
Abstract
Agenesis of the permanent teeth is a congenital anomaly that is frequently seen in humans. Oligodontia is a severe type of tooth agenesis involving 6 or more congenitally missing teeth, excluding the third molars. Previous studies have indicated that mutations in the homeobox gene MSX1, paired domain transcription factor PAX9, and EDA are associated with non-syndromic oligodontia. This study reports a Japanese family (eight of 14 family members affected) with non-syndromic oligodontia who preferentially lacked molar teeth. In this family, a novel frameshift mutation (321_322insG) was identified in the paired domain of PAX9. The frameshift mutation caused altered amino acids in the paired domain and premature termination of translation by 26 amino acids. When transfected into COS-7 cells, the mRNA expression of 321_322insG PAX9 was comparable with that of wild-type PAX9. However, the mRNA of 321_322insG PAX9 was more unstable than that of wild-type PAX9. This mRNA instability caused a marked decrease in protein production, as evaluated by Western blot analysis and immunostaining. These findings suggest that the 321_322insG mutation causes insufficient function of PAX9 protein and haploinsufficiency as a genetic model of familial non-syndromic oligodontia with a PAX9 mutation.
Introduction
Agenesis of the permanent teeth is an anomaly that is frequently seen in humans. The prevalence of agenesis of 1 or 2 teeth (excluding the third molars) is from 1.6% to 9.6% (Symons et al., 1993; Vastardis, 2000). Oligodontia is a severe type of tooth agenesis involving 6 or more congenitally missing teeth, excluding the third molars (Schalk-van der Weide et al., 1992; Stockton et al., 2000). In a study on Danish schoolchildren, 0.16% of the children were found to have oligodontia (Rølling and Poulsen, 2001). To date, 3 genes, namely, the homeobox gene MSX1, paired domain transcription factor PAX9, and EDA, have been associated with non-syndromic oligodontia (Vastardis et al., 1996; Stockton et al., 2000; Tao et al., 2006).
Familial non-syndromic oligodontia associated with PAX9 mutation preferentially affects molar formation (Stockton et al., 2000; Frazier-Bowers et al., 2002; Kapadia et al., 2006). To date, 10 gene mutations have been identified in the DNA-binding paired domain of PAX9 (Nieminen, 2009; Wang et al., 2009). One is a frameshift mutation (219_220insG) (Stockton et al., 2000; Mensah et al., 2004), one is a nonsense mutation (A340T) (Nieminen et al., 2001), and 8 are point mutations (Das et al., 2003; Lammi et al., 2003; Mostowska et al., 2003; Jumlongras et al., 2004; Kapadia et al., 2006; Ogawa et al., 2006; Wang et al., 2009).
In this study, a novel frameshift mutation of PAX9 (321_322insG) was identified in a Japanese family with non-syndromic oligodontia. The purpose of this study was to clarify the aberrant function of this gene mutation. We hypothesized that this PAX9 mutation caused abnormal mRNA and/or protein expression.
Materials & Methods
Family with Oligodontia
The proband was a 37-year-old female (III:3) in a Japanese family with oligodontia treated in Niigata University Hospital (Fig. 1A). IV:1 (an 18-year-old female) was also treated for oligodontia in the hospital, and their panoramic radiographs were used for the diagnosis. Oral examinations and interviews were performed on II:3 and III:2. An interview was performed by phone on I:1, II:5, III:4, and III:5, who lived far from the Dental Hospital of Tokyo Medical and Dental University and Niigata University Hospital. This was carried out by identification of the normal numbers and shapes of permanent teeth of each person. The extraction and loss of any teeth after eruption were carefully queried for each family member. Each individual with more than 6 congenitally missing teeth, excluding the third molars, was diagnosed as having oligodontia for the purposes of this study (Schalk-van der Weide et al., 1992; Stockton et al., 2000). The experimental protocol was approved by the Ethical Review Committee of the Tokyo Medical and Dental University and the Niigata University Graduate School of Medical and Dental Sciences.
Mutational Analysis
For mutational analysis, we obtained genomic DNA from individuals whose informed consent was obtained. Genomic DNA was isolated from nail samples of II:3, III:2, III:3, IV:1, and IV:2. The isolated DNA was amplified by polymerase chain-reaction in a GeneAmp PCR System 9700 and specific primers for PAX9 (Das et al., 2003). The amplified products were sequenced with a PRISM® 3100 Genetic Analyzer and a BigDye Terminator v.3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA).
Plasmid Construct
The human PAX9 cDNA (NM_006194.2) was subcloned in-frame with a C-terminal Myc-tag downstream of the cytomegalovirus (CMV) promoter into the pReceiver-M09 empty vector (termed EX-NEG; GeneCopoeia, Rockville, MD, USA). Site-directed mutagenesis was carried out with the QuickChange mutagenesis kit (Stratagene, La Jolla, CA, USA), as previously reported (Wang et al., 2009). The entire coding sequence of the mutant construct was verified by a PRISM® 3100 Genetic Analyzer and a BigDye Terminator v.3.1 Cycle Sequencing Kit.
Transfection of Cultured Cells
COS-7 cells were cultured in Dulbecco’s modified Eagle’s medium (Wako, Tokyo, Japan) supplemented with 10% fetal bovine serum (Biowest, Nualle, France). All cultures were maintained in a humidified, 5% CO2 atmosphere at 37°C. Transient transfection was performed with the Fugene 6 Transfection Reagent (Roche, Basal, Switzerland), according to the manufacturer’s instructions.
Western Blot Analysis
COS-7 cells transfected with wild-type or mutant PAX9 underwent lysis in buffer containing 50 mM Tris-HCl (pH 7.4), 125 mM NaCl, 0.1% Nonident P-40 (Sigma-Aldrich, St. Louis, MO, USA), and 1 mM each of EDTA (Wako) and phenylmethylsulfonyl fluoride (Sigma-Aldrich), followed by sonication, as described previously (Shiga et al., 2008). Western blot analysis was performed with cell lysates at 12 and 48 hrs after transfection. Briefly, a 20-µg quantity of protein was separated by 10% SDS-PAGE under reducing conditions and transferred electrophoretically to a Hybond-P polyvinylidene difluoride (PVDF) membrane (Amersham, Buckinghamshire, UK). The membrane was probed with an anti-Myc monoclonal antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The bound antibody was detected by ECL plus Western blotting detection reagents (Amersham), according to the manufacturer’s instructions.
Immunohistochemistry
Transfected COS-7 cells were fixed with methanol for 10 min and stained with the anti-Myc monoclonal antibody. After being washed with PBS, the cells were incubated with a fluorescein isothiocyanate (FITC)-conjugated secondary antibody (Santa Cruz Biotechnology, Inc.). After the washing, fluorescence was observed by fluorescence microscopy (AF6000, Leica, Wetzlar, Germany).
RNA Preparation
COS-7 cells were transfected with wild-type or mutated PAX9. At 12 hrs after the transfection, total RNA was isolated from the cells with ISOGEN (Nippon Gene, Tokyo, Japan), according to the manufacturer’s instructions. DNase treatment was performed with TURBO DNA-free (Ambion Inc., Austin, TX, USA). cDNA was synthesized from 1 µg of the total RNA with a QuantiTect Reverse Transcription Kit (Qiagen, Valencia, CA, USA).
Real-time PCR and Examination of mRNA Stability
Each cDNA was used as a template for quantitative analysis of PAX9 mRNA with a 7300 Real Time PCR system (Applied Biosystems). The real-time PCR amplifications were performed with TaqMan gene expression assays (Applied Biosystems), according to the manufacturer’s instructions. The primers and TaqMan probe used were as follows: forward primer, 5′-GGCATCCGACCGTGTGA-3′, reverse primer, 5′-TCCTGGCAAGATCGAGCC-3′, TaqMan probe, 5′-ATCCTGGCGCGA TAC-3′. These primers and the probe corresponded to the N-terminus of the paired domain and upstream of the 321st nucleotide. The expression of human PAX9 was normalized by the amplification of eukaryotic 18S rRNA (product number 4331182; Applied Biosystems).
To examine the mRNA stability, we similarly transfected COS-7 cells with wild-type or mutated PAX9. At 12 hrs after transfection, 1 µg/mL of actinomycin D (Wako) was added to the culture. Total RNA was isolated from the cells at 3 and 6 hrs after the actinomycin D addition. After treatment with TURBO DNA-free and cDNA synthesis with the QuantiTect Reverse Transcription Kit, quantitative analysis of PAX9 mRNA was carried out in the 7300 Real-time PCR System. The mRNA stability was represented as the PAX9 mRNA remaining after normalization by eukaryotic 18S rRNA (100% remaining PAX9 mRNA denoted the value without actinomycin D treatment).
Statistical Evaluations
We used Student’s t test to examine the differences in the relative expression levels of PAX9/18S rRNA and the decay of PAX9 mRNA following addition of actinomycin D, between wild-type and 321_322insG PAX9. The statistical analyses were carried out with StatView (SAS Institute, Cary, NC, USA). Values of P < 0.05 were considered to indicate statistical significance.
Results
The Family with Non-syndromic Oligodontia
The pedigree of the present family with oligodontia is shown in Fig. 1A. Eight (I:1, II:3, II:5, III:2, III:3, III:4, III:5, and IV:1) of 14 family members had oligodontia. In the regular health examinations, the family members had no medical history of abnormalities in their sweat glands, skin, hair, or nails, indicating the non-syndromic type of oligodontia. A panoramic radiograph of the proband (III:3; 37-year-old female) is shown in Fig. 1B. She had tooth agenesis of the upper and lower second premolars and all molars (agenesis of 12 permanent teeth in total, excluding the third molars) (Figs. 1B, 1C). Based on the oral examinations, interviews, and panoramic radiographs, the positions of the missing permanent teeth in this family are summarized in Fig. 1C. Four members (II:3, III:2, III:3, and IV:1) showed tooth agenesis of the premolars and molars, and the agenesis was also seen in the lower central incisor of IV:1.
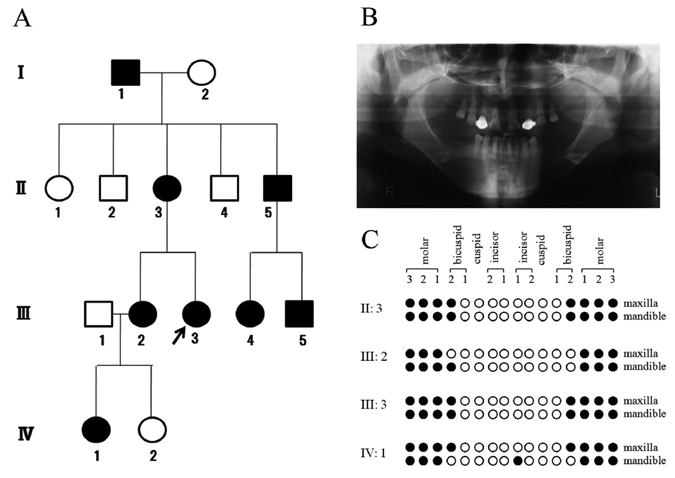
A Japanese family with non-syndrome oligodontia.
DNA Sequence Analysis of the PAX9 Gene
Since this family preferentially lacked teeth in the molar region, the DNA sequence of PAX9 was examined in II:3, III:2, III:3, IV:1, and IV:2. II:3, III:2, III:3, and IV:1 had the same heterozygous insertion of G after the 321st nucleotide according to NM_006194.2 (Fig. 2A). This mutation was not seen in IV:2, who was a healthy control without oligodontia. The 321st nucleotide corresponded to an amino acid in the paired domain (Fig. 2B). This mutation caused a heterozygous frameshift mutation, resulting in premature termination of translation by 26 amino acids (Figs. 2B, 2C).
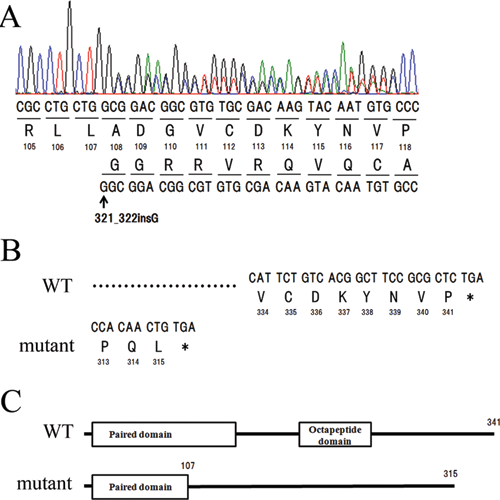
A novel mutation in a Japanese family with non-syndrome oligodontia.
Characteristics of the 321_322insG Mutation in the PAX9 Gene
Next, we transfected the 321_322insG mutated construct into COS-7 cells to examine the characteristics of the mutant protein. Expression was faintly seen at 12 hrs and clearly seen at 48 hrs after the transfection. A band of the expected size (40 kDa) was seen in the cell lysates of wild-type and mutated PAX9 transfected COS-7 cells, but not in the lysate of the empty vector-transfected cells (Fig. 3A). The protein level was much lower in the mutated PAX9-transfected COS-7 cells than in the wild-type PAX9-transfected cells. This finding was also noted in the immunostaining of COS-7 cells with the same antibody (Figs. 3B-3G). Positive staining was seen in the nuclei of COS-7 cells transfected with wild-type PAX9 (Figs. 3C, 3F), but was scarcely seen in the COS-7 cells transfected with the mutant construct (Figs. 3D, 3G). Positive staining was not seen in the empty vector-transfected cells (Figs. 3B, 3E).
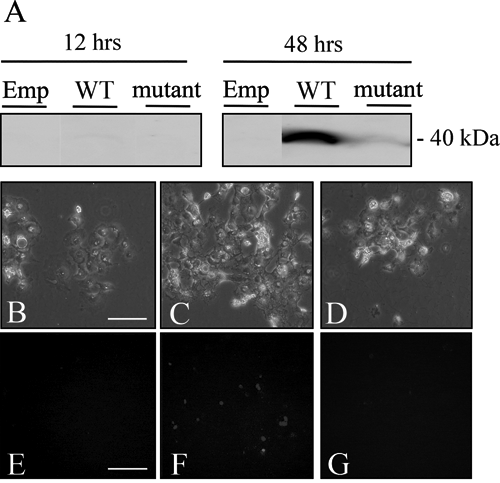
The protein expression of 321_322insG PAX9.
To clarify the mechanism for the decreased level of the mutant protein, we examined the mRNA expressions of the wild-type and 321_322insG PAX9. First, the mRNA expressions were examined by amplification of the N-terminus region of the paired domain. The results showed that the mRNA expression of 321_322insG PAX9 did not differ significantly from that of wild-type PAX9 at 12 hrs after transfection (Fig. 4A). Next, the mRNA stabilities of wild-type and 321_322insG PAX9 were examined. At 12 hrs after transfection, the cells were treated with actinomycin D, an inhibitor of transcription (Fig. 4B). The mRNA levels were decreased for both wild-type and 321_322insG PAX9 at 3 and 6 hrs after the actinomycin D addition. The mRNA of 321_322insG PAX9 was significantly more unstable than the mRNA of wild-type PAX9 during the examined time-course (Fig. 4B).
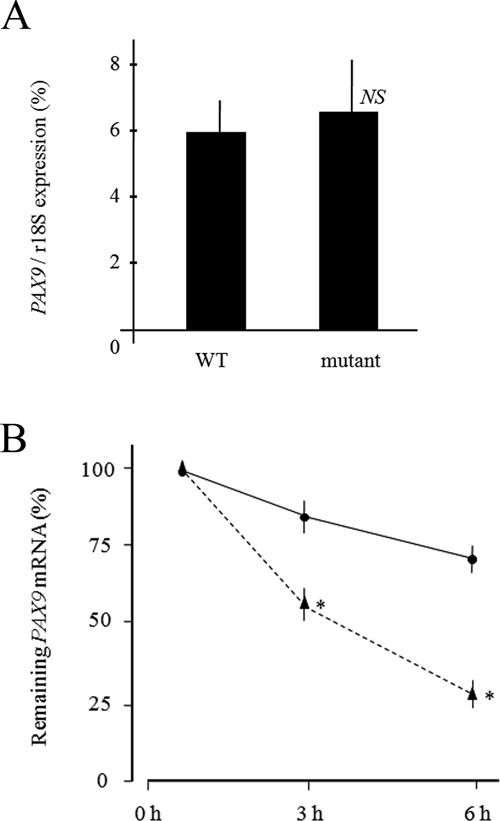
mRNA expression of 321_322insG PAX9.
Discussion
In this study, a novel frameshift mutation of PAX9 (321_322insG) was identified in a Japanese family with non-syndromic oligodontia. The family showed permanent tooth agenesis preferentially in the molar region. In vitro analyses revealed instability of the mRNA of the mutated PAX9, resulting in lower protein production.
The present family had tooth agenesis mainly of the molars. Since I:1, II:5, III:4, and III:5 did not live close to our universities, it was not possible to determine their tooth agenesis directly by oral examination or panoramic radiograph. However, the interview provided us with sufficient information that they had tooth agenesis in the molar region. In addition to the molars, II:3, III:2, III:3, and IV:1 had tooth agenesis of the premolars. This type of tooth agenesis was first reported in a family with a mutation in the paired domain of PAX9 (Stockton et al., 2000). The reported family also showed missing second premolars and mandibular central incisors. Interestingly, tooth agenesis of the mandibular central incisor was also seen in one member of the present family (IV:1). The 321_322insG PAX9 mutation was identified in this family. Ten different gene mutations have been identified in the paired domain of PAX9 to date (Nieminen, 2009; Wang et al., 2009), but no previous studies have reported the present frameshift mutation.
It has been reported that 219_220insG Pax9 (frameshift mutation in the paired domain) abrogates translocation of the protein from the cytoplasm to the nucleus (Wang et al., 2009). However, the total protein expression (including both the nuclear and cytoplasmic fractions) of this mutated mouse Pax9 was comparable with that of mouse wild-type Pax9. In contrast to this mouse study, the protein expression of the present human mutated PAX9 was much lower than that of human wild-type PAX9, and the mRNA of 321_322insG PAX9 was significantly more unstable compared with that of the wild-type gene. The results for these mouse and human mutated proteins were obtained by experiments with the same COS-7 cells and mutagenesis kit. Therefore, it is likely that these two frameshift mutations have different gene functions from one another.
Further studies are required to clarify the instability of 321_322insG PAX9 mRNA. Degradation of mRNA is an important mechanism for controlling gene expression, and eukaryotic mRNAs are primarily degraded by removal of the 3′ poly(A) tail (deadenylation) and cleavage of the 5′ cap structure (decapping) (Mitchell and Tollervey, 2000; Wilusz et al., 2001). In mammalian cells, it has been reported that PAN2 and CCR4 are the major poly(A) nucleases active in cytoplasmic deadenylation, and that DCP2-mediated de-capping is required after deadenylation (Yamashita et al., 2005). To clarify the mechanism of the mRNA instability, it is essential that we examine the activities of these molecules in the degradation process of the 321_322insG PAX9 mRNA.
The discovery of one oligodontia family with a deletion encompassing the PAX9 locus has suggested haploinsufficiency (not a dominant-negative effect) as a cause of non-syndromic oligodontia with mutations of this gene (Das et al., 2002). The present study showed that the 321_322insG mutation impaired the mRNA stability of PAX9, resulting in lower protein expression. These findings support the model of haploinsufficiency of PAX9 as a cause of non-syndromic oligodontia.
Footnotes
Acknowledgements
This study was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (19659547, 21390546), and for the Supporting Project for Strategic Research of Nihon University School of Dentistry at Matsudo by the Ministry of Education, Culture, Sports, Science, and Technology of Japan. The authors thank Professor Hayato Ohshima (Niigata University Graduate School of Medical and Dental Sciences) for recruiting study participants.