
Abstract
Selective interaction of mutans streptococci with benign bacteria could present an opportunity for their removal from the mouth without disruption of other oral flora. This study was conducted to find probiotic lactobacilli that could specifically co-aggregate in vitro with mutans streptococci, but not with other plaque commensals. A search of 624 lactobacilli among a large culture library revealed 6 strains, all classifiable as L. paracasei or L. rhamnosus, which met this criterion. Such novel, specific co-aggregation, however, was not a general characteristic of these species or the genus Lactobacillus. The co-aggregation by these specific lactobacilli was characterized as heat treatment (autoclaving)- and protease-resistant, lectin-independent, not inactivated by sugar substitutes, operational over a wide pH range, unaffected by whole saliva, but calcium-dependent. It is thus seen to present a potential strategy for in vivo alteration of plaque biofilm and caries.
INTRODUCTION
Caries is strongly associated with colonization of the tooth-surface plaque biofilm by mutans streptococci; most common among humans being Streptococcus mutans (Loesche, 1986; Tanzer et al., 2001). The ability of mutans streptococci to adhere persistently to teeth is required for promoting lesion development. They increase in the plaque as a result of growth and recruitment from the planktonic phase (saliva) by auto-aggregation upon exposure to dietary sucrose (Kuramitsu, 1993; LeBlanc, 1994).
Bacterial interactions with mutans streptococci, masking their tooth-/plaque-binding sites, could be used to promote their selective removal from oral biofilms. While mutans streptococci use teeth as primary colonization sites, they are shed into saliva during oral function. They are, nonetheless, a relatively small proportion of the salivary planktonic flora (Dahlén, 1984). From suspension in saliva, mutans streptococci can either re-adhere to the teeth or existing plaque (Gibbons et al., 1986; Schilling and Bowen, 1992) or be cleared by swallowing (Russell et al., 1999). Thus, specific aggregation of planktonic mutans streptococci could provide a strategy to reduce caries by fostering oral clearance, promoting health without adverse effects on other oral biofilm members.
This study was conducted to find probiotic lactobacilli that could specifically co-aggregate in vitro with mutans streptococci, but not with other prominent supragingival oral organisms, and to characterize them in vitro. In a subsequent paper, we shall describe the utility of such bacteria in vivo.
MATERIALS & METHODS
Micro-organisms and Cultivation
Although the literature does not report prominent co-aggregation phenomena with lactobacilli (Kolenbrander, 1988), we sought lactobacilli that co-aggregate mutans streptococci among our (ORGANOBALANCE GmbH, Berlin, Germany) culture collection. Out of the 624 strains of lactobacilli that were grown in MRS medium (De Man et al., 1960) at 37°C, Lactobacillus paracasei DSMZ16671 became the focus of study. L. paracasei strains are naturally found in food products (Siragusa et al., 2007).
Mutans streptococci were grown at 37°C in trypticase soy broth (Difco BD, Heidelberg, Germany) supplemented with 3 g/L yeast extract (Ohly, Hamburg, Germany), termed TSY. Several reference strains were studied (Table), as were S. mutans wild-types isolated from mixed saliva of eight healthy adults, after informed consent was obtained, on selective medium (Schaeken et al., 1986). One skin commensal was also studied.
The taxonomic identification of the strains to species level relied on phenotypic characterization by the API 50 CH system and APILAB PLUS software v3.3.3 (bioMerieux, Craponne, France). Bacterial counts were determined from calibration curves of optical density vs. microscopic cell counts in a Petroff-Hausser chamber.
Chemicals and Enzymes
EDTA, sugars, sugar substitutes, and inorganic chemicals were reagent grade (Merck, Darmstadt, Germany; Carl Roth, Karlsruhe, Germany), and proteases (pronase E, proteinase K, trypsin, and chymotrypsin) were of the highest commercially available grade (Sigma, Taufkirchen, Germany).
Screening for Co-aggregates
Stationary-phase cells were washed by centrifugation and re-suspended, to OD600nm 10 for lactobacilli and 4 for non-lactobacilli, at room temperature in PBS (pH 7.0), then used for auto-aggregation and co-aggregation assays in 2 mL volumes. Ratios of potential co-aggregants/lactobacilli ranged from 1/1 to 20/1 (cell/cell) in various experiments. Mixtures were vortexed and incubated at 25°C, and immediate flocculation was observed. If after 20 min none had occurred, pairs were judged to be non-co-aggregating. For some experiments, cells were stained separately with hexidium iodide (HI, 10 μg/mL) or carboxyfluorescein diacetate succinimidyl ester (CFDA-SE 1 μg/mL) (Invitrogen, Carlsbad, CA, USA) according to manufacturer’s instructions, and centrifuged. Un-reacted fluorescent dyes were removed by extensive washing with PBS. Cells were mixed in equal proportions and vortexed 1 min prior to phase-contrast and fluorescence microscopy.
Sensitivity of Co-aggregation
We prepared serial dilutions of L. paracasei DSMZ16671 in PBS or saliva and incubated them with S. mutans DSMZ20523 to determine the sensitivity of co-aggregation.
Scanning Electron Microscopy (SEM)
Cells were prepared as described above and re-suspended in PBS. Co-aggregation was induced by mixing suspensions of DSMZ16671 and S. mutans DSMZ20523 in a ratio of 1/1 (cell/cell). The resulting co-aggregates were pelleted by centrifugation, re-suspended in PBS, vortexed for 2 min, re-centrifuged (350 × g,10 sec, Heraeus Multifuge 3S-R centrifuge; Kendro, Langenselbold, Germany), then fixed in 2.5 % glutaraldehyde, dehydrated in graded ethanol solutions, dried in liquid CO2, and sputter-coated with palladium before SEM. Some images were colorized according to bacillary or coccal cell shape to facilitate viewing.
Sugar, pH, and Ca++ Effects on Co-aggregation
Co-aggregation was tested both after 30-minute incubation of mutans streptococci with 50 mM sucrose, followed by washing with PBS, and simultaneously with incubation of known co-aggregating pairs. We performed analogous simultaneous incubations, testing lactose, mannose, rhamnose, isomaltulose, isomaltitol, trichlorogalactosucrose, mannitol, xylitol, or sorbitol to detect possible interference with the ability of DSMZ16671 to co-aggregate mutans streptococci. For evaluation of the pH-dependency of co-aggregation, pure cultures of DSMZ16671 and mutans streptococci were grown to stationary phase, and harvested by centrifugation at 4°C. The cells were re-suspended in 0.1 M acetate buffer adjusted to pH values from 4.0–7.0 and in Sorensen’s phosphate buffer from pH 6.0–8.0, in 0.5 pH unit intervals. Co-aggregation was then assessed in a ratio of 1/3 (lactobacilli/streptococci). No pure cultures evidenced auto-aggregation within this pH range.
To determine the dependency on Ca++ for auto-aggregation and co-aggregation, we washed DSMZ16671 cells twice in 200 mM EDTA. For the removal of EDTA, they were repeatedly washed in water. Co-aggregation was then assayed, as above. Then cells were re-suspended in 0.2, 0.5, or 2.0 mM CaCl2 or in 100 mM MgCl2, and co-aggregation was assayed again.
Protease Pre-treatment
The susceptibility to protease inactivation of the co-aggregation determinants on the surfaces of DSMZ16671 and of mutans streptococcal strains was tested after treatment with 1 of 4 proteases—pronase E (4 U/mg), proteinase K (46 U/mg), trypsin (12.4 U/mg), or chymotrypsin (58.3 U/mg)—as follows. Lactobacilli and streptococci were grown separately to stationary phase, harvested by centrifugation, washed in PBS, and 1-mL aliquots adjusted to OD = 2 (for lactobacilli) or OD = 4 (for mutans streptococci) in PBS (pH 7.0) containing individual proteases at a final concentration of 2.5 mg/mL. After incubation for 1 hr at 37°C, cells were washed and re-suspended again in PBS (pH 7.0). Then, 50-μL aliquots of each of the various potentially co-aggregating pairs were mixed, and co-aggregation was assayed visually and microscopically, as detailed above.
Saliva Effects
Two-mL aliquots of various stationary-phase pure cultures were harvested and re-suspended in 2 mL of human whole saliva. Whole saliva, individually collected after 2–3 hrs of fasting and induced by the chewing of sugar-free sorbitol gum, was used immediately after collection. Thus, both auto-aggregation and co-aggregation were assayed in salivary suspensions, as detailed above. Whole saliva of healthy adults was collected and used following a previously approved study plan, after informed consent of the participants was obtained.
Heat Treatment
Pure cultures of stationary-phase L. paracasei DSMZ16671, washed and re-suspended in PBS, were autoclaved at 121°C for 20 min. The bacteria, untreated or heat-treated, were mixed, and co-aggregation was assayed as described above.
Binding of L. paracasei DSMZ16671 to Hydroxyapatite
CFDA-SE-stained cells of either DSMZ16671 or S. mutans DSMZ20523 were incubated with hydroxyapatite (BASF SE, Ludwigshafen, Germany) suspensions at concentrations between 2 and 0.01% (w/v). Binding was monitored microscopically.
RESULTS
Screening for Lactobacilli that Co-aggregate Specifically with Mutans Streptococci
Among 624 Lactobacillus strains in a proprietary collection, only 6 bound (co-aggregated with) S. mutans DSMZ20523 (Fig.1). Four were identified as Lactobacillus paracasei subsp. paracasei, 2 were Lactobacillus rhamnosus, and all were deposited at the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany). Numerous L. paracasei and L. rhamnosus strains did not co-aggregate with S. mutans. The lactobacilli did not auto-aggregate, and the streptococci neither auto-aggregated nor aggregated in spent culture supernatants of lactobacilli (Fig. 1). Strain DSMZ16671 had the highest co-aggregating activity and was investigated further, but the behavior of all co-aggregating lactobacilli was essentially the same.
Phase-contrast and brightfield images of co-aggregates of cells stained with HI or CFDE-SE, then allowed to co-aggregate, demonstrated that both DSMZ16671 and S. mutans participated in the aggregation, but did not auto-aggregate (Fig. 2). Both live and autoclaved L. paracasei DSMZ16671 induced co-aggregation with S. mutans DSMZ20523 (Fig. 2). SEM images of co-aggregants are presented in Fig. 2. Clearly, single cells of L. paracasei DSMZ16671 bound several S. mutans cells, resulting in a cross-linking of the co-aggregates; and the binding sites on DSMZ16671 appeared evenly distributed over its surface.
Failure of L. paracasei DSMZ16671 to Co-aggregate with Common Non-mutans Members of the Oral Flora, and to Adhere to Hydroxyapatite
None of the non-mutans oral commensals detectably co-aggregated with L. paracasei DSMZ16671 (Table) or its spent culture filtrate, and no auto-aggregation was observed. Notably, no co-aggregation was induced by DSMZ16671 with A. naeslundii I and II or with S. oralis, S. sanguinis, S. mitis, and S. gordonii strains, all implicated in tooth surface biofilm participation (Kolenbrander et al., 2002). Co-aggregation was, however, observed with 2 Staphylococcus epidermidis strains typical of skin microflora, but not prominent in the mouth (Table).
In contrast to mutans streptococci, which had high affinity for hydroxyapatite, L. paracasei DMSZ16671 did not bind to hydroxyapatite (data not shown).
Sensitivity of Co-aggregate Formation
With a microtiter plate titration method, quantitative co-aggregation potency of L. paracasei DSMZ16671 against mutans streptococci can be illustrated (Fig. 3). Strain DSMZ16671 induced visible sediment formation with DSMZ20523 at cell/cell ratios from 1/2.5 up to at least 1/20 (Fig. 3), and, in larger-scale tests, to 1/56. Autoclaved lactobacilli induced similar sediment formation with the approximate same sensitivity (Fig. 2).
Effects of Sugars on Co-aggregation
Although incubation of mutans streptococci with sucrose causes them to auto-aggregate (Freedman and Tanzer, 1974; Kuramitsu, 1993), it was still possible to observe persistent interaction in this setting when S. mutans and L. paracasei DSMZ16671 were fluorescence-labeled with different dyes prior to co-incubation. Both lactobacilli and streptococci were present in the apparent co-aggregates formed in the presence of sucrose, independent of pre-incubation of S. mutans with sucrose and subsequent extensive washing with PBS, thus suggesting persistent exposure of L. paracasei DSMZ16671-reactive sites on S. mutans cells after being coated with cell-surface-associated glucans.
Because the interaction between cells of L. paracasei DSMZ16671 and S. mutans during co-aggregation must involve cell surfaces, we explored possible interference by other sugars and sugar alcohols. Simultaneous or previous incubation in the presence of the sugars lactose, rhamnose, or mannose, the sugar alcohols sorbitol, mannitol, or xylitol, and the artificial sweeteners trichlorogalactosucrose, isomaltulose, or isomaltitol had no effect on co-aggregate formation between S. mutans and DSMZ16671 by comparison with standard incubations that had never been exposed to them.
Effects of Protease and Heat
Incubation with 4 different proteases (Jenkinson et al., 1990) with DSMZ16671 did not alter its binding to non-treated S. mutans. However, protease treatment of S. mutans abolished co-aggregation with DSMZ16671, regardless of the protease used.
Heat treatment (autoclaving) of DSMZ16671 did not reduce its ability to co-aggregate S. mutans, as shown by studies of log2 dilutions of either live or autoclaved cells of DSMZ16671 (Figs. 2, 3). However, heat treatment of mutans streptococci abolished co-aggregation with L. paracasei DSMZ16671.
Effects of pH, Saliva, and Divalent Cations
Co-aggregation had comparable efficiency at pH values from 4.0 to 8.0. Thus, at most pH values encountered in saliva, co-aggregation of S. mutans by DSMZ16671 persists.
Co-aggregation of mutans streptococci and L. paracasei DSMZ16671 was not hindered or altered in its intensity by the presence of fresh unaltered whole human saliva.
EDTA washing of DSMZ16671 to remove divalent cations resulted in loss of ability to co-aggregate with S. mutans. Subsequent re-incubation in 0.20 and 0.5 mM CaCl2 partially and completely, respectively, restored its ability to co-aggregate S. mutans; however, the use of up to 100 mM MgCl2 instead of CaCl2 did not restore its ability to co-aggregate S. mutans. Unlike shaking or heat treatment, dissociation of existing co-aggregates did occur in the presence of 200 mM EDTA.
DISCUSSION
The specific co-aggregation of mutans streptococci by 6 L. paracasei and L. rhamnosus strains, typified L. paracasei DSMZ16671, is novel. Co-aggregation also occurred with S. epidermidis. Multiple oral commensals thought to participate in biofilm formation on the teeth (Kolenbrander, 1988; Kolenbrander et al., 2002) were not co-aggregated by these lactobacilli. Numerous other lactobacilli in a very large culture collection, including many other L. paracasei and L. rhamnosus strains, failed to manifest mutans-specific co-aggregation. Because the co-aggregation activity of these specific lactobacilli was heat-stabile and protease-resistant, and was not disrupted by lactose, rhamnose, or mannose, it was apparently lectin- and surface-protein-independent (Simões et al., 2008). It was also not affected by sugar alcohols or other sugar substitutes, but was calcium-dependent, as demonstrated by EDTA-abolition and calcium-restoration effects. This novel co-aggregation was not detectably affected by the presence of whole fresh saliva or by pH values within the range of 4.0 and 8.0, beyond the range existing in saliva. While mutans streptococci have high affinity for hydroxyapatite, L. paracasei DSMZ16671 has none.
It is reasonable to speculate that in vivo co-aggregation of mutans streptococci, masking their surface sites ordinarily available for binding to exposed teeth or mutans-rich plaque biofilm, could reduce caries risk by fostering clearance of planktonic mutans streptococci from the mouth by swallowing.
Because L. paracasei DSMZ16671 did not co-aggregate ubiquitous commensals of the plaque biofilm and other seemingly benign oral flora colonizing the tongue, e.g., S. salivarius (Teughels et al., 2007), such a strategy would seem unlikely to induce opportunistic infections or create voids in the otherwise seemingly benign supragingival and mucosal oral flora.
The heat stability of the mutans-co-aggregating trait of the probiotic strain L. paracasei DSMZ16671 suggests that it could be delivered to the mouth in a variety of vehicles. Ongoing work is exploring validation of these concepts of managing mutans streptococci in the mouth by application of strain L. paracasei DSMZ16671 via delivery vehicles such as, e.g., toothpaste, mouthwash, or confections.
Strains of the Oral Microflora and Their Co-aggregation Behavior with L. paracasei
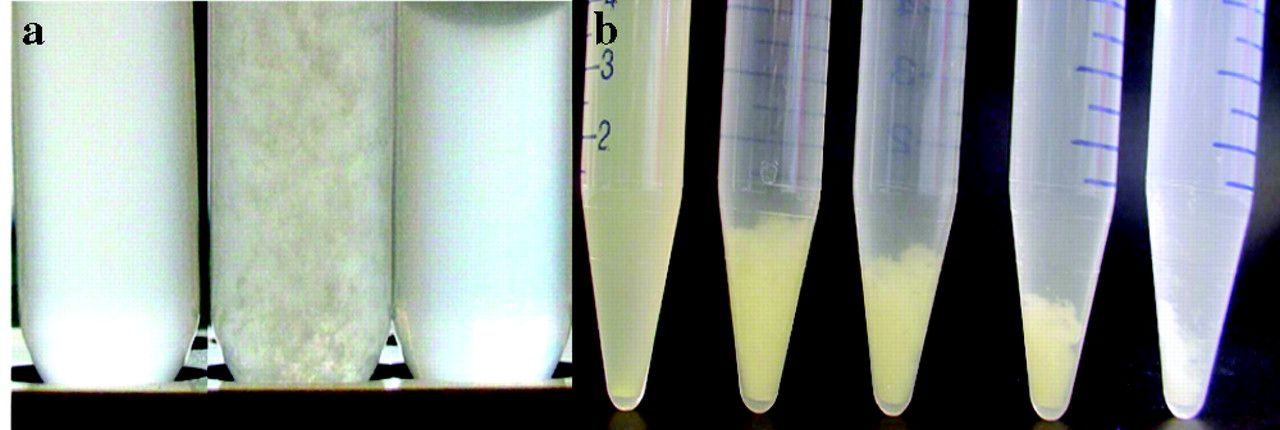
Co-aggregation of Lactobacillus paracasei DSMZ16671 with Streptococcus mutans DSMZ 20523.
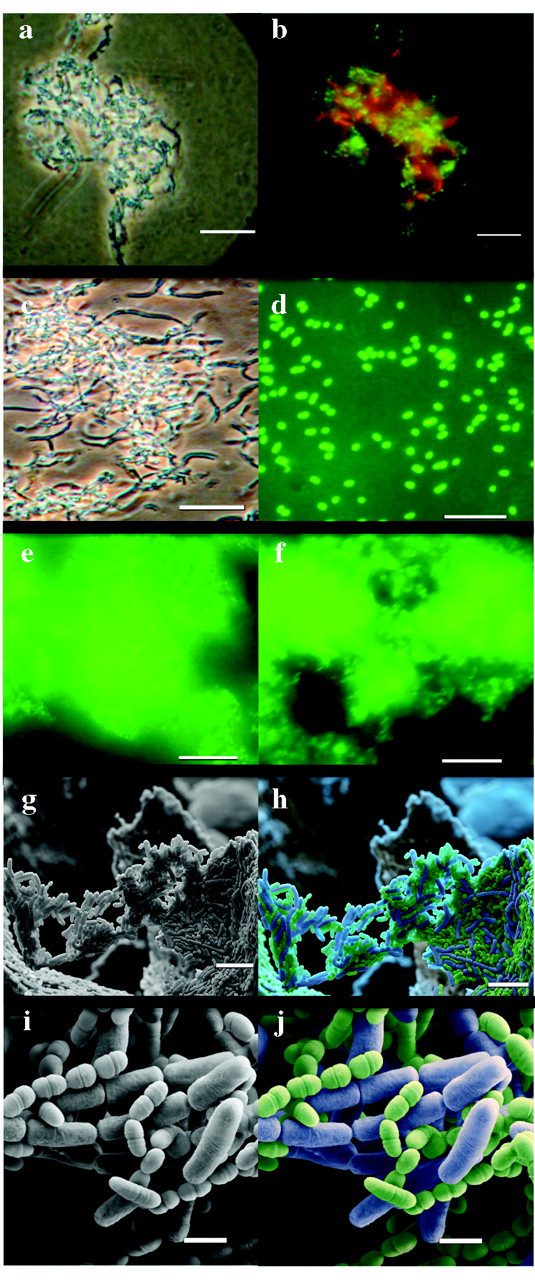
Microscopic analysis of co-aggregation of Lactobacillus paracasei DSMZ16671 with Streptococcus mutans DSMZ20523. A single co-aggregate is shown (
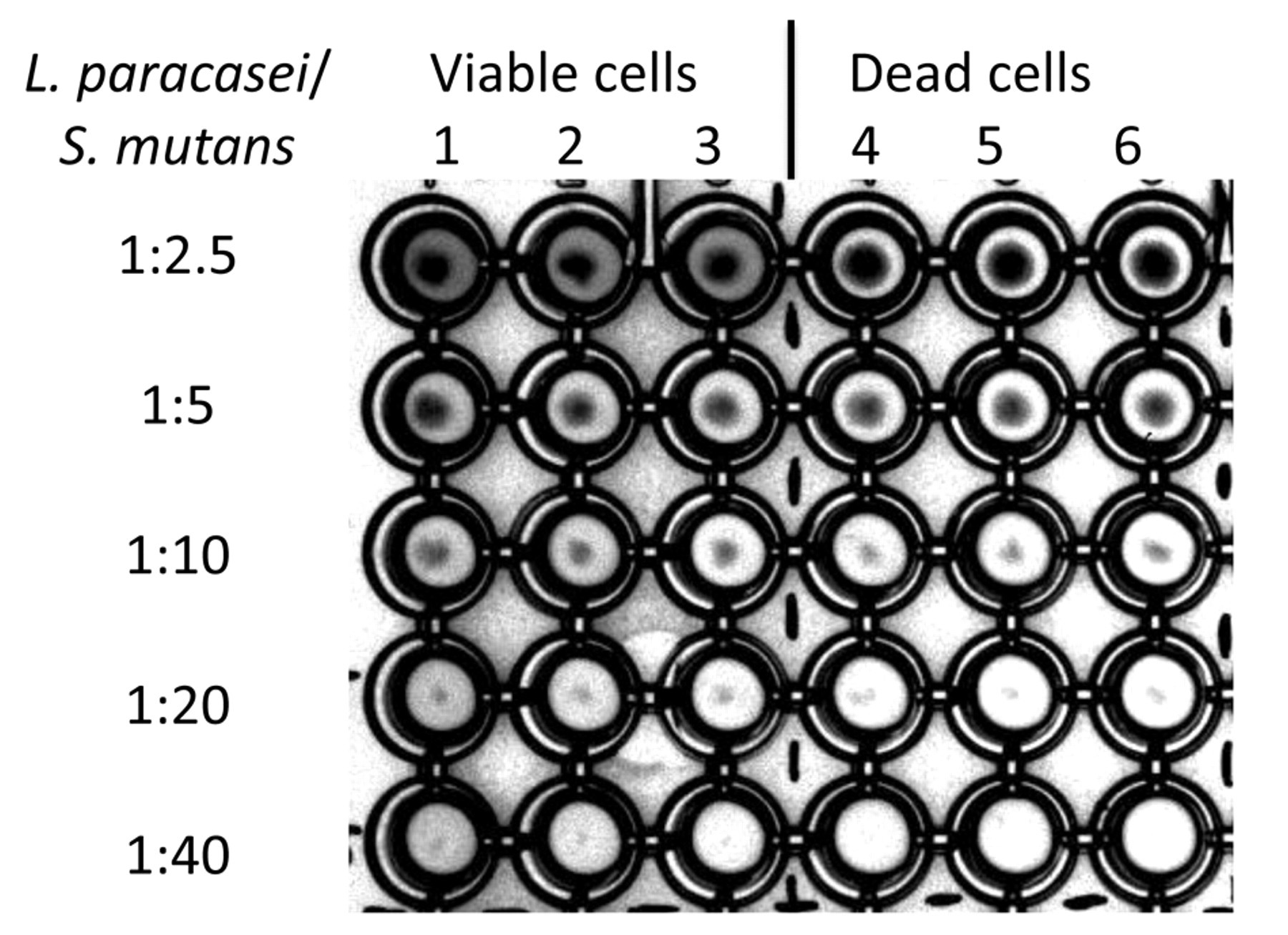
Titration of co-aggregation of L. paracasei DSMZ16671 with mutans streptococci by a microtiter plate co-aggregation assay and visual inspection. Cells were mixed at an approximate ratio of lactobacilli and S. mutans of 1 to 2.5 (row 1). L. paracasei cells were diluted in a log2 manner, while the S. mutans cell numbers were kept constant. S. mutans DSMZ20523 was added to L. paracasei DSMZ16671, and vortexed for 12 min at 1000 rpm (Shaker PSU-2T Plus, lab4you GmbH, Berlin, Germany). Resultant co-aggregates were observed as sediments in wells. Viable (columns 1, 2, and 3) and dead (columns 4, 5, and 6) lactobacilli were compared as to efficiency in co-aggregating S. mutans. Titers of 1:20 (L. paracasei/S. mutans) resulted in visible co-aggregation with pellet formation. Live cells and dead (autoclaved) cells had comparable activity.
Footnotes
Notes
Acknowledgements
The authors thank Oliver Meckes and Nicole Ottawa (eye of science, Reutlingen, Germany) for the SEM photographs. The authors acknowledge the excellent technical assistance of Christina Balcke and Peggy Jacob. This work was entirely funded by BASF Future Business GmbH. No financial considerations were paid directly to any of the co-authors except by virtue of their past or present employment by ORGANOBALANCE or BASF-FB. J.M. Tanzer was not paid in any form.