatedly (as indicated by arrows), 10 min after the administration of sub- and suprathreshold intravenous secretory doses of clozapine. On its own, the isoprenaline-evoked secretory response was completed within 3 min. The salivary collection periods were 5 min in all tests. Atropine (0.2 mg/kg) abolished not only the clozapine-induced secretion, but also the enhancement of the isoprenaline-evoked response. Three rats were used. Columns represent mean values, and vertical bars are ± SEM. *P < 0.05 and **P < 0.01 compared with the isoprenaline-evoked response alone or with the calculated additive secretory response to isoprenaline and clozapine given separately. Effects of clozapine on Effects of clozapine (n = 3) at various doses and the corresponding vehicle (n = 3) on mean arterial blood pressure integrated over 5 min. Columns represent mean values, and vertical bars are ± SEM. Comparisons were made between pre- and post-administration values. *P < 0.05 and **P < 0.001.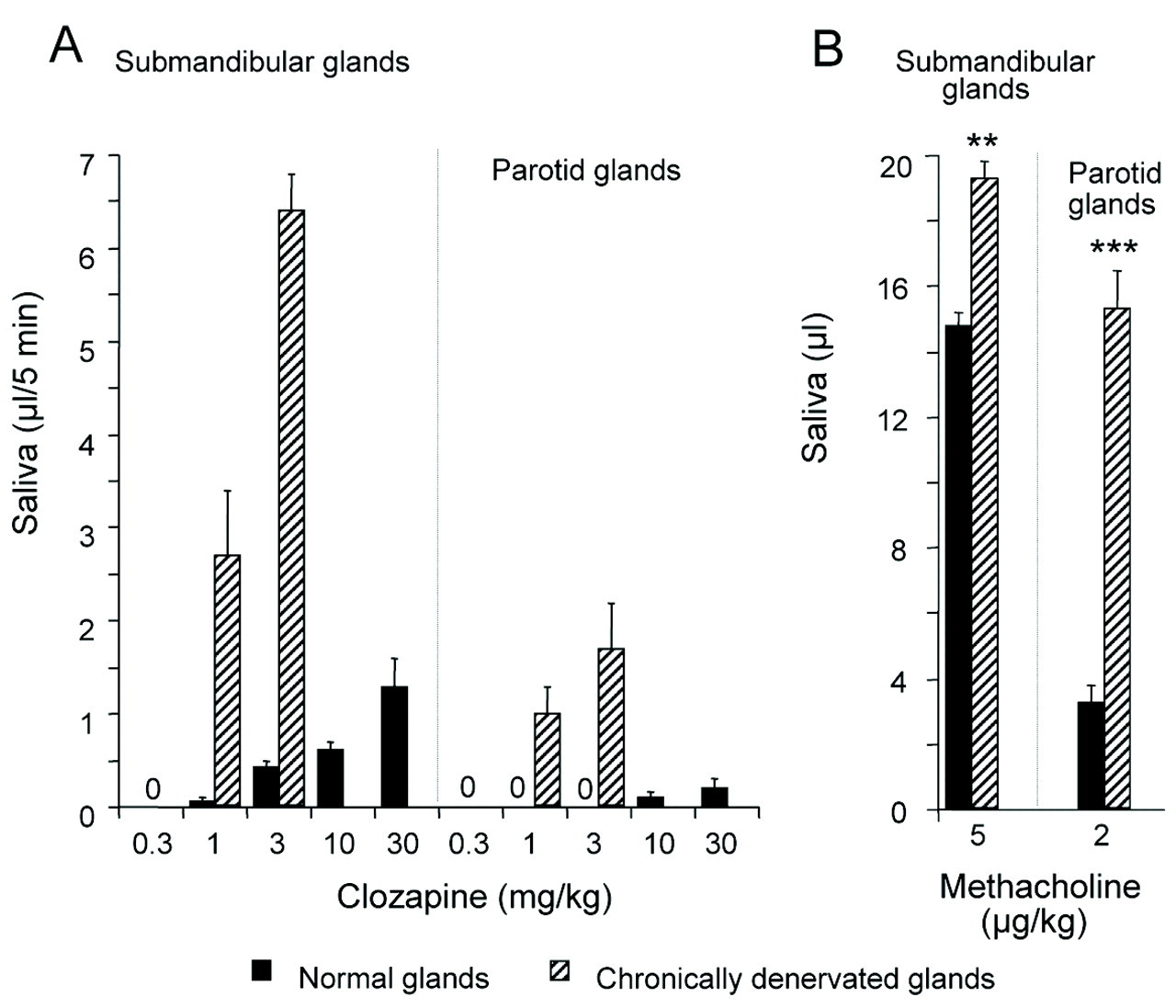
Footnotes
Acknowledgements
This study was supported by the Swedish Science Council (05927), the LUA/ALF agreement (ALFGBG-11907), the Swedish Dental Society, the Ministero Istruzione Università Ricerca PRIN, and the “INTERLINK” agreement (II04C00H8M). J. Ekström acknowledges support from the University of Cagliari for periods as a “Visiting Professor”. Clozapine was a gift from Novartis. We thank Mrs. Ann-Christine Reinhold for her excellent assistance. This paper is based on an abstract presented at the 2009 General Session of the International Association for Dental Research (Miami, FL, USA) [Ekström J, Godoy T, Riva A. Mixed Actions of Clozapine on Salivary Secretion. J Dent Res 88(Spec Iss A):767, ![]() ].
].