
Abstract
Although it is known that diabetes impairs oral wound healing, relatively little is known about the cellular parameters affected, particularly in connective tissue. This study investigated the hypothesis that diabetes impairs connective tissue formation in healing gingiva, and that impaired healing is associated with factors that decrease fibroblast numbers. Full-thickness wounds were created in the palatal gingiva of type 1 and type 2 diabetic and normoglycemic mice. Five days after wounding, diabetic mice had less epithelial wound coverage, less new connective tissue formation, and reduced fibroblast density (p < 0.05). This occurred with increased numbers of caspase-3- and TUNEL-positive fibroblasts, decreased fibroblast proliferation, increased nuclear translocation of the pro-apoptotic transcription factor FOXO1, and increased numbers of polymorphonuclear leukocytes, all of which were significant (p < 0.05). The results suggest that diabetes may decrease fibroblast numbers through increased apoptosis and reduced proliferation, both of which may be mediated through increased activation of FOXO1.
Introduction
Diabetes is a systemic disease that results from an insufficient level of insulin or a relative insensitivity to this hormone’s actions that in turn causes hyperglycemia. Diabetes causes several complications, including impaired wound healing (Singer and Clark, 1999). Wound healing requires the integration of complex cellular and molecular events in successive phases of inflammation, granulation tissue formation, and re-epithelialization (Singer and Clark, 1999; Clark, 2001). During the inflammatory stage, leukocytes remove debris and produce growth factors to stimulate the repair process (Barrientos et al., 2008). This is followed by the formation of granulation tissue, which invades the wound area and is characterized by the proliferation of fibroblasts and endothelial cells. Remodeling is the last and longest phase of healing, during which the tensile strength of the wound increases as a result of the structural modification of the newly deposited collagen. In diabetes, wound healing is impaired. Numerous aspects of the wound repair process are altered in diabetes, including dysfunction in the inflammatory response, reduced granulation tissue formation, and impaired angiogenesis. It has been proposed that increased fibroblast apoptosis is a mechanism for diabetes-impaired dermal wound healing (Darby et al., 1997; Liu et al., 2004).
Apoptosis is a highly regulated form of programmed cell death, defined by distinct morphological and biochemical features (Vaux and Strasser, 1996; Peter et al., 1997). It occurs when a cell is damaged, infected with a virus, or undergoes stressful conditions such as starvation or exposure to harmful conditions such as oxidative stress. During the process of wound healing, remodeling of granulation tissue is accompanied by the apoptosis of fibroblasts (Desmoulière et al., 1995). The balance between cell proliferation and apoptosis is an important factor in the determination of whether there is an increase or decrease in cell numbers. Inflammatory mediators and advanced glycation end-products are elevated in diabetic wounds and may stimulate apoptosis through mechanisms that involve activation of reactive oxygen species, the MAP kinase signaling pathway, activation of the pro-apoptotic transcription factor FOXO1, and activation of caspase-3 (Alikhani et al., 2005). FOXO1 stimulates apoptosis by regulating the expression of pro-apoptotic genes and also has other effects depending upon the cell type and tissue (Birkenkamp and Coffer, 2003; Alikhani et al., 2005). For example, FOXO1 causes cell cycle arrest and decreased proliferation (Furukawa-Hibi et al., 2005) and regulates transcription of genes involved in gluconeogenesis and adipocyte differentiation (Hiromura et al., 2007). Furthermore, FOXO1 has been implicated in the inflammatory events that contribute to diabetic retinopathy and impair bone formation (Behl et al., 2008, 2009).
In this study, we investigated enhanced fibroblast apoptosis as a potential mechanism through which diabetes affects gingival wound healing in type 1 and type 2 diabetic mice, to gain potential new insights into mechanisms through which diabetes may impair connective tissue healing in the oral cavity.
Materials & Methods
Mice
Eight-week-old male CD-1 mice were purchased from Charles River Laboratories (Wilmington, MA, USA), and type 1 diabetes was induced by multiple low-dose intraperitoneal injection of streptozotocin (STZ, 40 mg/kg; Sigma, St. Louis, MO, USA) in 10 mM citrate buffer daily for 5 days as we have previously described (Kayal et al., 2007). Control mice were treated identically with vehicle alone, 10 mM citrate buffer. Mice that spontaneously develop diabetes, C57BL/KsJ-lepr-/- (db/db) mice, and their non-diabetic littermates, C57BL/KsJ-lepr-/+ (db/db+), were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). Serum glucose levels were measured (Accu-Chek; Roche Diagnostics, Indianapolis, IN, USA), and mice were considered to be diabetic when blood glucose levels exceeded 250 mg/dL, and experiments were started when mice were hyperglycemic for a minimum of 3 wks. For both types of diabetic mice, serum glucose levels were typically 400 to 450 mg/dL and 100 to 150 mg/dL for normoglycemic controls. All animal procedures were approved by the Institutional Animal Care and Use Committee, Boston University Medical Center.
Specimens
Full-thickness excisional wounds (1.5 mm in diameter) were created in the anterior palatal gingiva close to the central incisors. The animals were killed 5 days after creation of the wounds. This time-point was determined based on previous studies and was selected since it is a time-point when healing has progressed, but one can also distinguish between normal and impaired healing (Nooh and Graves, 2003). The head of each animal was dissected free and fixed in cold 4% paraformaldehyde, followed by dissection of the palate with intact soft tissue and decalcification by incubation in cold Immunocal (Decal Corporation, Congers, NY, USA). Specimens were embedded in paraffin, and 5-µm sections were prepared. All counts and measurements were carried out under blinded conditions by one examiner and confirmed by a second independent examiner.
Histomorphometric Analysis
Histomorphometric analysis was undertaken with hematoxylin- and eosin-stained sections. The entire wound was sectioned, and those sections where the wound was widest, representing the center of the wound, were selected for analysis. These measurements were made by computer-assisted image analysis (ImagePro Plus, Media Cybernetics, Bethesda, MD, USA). Measurements of the gap between the epithelial edges of the wound as well as the area of connective tissue fill were performed with this software. The numbers of fibroblasts and polymorphonuclear leukocytes (PMNs) were identified by their characteristic morphology in hematoxylin- and eosin-stained sections and counted at 1000X magnification in the connective tissue of the healing wound as we have previously described (Liu et al., 2006). Fibroblasts were identified by their characteristic fusiform shape when sectioned parallel to the long axis or the presence of a “nuclear tail” in cross-section. PMNs have a multi-lobed appearance, which is distinct and readily detected at 1000X magnification. The results are expressed as the cell number per area of connective tissue.
TUNEL Assay
Apoptotic cells were identified with a TUNEL assay in situ by means of a TACS 2 TdT-Blue Label kit purchased from Trevigen (Gaithersburg, MD), following the manufacturer’s instructions. The number of TUNEL-positive fibroblasts were counted at 1000X magnification. Apoptotic fibroblastic cells had a nucleus with a fusiform appearance when sectioned parallel to the long axis, or a nuclear “tail” in cross-section, which could be distinguished in sections counterstained with hematoxylin following the TUNEL assay.
Immunohistochemistry for Caspase-3 and PCNA
Immunopositive cells were identified by immunohistochemistry with an antibody specific for cleaved caspase-3 (Cell Signaling, Danvers, MA, USA) or PCNA (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Biotin-labeled secondary antibody and avidin-biotin-peroxidase complex (Vector Laboratories, Burlingame, CA, USA) were used to localize the primary antibody. Fibroblastic cells that were immunopositive for caspase-3 and PCNA were counted at 1000X magnification. Immunopositive fibroblasts were distinguished from other cells by their characteristic appearance when counterstained with hematoxylin following immunostaining. There was no immunostaining with matched control IgG for each antibody (data not shown).
FOXO1 Nuclear Translocation
FOXO1 nuclear translocation was detected by fluorescence microscopy. Primary antibody to FOXO1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was detected with a biotinylated secondary antibody followed by streptavidin-Texas Red (Vector Laboratories, Burlingame, CA, USA) with DAPI nuclear stain in the mounting medium (Vector Laboratories). We scanned the new healing tissue for the presence of FOXO1 in the nuclear compartment by comparing FOXO1 staining with Texas Red, nuclear stain with DAPI, and a corresponding merged image. Matched control antibody was used as a negative control.
Results
Impaired Oral Wound Healing in Type 2 Diabetes
To assess wound healing, we measured the degree of epithelial coverage and formation of connective tissue. Healing of excisional wounds created in the anterior palatal gingiva of both type 1 and type 2 diabetic mice was impaired compared with that in matched normoglycemic control mice. The epithelial gap was almost closed in the normoglycemic mice and was 6-fold greater in the type 1 diabetic mice (Fig. 1A) (p < 0.05). The amount of connective tissue fill of the wounded tissue (Fig. 1B) was 2-fold higher in the normoglycemic compared with type 1 diabetic mice (p < 0.05). In addition, there were 3-fold more PMNs (p < 0.05) in healing connective tissue in type 1 diabetic mice compared with the control mice (Fig. 1C).
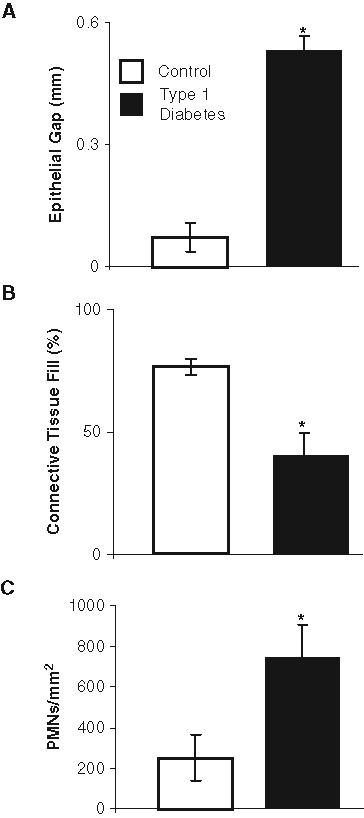
Impaired gingival wound healing in type 1 diabetes. A 1.5-mm wound was created in the anterior palatal gingiva. Healing was examined 5 days after wounding in streptozotocin-induced type 1 diabetic mice and compared with that in normoglycemic controls by assessment of the gap between the healing edges of the epithelium
The effect of diabetes on wound healing in type 2 mice was similar to that in type 1 diabetic mice. The epithelial gap in type 2 diabetic mice was 3-fold greater than that in the matched control mice (Fig. 2A). The amount of connective tissue fill of the wounded tissue (Fig. 2B) was 2-fold higher in the normoglycemic compared with type 2 diabetic mice (p < 0.05). In type 2 diabetic mice, there were 3-fold more PMNs (p < 0.05) in the area of the wounded tissue compared with those in control mice (Fig. 2C).
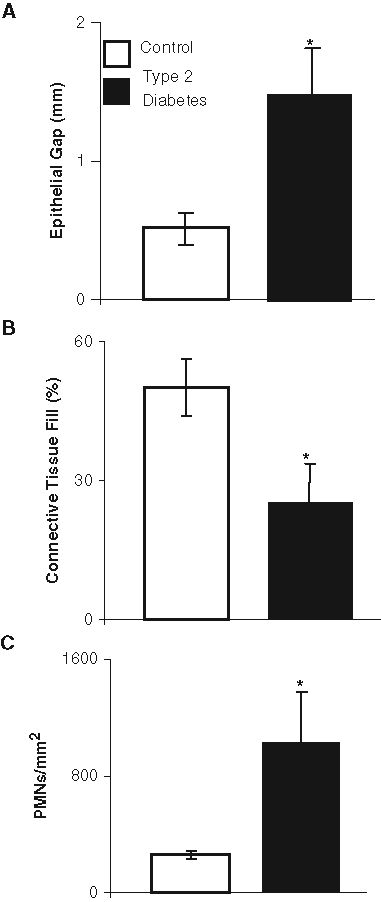
Impaired gingival wound healing in type 2 diabetic mice. Healing was examined 5 days after wounding in db/db type 2 diabetic and normoglycemic littermates, by assessment of the gap between the healing edges of the epithelium
Impact of Diabetes on Fibroblasts in Wound Healing
We then undertook experiments in type 2 diabetic mice to investigate the impact of diabetes on fibroblasts in oral wounds. Fibroblast density in the healing connective tissue was 2.5-fold higher in normoglycemic compared with diabetic mice (Fig. 3A). To determine how this difference may occur, we assessed fibroblast proliferation by measuring the number of PCNA-positive fibroblasts and the apoptotic fibroblasts by the TUNEL assay. The diabetic mice had half the number of PCNA-positive fibroblasts as normoglycemic mice (Fig. 3B) (p < 0.05). The number of apoptotic fibroblasts was ~ 6-fold higher in the diabetic group compared with the normoglycemic group (Fig. 3C). A photomicrograph of apoptotic fibroblasts is shown in the Appendix Figure. We further examined the fibroblastic cells that expressed activated caspase-3, which is a critical effector caspase that induces apoptosis (Graves et al., 2006) (p < 0.05). Type 2 diabetic mice revealed 3-fold more fibroblasts with activated caspase-3 compared with control mice (Fig. 3D) (p < 0.05).
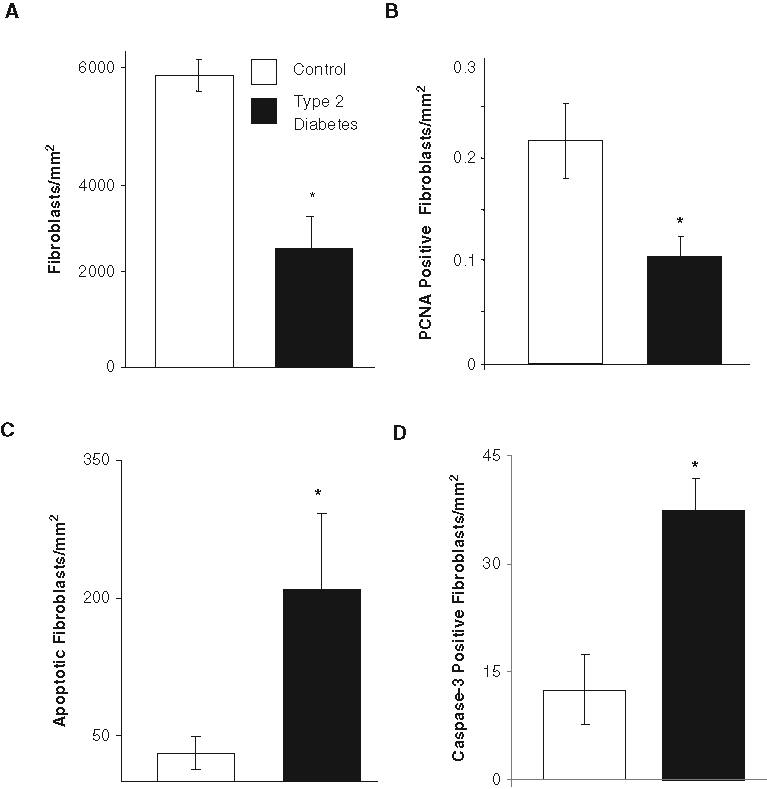
Increased fibroblast apoptosis in gingival wound healing in type 2 diabetic mice. The newly formed connective tissue 5 days after wounding was examined for fibroblast density
FOXO1 Nuclear Translocation
FOXO1 regulates fibroblast apoptosis (Alikhani et al., 2005) and is decreased under conditions when gingival fibroblast apoptosis is reduced (Kantarci et al., 2007). We measured translocation of FOXO1 to the nuclear compartment in cells with a fibroblastic appearance by examining merged images representing FOXO1-immunopositive cells and DAPI-stained nuclei (Fig. 4A). The number of fibroblastic cells with FOXO1 nuclear translocation was 3-fold higher in wounds of diabetic compared with matched normoglycemic mice (p < 0.05) (Fig. 4B).
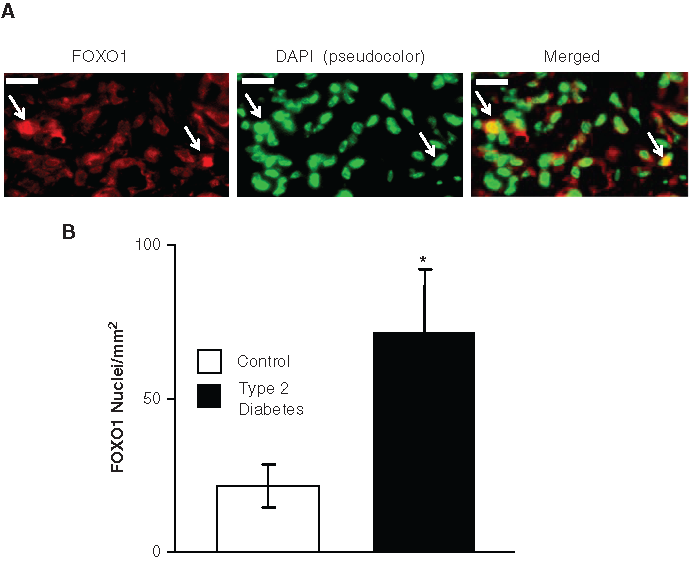
Increased FOXO1 nuclear translocation in fibroblasts in wounds of type 2 diabetic mice.
Discussion
Wound healing is a complex process that is regulated by the coordinated action of several cell types. Based on our previous studies, we examined a time-point where healing had progressed, but where we could also distinguish compromised from normal healing (Graves et al., 2001; Nooh and Graves, 2003). The results indicate that diabetic mice exhibited significantly impaired formation of gingival connective tissue compared with normoglycemic mice. Diabetic healing was characterized by the presence of more PMNs, consistent with a more persistent inflammation in diabetic mice and reduced connective tissue formation. This was evident in both type 1 and type 2 diabetic mice, establishing that the results are not restricted to a specific model of diabetes.
To explain the decreased healing, we examined two parameters, apoptosis and proliferation in the type-2 diabetic mice. There were reduced numbers of proliferating fibroblasts, as determined by fewer PCNA-immunopositive fibroblastic cells. Diabetes caused an increase in the number of apoptotic fibroblasts. The latter is in agreement with other studies showing increased fibroblast apoptosis in murine diabetic animal models (Darby et al., 1997; Al-Mashat et al., 2006) and human diabetic wounds (Rai et al., 2005; Niu et al., 2008). Several lines of evidence support both as being important in the reduced number of fibroblasts in diabetic healing. Significantly reduced numbers of PCNA-positive fibroblasts were observed in the diabetic animals, consistent with reduced proliferation of these cells. The number of TUNEL-positive fibroblasts was significantly higher in type 2 diabetic compared with normoglycemic wounds. This may be explained by the significantly elevated number of fibroblasts with activated caspase-3 in the diabetic compared to normoglycemic wounds, agreeing well with the TUNEL assay results.
There are several mechanisms by which diabetes could cause elevated apoptosis. A common feature of diabetic complications in humans and murine models is increased levels of inflammatory cytokines, advanced glycation end-products, and reactive oxygen species (Graves et al., 2006; Kanwar et al., 2008). They stimulate fibroblast apoptosis through conserved pathways in mice and human fibroblasts that involve caspase-3 (Alikhani et al., 2007; Chen et al., 2007). High glucose treatment has been shown to induce ATP-mediated morphological changes, caspase-3 activation, and apoptosis in fibroblasts (Solini et al., 2000). Moreover, fibroblast apoptosis is elevated in healing diabetic human (Rai et al., 2005; Ladin et al., 1998; Messadi et al., 1999; Rai et al., 2005; Danciu et al., 2004) and diabetic murine dermal wounds (Danciu et al., 2004; Al-Mashat et al., 2006).
Pro-apoptotic factors such as advanced glycation end-products and TNF-α stimulate fibroblast apoptosis by up-regulating pro-apoptotic gene expression through a process regulated by the transcription factor FOXO1 (Alikhani et al., 2005, 2007). In the inactive state, FOXO1 is restricted to the cytoplasm and, upon activation, translocates to the nucleus and regulates gene transcription. To assess the level of FOXO1 activation, we measured the number of fibroblastic cells where FOXO1 was clearly detected in the nucleus. In diabetic wounds, the number was 3-fold greater than in normoglycemic controls. The increase in FOXO1 nuclear translocation is consistent with the higher level of apoptosis and reduced proliferation in the diabetic wounds.
In conclusion, impaired oral wound healing seen in diabetic mice was linked to several aspects of increased fibroblast apoptosis, including enhanced cleaved caspase-3 and FOXO1 nuclear translocation. The results may suggest a model whereby diabetes increases the pro-apoptotic microenvironment of the healing wound, leading to a decrease in fibroblast numbers and diminished healing capacity.
Footnotes
Acknowledgements
We thank Dr. Sadru Kabani, DMD, Oral Pathology, Boston, MA, for help in cell counts.
This work was supported by grants R01 DE017732 and R01 DE019108 from the National Institute of Dental and Craniofacial Research (NIDCR).
The authors have no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.