
Abstract
Prostaglandin E2 (PGE2) plays important roles in the bone resorption of inflammatory diseases such as rheumatoid arthritis and periodontitis via specific prostaglandin receptors (i.e., EP1-EP4). In this study, the authors examined whether Prevotella intermedia regulates PGE2 production and EP expression in human periodontal ligament fibroblasts (hPDLs); they also explored the potential signaling pathways involved in PGE2 production. P. intermedia induced PGE2 production and cyclooxygenase-2 (COX-2) expression in a dose- and time-dependent manner. Indomethacin and NS-398 completely abrogated the P. intermedia–induced PGE2 production without modulating COX-2 expression. Specific inhibitors of extracellular signal-regulated kinase, c-Jun N-terminal kinase, p38, phosphatidylinositol 3-kinase, and protein kinase C—but not c-AMP and protein kinase A—significantly attenuated the P. intermedia–induced COX-2 and PGE2 expression. P. intermedia reduced EP1 expression in a concentration- and time-dependent manner. The results indicate that the COX-2-dependent induction of PGE2 by P. intermedia in hPDLs is mediated by multiple signaling pathways.
Prostaglandins—particularly, prostaglandin E2 (PGE2)—play important roles in bone metabolism. In physiological conditions, such as response to mechanical forces and bone fracture healing, PGE2 stimulates bone formation (Raisz, 2001; Radi and Khan, 2005), whereas under pathological conditions, such as osteoporosis, rheumatoid arthritis, periodontal diseases, and cancerous bone diseases, PGE2 plays a critical role in bone resorption (Miyaura et al., 1995; Ono et al., 2002; Noguchi and Ishikawa, 2007).
The biosynthesis of prostaglandins involves 3 consecutive steps: (1) the release of arachidonic acid from cell membrane phospholipids by phospholipase A2; (2) the conversion of arachidonic acid into prostaglandin H2 by cyclooxygenases (COX-1 and COX-2); and (3) the conversion of prostaglandin H2 into different prostanoids, including PGE2, by specific prostaglandin synthases. COX-1 is constitutively expressed in most cells and regulates normal physiological responses, whereas COX-2 is induced by various stimuli, including cytokines, lipopolysaccharide (LPS), and growth factors (Tanabe and Tohnai, 2002). Therefore, COX-2 seems to be the primary COX that controls PGE2 synthesis in response to inflammation.
The biological activities of PGE2 are mediated through multiple functionally distinct G protein–coupled receptors—namely, EP1-EP4 (Narumiya et al., 1999; Suzawa et al., 2000). EP1 receptor is coupled to Ca2+ signaling and can lead to elevated intracellular calcium levels. EP2 and EP4 receptors activate adenylate cyclase and stimulate intracellular cAMP production and PKA signaling. EP2 and EP4 play important roles in regulating bone formation and resorption. Multiple isoforms of EP3 receptors mediate several signaling pathways, including the inhibition and stimulation of adenylate cyclase and the mobilization of intracellular calcium.
Chronic periodontitis is an inflammatory disease initiated by periodontal pathogens and characterized by periodontal connective tissue destruction and alveolar bone resorption. Prevotella intermedia is one of the major periodontal pathogens that can modulate host immune response and cause tissue destruction (Eley and Cox, 2003). We recently demonstrated that P. intermedia increases matrix metalloproteinases expression and induces proinflammatory cytokines expression in human periodontal ligament cells (Guan et al., 2008, 2009a,b). However, the effect of P. intermedia on PGE2 production has not been explored.
To extend our understanding of the virulent roles of P. intermedia, we examined whether P. intermedia stimulates PGE2 production; we also investigated the signaling pathways involved in P. intermedia–induced PGE2 production.
Materials & Methods
Bacterial Culture
P. intermedia ATCC25611 and ATCC49046 were plated on trypticase agar supplemented with 5% defibrinated rabbit blood in a Bugbox anaerobic workstation at 37°C (Ruskinn Life Sciences Ltd, Wales, United Kingdom) under 80% N2, 10% CO2, and 10% H2. A single colony was inoculated into 10 mL trypticase soy broth (BBL Microbiology Systems, Cockeysville, Maryland) supplemented with yeast extract (5 mg/mL), menadione (5 µg/ mL), and hemin (5 µg/mL). After 24 hours, the bacteria were further cultured by 5% inoculation. They were centrifugated (10,000 g, 15 minutes, 4°C) after 20 hours. The culture supernatant was filter sterilized with a 0.22-µM filter and stored at −80°C until use.
Cell Culture
Healthy human periodontal ligament tissue was obtained from the middle third of extracted premolars of orthodontic patients with informed consent. Experimental protocols were approved by the Ethics Committee of School of Stomatology, the Fourth Military Medical University. The cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Carlsbad, California) as supplemented with 10% heat-inactivated fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin. Cell cultures were maintained at 37°C in a humidified atmosphere (5% CO2, 95% air). Cells between the fifth and eighth passages were used in this study.
Treatment of Human Periodontal Ligament Fibroblasts (hPDLs) by P. Intermedia
The hPDLs were plated in 6-well plates (1 × 106 cells/well) and cultured until confluence. The confluent cells were silenced overnight with serum-free DMEM and subsequently treated with 1%-15% P. intermedia supernatant, prepared by diluting aliquots of P. intermedia supernatant in DMEM, for 24 hours. The time-course effects were determined by treating hPDLs with 5% P. intermedia supernatant for 3 to 72 hours. Cells treated with same amount of blank bacterial culture medium served as controls. All experiments were conducted in duplicate and in 3 different cell lines.
To examine the signaling pathways involved in P. intermedia–induced PGE2 production, the hPDLs were preincubated with PD98059, SP600125, SB203580, LY294002, bisindolylmaleimide, Rp-adenosine 3′,5′-cyclic monophosphorothioate triethylammonium salt hydrate (Rp-cAMP), and H89 (Sigma-Aldrich, St Louis, Missouri), which are specific inhibitors of extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38, phosphatidylinositol 3-kinase (PI3K), protein kinase C (PKC), cAMP, and protein kinase A (PKA), for 1 hour before a 24-hour 5% P. intermedia stimulation.
PGE2 Measurement by ELISA
Secreted PGE2 in conditioned culture media was measured by an ELISA kit according to the manufacturer’s instructions (R&D Systems, Minneapolis, Minnesota). All determinations were done in triplicate.
Blockage of PGE2 Synthesis by COX Inhibitors
To examine whether the P. intermedia–induced PGE2 production was mediated by COX-2 expression, indomethacin and NS-398 (Sigma-Aldrich) were incorporated into the culture medium for 1 hour before P. intermedia stimulation.
Reverse Transcription Polymerase Chain Reaction
Total RNA was extracted from hPDLs with the Total RNA Extraction Kit I (Omega Bio-Tek, Norcross, Georgia) and quantified spectrophotometrically. Total RNA (500 ng) was reverse transcribed with the PrimeScript RT Reagent Kit (TaKaRa Biotechnology, Dalian, China) using OligodT(18) primer in a 20-µL reaction system. The reverse transcription was conducted following the manufacturer’s instructions. Polymerase chain reaction was carried out with 2 µL reverse transcript with 1 U Platinum Taq DNA polymerase (Invitrogen). The COX primer sequences have been described (Kanzaki et al., 2002), whereas the EP primer sequences were found at http://pga.mgh.harvard.edu/primerbank/ (Table). Amplification was performed with denaturation at 94°C for 35 seconds, annealing at respective temperatures for 35 seconds, extension at 72°C for 45 seconds, and a final extension at 72°C for 10 minutes. In addition, 5-µL polymerase chain reaction products were run on 1.5% agarose gels and stained by ethidium bromide. The stained gels were visualized with the Gel Doc XR system (Bio-Rad, Hercules, California), and band intensities were analyzed with AlphaView 1.3.0 (Alpha Innotech Corp, San Leandro, California).
Primer Sequences and Polymerase Chain Reaction Conditions a
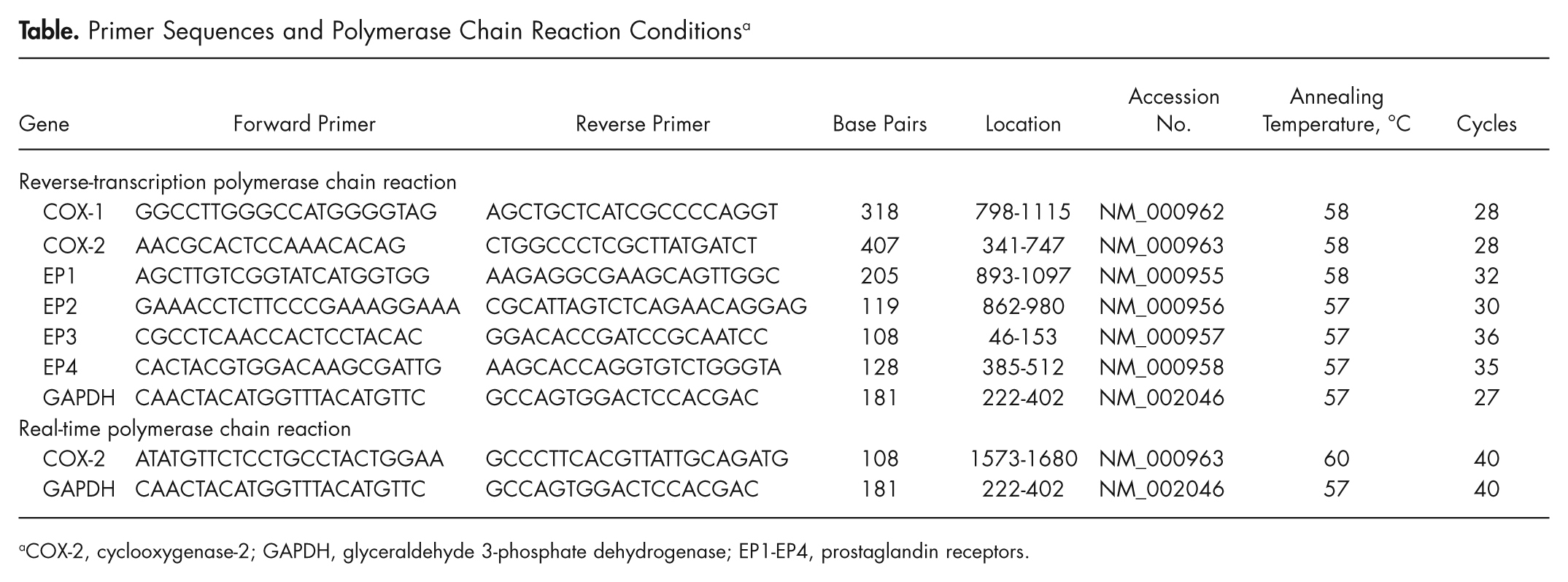
COX-2, cyclooxygenase-2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; EP1-EP4, prostaglandin receptors.
Western Blot
At the indicated times, hPDLs were solubilized with RIPA lysis buffer (Promega, Madison, Wisconsin). The lysates were centrifugated (15,000 g, 30 minutes, 4°C), and protein concentrations in the supernatants were determined by the BCA Protein Assay Kit (Thermo Scientific, Rockford, Illinois). Proteins (15 µg) were boiled with SDS-PAGE sample buffer, separated with 10% SDS-PAGE separation gels, and transferred to PVDF membrane. The membranes were blocked with 5% nonfat milk in phosphate buffered saline containing 0.05% Tween-20 for 1 hour at room temperature and subsequently incubated with 1:200 mouse anti-human COX-2 monoclonal antibody (sc-19999, Santa Cruz Biotechnology, Inc, Santa Cruz, California) or 1:5000 mouse anti-human monoclonal α-tubulin antibody (T6074, Sigma-Aldrich) overnight at 4°C. The membranes were then incubated with 1:5000 horseradish peroxidase–conjugated goat anti-mouse immunoglobulin G (Zhongshan Goldenbridge Biotechnology, Beijing, China) for 1 hour at room temperature and developed with the ChemiGlow West chemiluminescent substrate kit (Alpha Innotech). The signals were captured with FluorChem FC2 (Alpha Innotech).
Data Analysis
Data were expressed as means ± standard error. One-way analysis of variance, followed by post hoc Bonferroni’s multiple comparison test, was used to compare the results with GraphPad Prism 5 (GraphPad Software Inc, San Diego, California). P values of < .05 were considered statistically significant.
Results
To find out whether P. intermedia induces PGE2 production in hPDLs, we first measured secreted PGE2 concentrations in the conditioned culture media by ELISA. Both P. intermedia ATCC25611 (Figure 1A and 1B) and ATCC49046 (Figure 1C and 1D) increased PGE2 production in a dose- and time-dependent manner; furthermore, 15% P. intermedia showed maximal PGE2 stimulatory effects (P < .001) (Figure 1A and 1C). Time course study showed that at 6 hours poststimulation, secreted PGE2 was significantly elevated (P < .05) and the PGE2 level reached a peak at 72 hours poststimulation (P < .001) (Figure 1B and 1D).
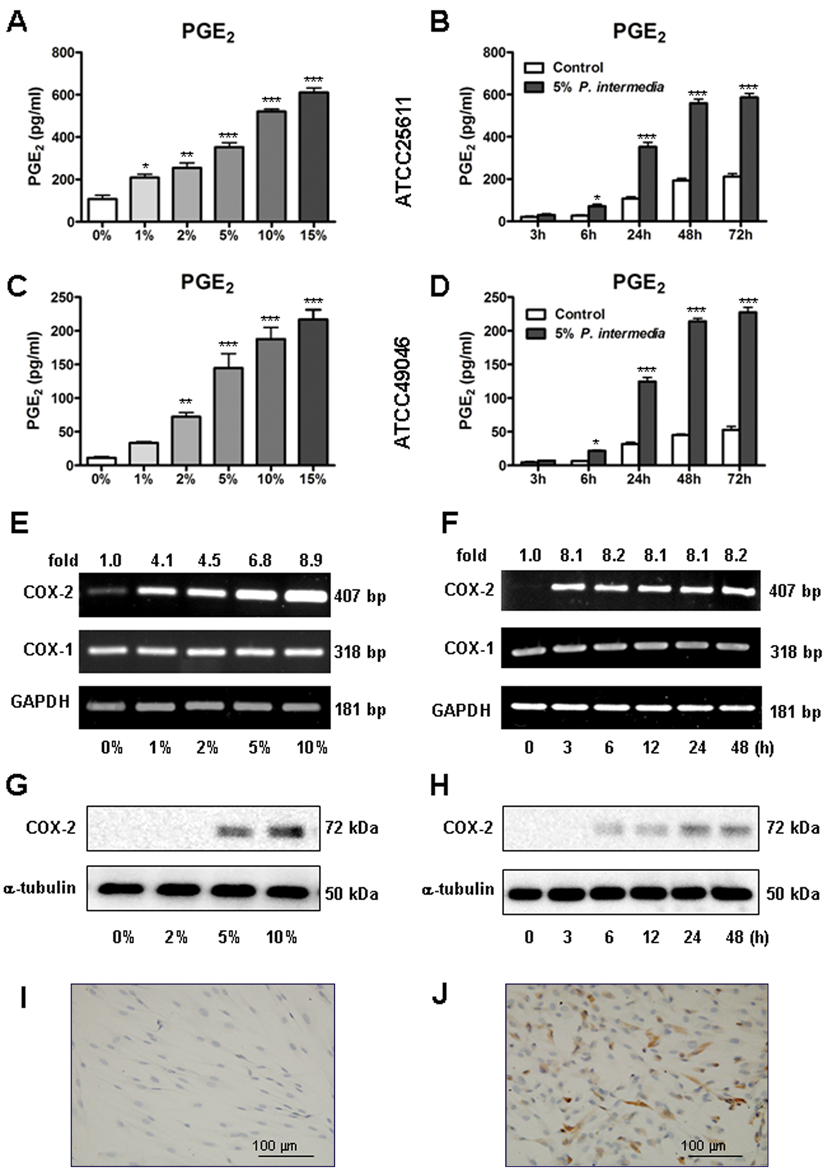
Effects of Prevotella intermedia on prostaglandin E2 (PGE2) production and cyclooxygenase-2 (COX-2) expression in human periodontal ligament fibroblasts (hPDLs). hPDLs were stimulated with various concentrations of P. intermedia supernatant for the indicated times. PGE2 levels in the culture media were evaluated by ELISA. COX-2 mRNA expression was determined by reverse transcription polymerase chain reaction, and COX-2 protein expression was analyzed by Western blot and immunocytochemistry.
COX-1 mRNA was constitutively expressed, while COX-2 was weakly expressed, in hPDLs without P. intermedia stimulation. P. intermedia treatment induced a dose-dependent increase of COX-2 mRNA and protein expression (Figure 1E and 1G). Moreover, P. intermedia induced COX-2 mRNA expression maximally within 3 hours poststimulation (8.1-fold increase, P < .001), and the effects sustained for 48 hours (Figure 1F). The induced COX-2 protein expression was not significant until 6 hours poststimulation, and it increased time dependently thereafter (Figure 1H). P. intermedia stimulation produced no detectable change in COX-1 mRNA expression (Figure 1E and 1F). Immunocytochemistry confirmed that after 24 hours of P. intermedia stimulation, COX-2 expression was significantly enhanced (Figure 1J).
To examine whether the P. intermedia–induced PGE2 production was mediated by COX-2, we used nonselective and specific COX-2 inhibitors—indomethacin and NS-398—to block the actions of COX-2. Indomethacin and NS-398 completely abrogated the P. intermedia–induced PGE2 production (P < .001) (Figure 2B) but did not influence the induced COX-2 mRNA and protein expression (Figure 2A).
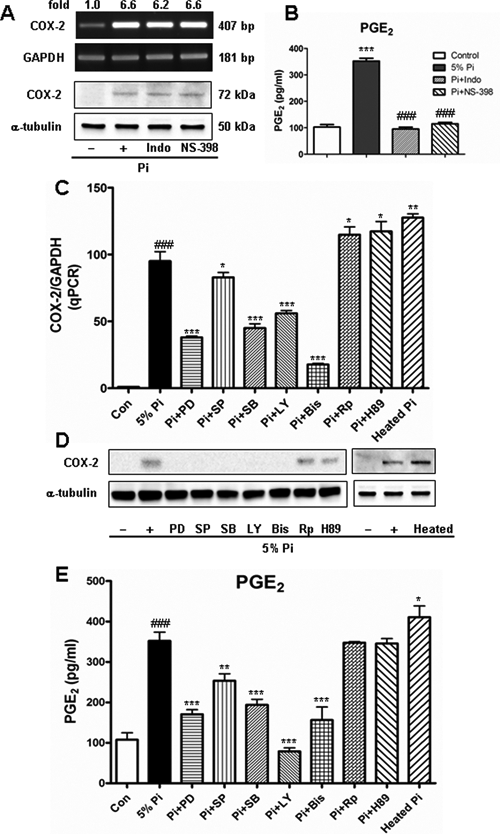
Effects of cyclooxygenase (COX) inhibition and signaling pathway inhibition on Prevotella intermedia ATCC25611-induced COX-2 expression and prostaglandin E2 (PGE2) production in human periodontal ligament fibroblasts, which were pretreated with 20 µM indomethacin (Indo), 1 µM NS-398, 20 µM PD98059, 10 µM SP600125, 20 µM SB203580, 5 µM LY294002, 0.5 µM bisindolylmaleimide (Bis), 10 µM Rp-cAMP, and 10 µM H89 for 1 hour before a 24-hour P. intermedia ATCC25611 stimulation.
To investigate the signaling pathways involved in P. intermedia–induced PGE2 production and COX-2 expression in hPDLs, we preincubated the hPDLs with specific inhibitors of ERK, JNK, p38, PI3K, PKC, cAMP, and PKA before P. intermedia challenge. ERK, JNK, p38, PI3K, and PKC inhibitors significantly reduced the P. intermedia–induced COX-2 mRNA (P < .05 or P < .001; Figure 2C) and protein expression (Figure 2D). They also significantly decreased PGE2 production (P < .01 or P < .001; Figure 2E). Rp-cAMP and H89—inhibitors of cAMP and PKA—slightly increased the induced COX-2 expression and PGE2 production (Figure 2C to 2E).
To find out the responsible components in P. intermedia supernatant for COX-2 and PGE2 induction, we heated the supernatant at 95°C for 30 minutes before incubating it with hPDLs. Heat treatment did not reduce the P. intermedia–induced COX-2 expression and PGE2 production; on the contrary, it significantly increased COX-2 expression and PGE2 production (Figure 2C to 2E), indicating that heat-resistant LPS or glycoprotein components, rather than heat-labile proteinaceous components, were responsible for the effects of P. intermedia on COX-2 and PGE2 induction.
EP1, EP2, EP3, and EP4 mRNA expression were detected in hPDLs. The sequential expression intensities of the EPs were EP2 > EP1 > EP4 > EP3 (Figure 3A, Table). P. intermedia significantly reduced EP1 expression in a dose-dependent manner (Figure 3A and 3B) and time-dependent manner (Figure 3C and 3D), whereas it showed no regulating effects on other EPs (Figure 3A and 3B).
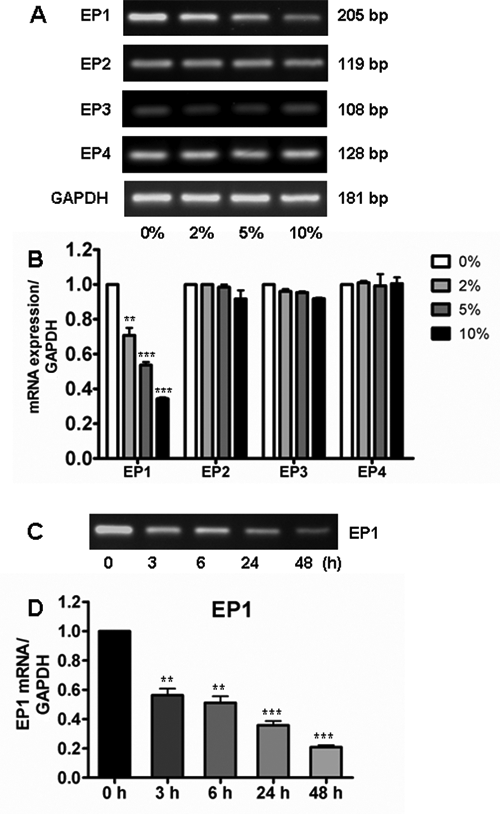
Effects of Prevotella intermedia ATCC25611 stimulation on prostaglandin receptors (EP1-EP4) expression. human periodontal ligament fibroblasts were stimulated with various concentrations of P. intermedia supernatant for the indicated times. Total RNA was extracted and subjected to reverse transcription polymerase chain reaction.
Discussion
In the present study, we demonstrated for the first time that P. intermedia induced PGE2 production in a concentration- and time-dependent manner in hPDLs. In addition, P. intermedia augmented COX-2, but not COX-1, gene expression. Indomethacin and NS- 398—nonselective and selective COX-2 inhibitors—completely attenuated the induced-PGE2 production, suggesting that P. intermedia–induced PGE2 production is mediated by COX-2. Our results are in agreement with those of previous studies that demonstrated that NS-398 significantly reduced PGE2 production in colon cancer cells and vascular smooth muscle cells (Banu et al., 2007; Choi et al., 2008) but did not affect COX-2 expression. Our results are also in accord with the notion that nonsteroidal anti-inflammatory drugs, including indomethacin and NS- 398, exert their effects on COX-2 activity by binding to the active site of cyclooxygenase (DeWitt, 1999).
Our study indicated that P. intermedia LPS might be responsible for the effects of P. intermedia on COX-2 and PGE2 induction. LPS of other periodontal pathogens, such as Porphyromonas gingivalis, Actinobacillus actinomycetemcomitans, Prevotella nigrescens, Treponema denticola, and Tannerella forsythia, were reported to stimulate PGE2 production and COX-2 expression in various cell types, including gingival fibroblasts, monocytes, bone marrow cells, and macrophage/epithelial cells (Noguchi et al., 1996; Ueda et al., 1998; Noguchi et al., 2000; Bodet et al., 2006; Chung et al., 2006), indicating that the COX-2 and PGE2 stimulatory effect is a common phenomenon to periodontal pathogens.
PGE2 has been suggested to play a role in the pathogenesis of periodontal disease (Noguchi and Ishikawa, 2007). In vitro studies have demonstrated that PGE2 stimulates RANKL (receptor activator of NF-κB ligand) expression (Kanzaki et al., 2002; Nukaga et al., 2004; Oka et al., 2007) and downregulates OPG (osteoprotegerin) expression (Sakata et al., 2002; Oka et al., 2007) in various periodontal cell types, hence disrupting the balance of RANKL/OPG and promoting osteoclastogenesis. Furthermore, PGE2 up regulates interleukin 1α–induced interleukin 6 expression (Noguchi et al., 2002, 2007), indicating its role in inflammatory responses. We previously demonstrated that PGE2 inhibition resulted in a significant decrease in P. intermedia–induced matrix metalloproteinases and proinflammatory cytokines expression (Guan et al., 2009a,b), suggesting that PGE2 is a main mediator in P. intermedia–induced inflammatory responses in hPDLs.
The signaling pathways involved in PGE2 production in hPDL cells are poorly understood, although some studies demonstrated the involvement of MAPK, PI3K-Akt, PKC, and NF-κB signaling pathways in cytokine- or LPS-induced COX-2 expression and PGE2 production in other cell types, including human chondrocytes (Nieminen et al., 2005), odontogenic keratocyst fibroblasts (Ogata et al., 2007), and mouse intestinal myofibroblasts (Walton et al., 2009). Recently, ERK involvement in interleukin 1α–induced COX-2/PGE2 expression was reported in hPDLs (Fukushima et al., 2005). Our results demonstrate that P. intermedia–induced COX-2/PGE2 is regulated by various signaling pathways. COX-2 expression and PGE2 production were not completely abolished, but rather significantly reduced, by blockage of individual ERK, JNK, p38, PI3K, and PKC signaling pathways, indicating that these pathways are independently involved in COX-2/PGE2 expression in stimulated hPDLs. Whether there exist cross-talks among MAPK, PI3K, and PKC signaling pathways in P. intermedia–induced COX-2/PGE2 production remains to be clarified. Furthermore, given that we used P. intermedia culture supernatant, which contains various components, to stimulate hPDLs, we cannot rule out the possibility that there may be multiple components responsible for activating the different signaling pathways.
PGE2 exerts its action through binding to different receptors. In the present study, all 4 EPs were detected in hPDLs. Noguchi et al. (2002) previously reported EP1, EP2, and EP4, but not EP3, mRNA detection in human gingival fibroblasts. We first reported EP3 expression in hPDLs. The discrepancy concerning EP3 expression in our study and the previous study might be due to either of the following reasons: (1) Different cell types were investigated; that is, we used hPDLs, whereas human gingival fibroblasts were used in the previous study. (2) Different amplification cycles were adopted; that is, we amplified EP3 for 36 cycles, yet the EP3 signal was still very low, whereas the previous study amplified all EPs for 35 cycles. EPs regulation has rarely been studied despite a recent study showing that Escherichia coli LPS significantly decreased EP1, EP2, and EP3 expression, but increased EP4 expression, in osteoblasts (Shoji et al., 2006). We first reported that P. intermedia reduced EP1 expression in a dose- and time-dependent manner.
The role of EP1 in bone metabolism has not been clearly defined. PGE2 enhanced interleukin 1α– or interleukin 1β–induced interleukin 6 expression in mouse cementoblasts (Noguchi et al., 2007) and in human gingival fibroblasts via EP1 (Noguchi et al., 2002). Moreover, EP1 antagonist SC-19220 inhibited RANKL- and interleukin 6–induced osteoclastogenesis (Inoue et al., 1999; Tsujisawa et al., 2005). These studies indicate that EP1 plays certain roles in inflammation and bone absorption. At present, we are unable to explain what effects the P. intermedia–reduced EP1 may have in the development of periodontitis, and this needs to be addressed in future studies.
In summary, we demonstrated that P. intermedia induced PGE2 production via COX-2 upregulation through multiple signaling pathways. Taken with our previous results, it can be reasonably concluded that P. intermedia regulates diverse inflammatory and immune responses and may hence play a role in tissue destruction during chronic periodontitis.
Footnotes
Acknowledgements
This study was supported by Shaanxi Natural Science Foundation (2010JM4017). The authors are grateful to Prof. Yan Jin for his support of the study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.