
Abstract
The nitric oxide (NO) receptor enzyme soluble guanylate cyclase (sGC) contains one prosthetic heme group as an αβ heterodimer, and two heterodimer isoforms (α1β1, α2β1) were characterized to have enzyme activity. To test the irreversible inflammation-dependent regulation of sGC in odontoblasts, we incubated decalcified frozen sections of healthy and inflamed human third molars with antibodies against β-actin, nitrotyrosine, inducible nitric oxide synthase (iNOS), α1-, β1-, and α2-subunits of sGC and analyzed them at protein levels by quantitative immunohistochemistry. The irreversible inflammation induced an increase in the signal intensities for nitrotyrosine and iNOS and a decrease for the α1-, β1-, and α2-subunits of sGC in odontoblasts. Inflammatory mediators, reactive oxygen, and nitrogen species may impair the expression of the α1-, β1-, and α2-subunits in odontoblasts. The decrease of sGC at the protein level in inflamed odontoblasts is compatible with a critical role for sGC to mediate biological effects of NO in health.
Keywords
Introduction
Nitric oxide (NO) is synthesized either by the constitutively expressed enzymes neuronal (n) and endothelial (e) NO synthases (NOS) or by the inducibly (i) expressed enzyme iNOS. In response to the physiological stimuli that trigger an intracellular Ca2+ signal, nNOS and eNOS are activated (Fleming, 2010). In resting cells, iNOS is not expressed but is regulated at the gene level by different inflammatory mediators such as bacterial lipopolysaccharide (LPS) or cytokines such as interleukin-1 (IL-1), tumor necrosis factor-α (TNF-α), or IFN-γ (Bogdan, 2001; Nathan, 2006). Under inflammatory conditions corresponding to mRNA and protein synthesis, iNOS generates NO at sustained high levels (Bogdan, 2001; Nathan, 2006).
The NO receptor enzyme soluble guanylate cyclase (sGC) is a heme-containing heterodimeric enzyme consisting of α and β subunits, and expression of both subunits is required for catalytic activity of the enzyme (Buechler et al., 1991; Behrends et al., 1995; Russwurm et al., 1998). Four subunits of sGC have been cloned (α1, β1, α1, and β1), and only two isoforms (α1β1, α2β1) were shown to have enzyme activity (Krumenacker et al., 2004). Homodimeric structures of the sGC (α1α1 and β1β1) have been purified, but little or no enzymatic activity of the homodimeric enzymes was detected (Zabel et al., 1999). In physiologically low concentrations, NO generated by nNOS and eNOS interacts directly with the iron atom in the heme group of the sGC to activate the enzyme to produce the intracellular effector molecule cyclic guanosine 3′,5′-monophosphate (cGMP) (Krumenacker et al., 2004). The formation of the intracellular effector molecule cGMP then triggers a variety of cellular responses, including cell proliferation, cell differentiation, neurotransmission, and vasodilatation (Burette et al., 2002; Krumenacker et al., 2004).
In addition to the nNOS localization in rat molar odontoblasts (Korkmaz et al., 2005), the existence of eNOS in human (Felaco et al., 2000) and rat molar (Korkmaz et al., 2005) odontoblasts indicates that nNOS and eNOS are a physiological source for NO in odontoblasts. BK-induced transient activation of eNOS at Ser1177 and BK-mediated phosphorylation of eNOS at Thr495 in odontoblasts have been detected (Korkmaz et al., 2006). In addition to the direct activation of NO on differentiation of osteoblasts (Hikiji et al., 1997), cGMP is involved in the formation of mineralized nodules (Inoue et al., 1995). The existence of α2- and β1-subunits of sGC and NO-dependent formation of cGMP in odontoblasts has been detected (Korkmaz et al., 2005).
In inflammation states, high concentrations of NO generated in inflammatory cells by sustained activation of iNOS are unstable and combined with high oxidative stress. High concentrations of NO are rapidly oxidized under inflammatory conditions to reactive nitrogen species (RNS). Under conditions of high NO, superoxide (O2 -) interacts with NO to produce the highly toxic peroxynitrite anion (OONO-) (Nathan, 2006). RNS and OONO- inhibit the activity of many proteins, including mitochondrial enzymes and transcription factors, and produce long-term cellular effects (Nathan, 2006). In inflammatory conditions, the induction of iNOS may be a source of high concentrations of NO in the dental pulp (Di Nardo Di Maio et al., 2004).
The regulation and interactions between inflammatory cells and odontoblasts influence the repair process of the dentin-pulp complex (Goldberg et al., 2008). However, the potential effects of inflammatory mediators and high concentrations of NO on protein levels of the α1-, β1-, and α2-subunits of sGC in odontoblasts are unknown. Therefore, we investigated the α1-, α2-, and β1-subunits of the sGC for their protein existence and protein levels in healthy and inflamed odontoblasts by quantitative immunohistochemistry using antibodies against the α1- (Koglin and Behrends, 2003), β1- (Behrends et al., 2001), and α2-subunits (Bamberger et al., 2001) of sGC.
Materials & Methods
Tissue Clinical Parameters
Sound healthy (n = 9) and carious inflamed (n = 9) third molars from 18- to 27-year-old patients were extracted for orthodontic and clinical reasons. The molars of the healthy group were unrestored, asymptomatic, and without pain on percussion. There was no radiographic radiolucency for caries and periapical periodontitis. The third molars with caries and an irreversibly inflamed dentin-pulp complex were extracted for orthodontic reasons (Appendix 1).
Tissue Preparation
Immediately after extraction, the molars were immersion-fixed for 24 hrs in a fixative containing 4% paraformaldehyde and 0.2% picric acid in 0.1 M phosphate-buffered saline (PBS), pH 7.4, and demineralized for 14 days in 4 M formic acid. The tissues were cryoprotected with 30% sucrose solution in 0.1 M PBS, pH 7.4, for 48 hrs, frozen-embedded, stored at -82°C, and frozen-sectioned on a cryostat at 30 µm in consecutive sections.
Determination of the Healthy and Inflamed Dentin-Pulp Complex
Hematoxylin and Eosin (H&E) Staining
To determine the health and inflammation state of the molars and premolars, we stained the sections by Hematoxylin and Eosin (H&E).
Confocal Microscopy Detection of CD3 and CD11b in Inflammatory Cells of the Inflamed Dental Pulp
In the irreversibly inflamed dental pulp, CD11b was used for identification of possible monocytes, granulocytes, and activated lymphocytes. For detection of the mature T-cells in the irreversibly inflamed dental pulp, CD3 was used.
The free floating sections were incubated with 10% NGS + 2% BSA for 30 min. Then, sections were incubated with mouse anti-CD11b (1:800) (eBioscience, San Diego, CA, USA) and CD3 (1:800) (eBioscience) antibodies overnight at 4°C. The sections were incubated with DyLight™ 488-conjugated goat anti-mouse IgG (1:500) for 1 hr at RT. For identification of the cell nuclei, the sections were incubated with chromatin marker DRAQ5™ at 1:2000 dilution for 10 min in the dark and coverslipped with Aqua Poly/Mount (Appendix 2).
The Antibodies against α1-, β1-, and α2-subunits of sGC
In earlier studies, generation of the antibodies against the α1- (Koglin and Behrends, 2003), β1- (Behrends et al., 2001), and α2-subunits (Bamberger et al., 2001) of sGC was described (Appendix 3).
Immunohistochemistry
The free-floating sections were incubated with 0.3% H2O2 in 0.05 M Tris-buffered saline (TBS) for 20 min to inhibit endogenous peroxidase. Non-specific immunoglobulin binding sites were blocked by incubation in 10% normal goat serum (Vector, Burlingame, CA, USA) + 2% bovine serum albumin (Sigma, St. Louis, MO, USA). The sections were incubated for 48 hrs with monoclonal antibodies against β-actin (1:2000) (Sigma), nitrotyrosine (1:800) (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and polyclonal antisera against iNOS (1:800) (Santa Cruz), sGC α1- (1:800), sGC β1- (1:800), and sGC α2-subunits (1:800) at 4°C. Then, the sections were incubated for 1 hr with biotinylated anti-mouse IgG (1:500) (Vector), anti-rabbit IgG (1:500) (Vector), and avidin-biotin peroxidase complex (1:100) (Vector). The immunohistochemical reaction was developed in all quantified incubations for 5 min with 0.05% 3,3′-diaminobenzidine tetrahydrochloride (Sigma) in 0.05 M Tris-HCl buffer, pH 7.6, containing 0.01% H2O2 and 0.01% nickel sulfate. In immunohistochemical controls, sections were separately incubated in the absence of the primary or secondary antisera.
Quantification of Immunohistochemistry
The densitometric staining intensities of the β-actin, nitrotyrosine, iNOS, α1-, α2-, and β1-subunits of sGC in odontoblasts of the healthy and in inflamed sections were measured by grey values of immunostaining (Appendix 4).
Statistical Analysis
Statistical comparisons between healthy and inflamed tissues for antibodies were performed with two-tailed Student’s t test for paired samples, as implemented in the software package SPSS for Windows, Version 18.0. Significance was considered at a p value ≤ 0.05.
Results
Histology of Cells in the Healthy and Inflamed Human Dentin-Pulp Complex
In the healthy dentin-pulp complex, H&E staining revealed a preservation of pulp stroma cells (Fig. 1A), blood vessels (Fig. 1A), nerve fibers (Fig. 1A), and odontoblasts (Fig. 1C) in a typical tissue architecture of the dentin-pulp complex. In the case of the inflamed dentin-pulp complex, a strong, diffuse pulpal and perivascular lymphocytic infiltrate was detected (Fig. 1B). In the inflamed dentin-pulp complex, stromal cells (Fig. 1B), odontoblasts, and pulpal tissue damage were identified (Fig. 1D). Beneath caries lesions, odontoblast cell bodies and their processes were detected with a destruction depending on the degree of the caries lesions in the dentin-pulp complex. In the inflamed dental pulp, the numerous blood vessels were filled with erythrocytes in different widely diameters (Figs. 1B, 1D).
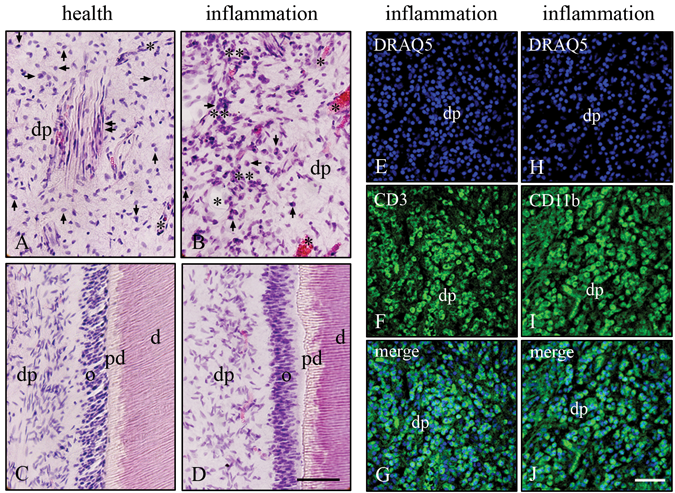
Histology of cells in the healthy and inflamed dentin-pulp complex. Healthy sections stained with Hematoxylin and Eosin (H&E) revealed dental pulp stroma cells (arrows), blood vessels (asterisks), nerve fibers (double arrows), and odontoblast cell bodies and their nuclei with a normal cell architecture
Detection of CD11b and CD3 in Cells of the Irreversibly Inflamed Dental Pulp
In the irreversibly inflamed human molar dental pulp, the nuclei were identified by DRAQ5 staining (Figs. 1E, 1H). In the numerous inflammatory cells (possible monocytes and activated lymphocytes), CD11b was detected (Fig. 1F). In the irreversibly inflamed human dental pulp of the same human molar, numerous mature T-cells were identified by CD3 immunoreactivity (Fig. 1I). Co-localizations of DRAQ5 in nuclei of the CD11b- (Fig. 1G) and CD3- (Fig. 1J) positive inflammatory cells were detected.
Quantitative Immunohistochemistry
β-actin, 3-nitrotyrosine, and iNOS in Odontoblasts of the Healthy and Inflamed Dentin-Pulp Complex
In odontoblasts of the healthy dentin-pulp complex, β-actin was detected (Figs. 2A, 2G). In comparison with the healthy odontoblasts, inflammation induced a non-significant increase in staining intensity for β-actin in odontoblasts (Figs. 2B, 2G). Immunoreactivity of 3-nitrotyrosine was absent in healthy odontoblasts (Figs. 2C, 2G), while a strong immunoreactivity for 3-nitrotyrosine was detected in odontoblasts of the inflamed dentin-pulp complex (Figs. 2D, 2G). In odontoblasts of the healthy dental pulp, only single odontoblasts were weakly positive for iNOS (Figs. 2E, 2G). In comparison with the healthy odontoblasts (Figs. 2E, 2G), the signal intensities of iNOS were detected with a higher increase in odontoblasts of the inflamed dentin-pulp complex (Figs. 2F, 2G).
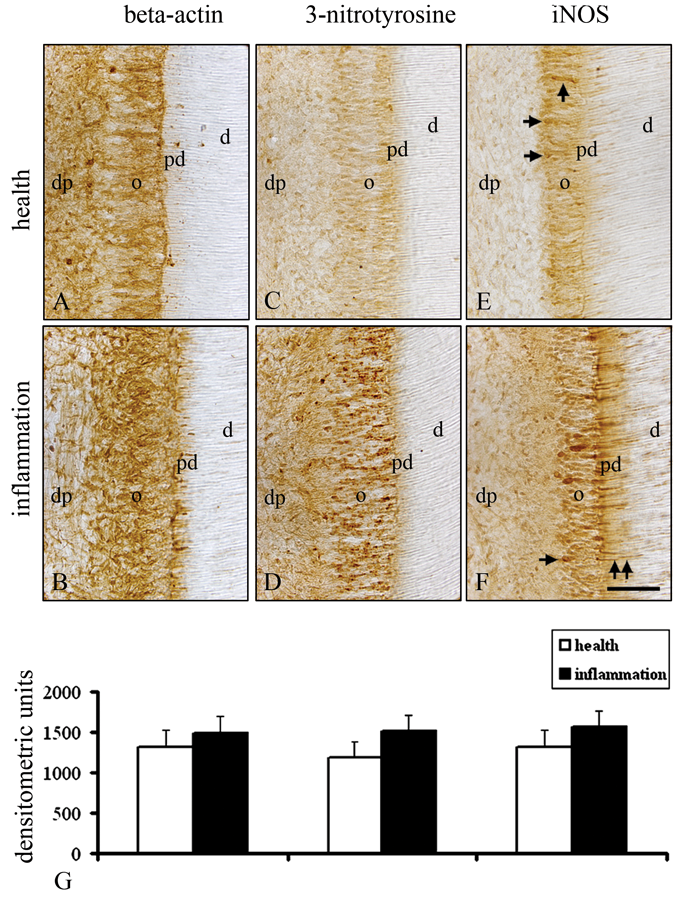
Immunohistochemical protein levels of β-actin, 3-nitrotyrosine, and iNOS in the healthy and inflamed human molar odontoblasts. A moderate immunoreactivity for β-actin in healthy odontoblasts (
In immunohistochemical controls, no immunoreactivity was detected in the incubations without primary or secondary antibodies (data not shown).
The α1-, β1-, and α2-subunits of sGC in Odontoblasts of the Healthy and Inflamed Dentin-Pulp Complex
In the healthy odontoblasts of the crown pulp, an immunohistochemical staining was detected for the α1-subunit (Figs. 3A, 3G), but the signal intensities of the α1-subunit in odontoblasts of the inflamed dental pulp were significantly decreased (Figs. 3B, 3G). In comparison with the higher staining intensity for the β1-subunit in odontoblasts of the healthy dental pulp (Figs. 3C, 3G), inflammation of the dentin-pulp complex induced a decrease in the immunostaining for the β1-subunit in odontoblasts (Figs. 3D, 3G). The α2-subunit was detected with a higher staining intensity in odontoblasts at the crown pulp areas of the healthy dentin-pulp complex (Figs. 3E, 3G). Inflammation of the dental pulp resulted in a significant decrease in the signal intensities for the α2-subunit in odontoblasts (Figs. 3F, 3G).
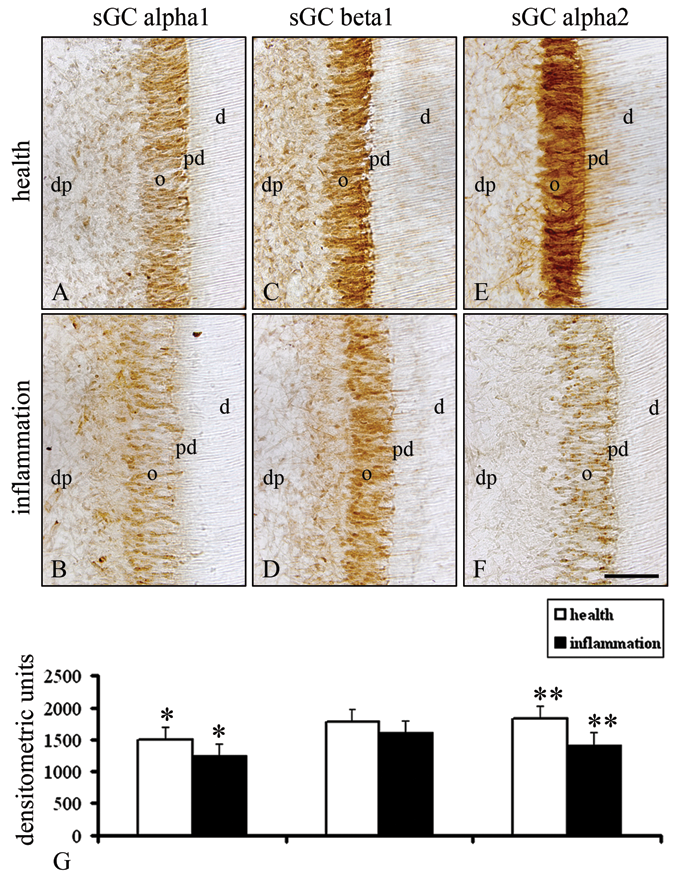
Immunohistochemical protein levels of the α1-, β1-, and α2-subunits of sGC in the healthy and inflamed human molar odontoblasts. In comparison with the higher staining intensities for the α1-subunit in healthy odontoblasts (
In immunohistochemical controls, no staining was detected in the separate incubations without primary or secondary antibodies (data not shown). The immunohistochemical localization of the α1-, β1-, and α2-subunits of sGC in blood vessels of the human (data not shown) and rat molar pulps (Korkmaz et al., 2005) served as positive controls.
Discussion
Treatment of the rat molar dentin-pulp complex with the NO donor NONOate increased cGMP staining intensities in odontoblasts, while the NO synthase inhibitor L-NAME attenuated the staining intensities for cGMP, suggesting an effect of NO on the activation of sGC in odontoblasts (Korkmaz et al., 2005). In the present study, quantitative immunohistochemical analysis revealed a decrease in the protein levels of the α1-, β1-, and α2-subunits of sGC in inflamed odontoblasts of the human molar dentin-pulp complex. Analysis of these data indicates a critical involvement of sGC in odontoblasts during health.
In odontoblasts of the inflamed dentin-pulp complex, induction of iNOS may be the source of NO and the high concentrations of ONOO- required for extensive nitrosylation of tyrosine residues in odontoblasts. In the present findings, the higher expression of iNOS in the inflamed odontoblasts was also supported by the higher amount of 3-nitrotyrosine in inflamed odontoblasts. In the inflamed cells, higher levels of ROS and reactive nitrogen species (RNS) have been described (Nathan, 2006). The higher levels of ROS decreased the sGC activity associated with down-regulation of the protein expression of α1- and β1-subunits of sGC (Ruetten et al., 1999). The enzymatic function of sGC is also decreased in the presence of excess levels of ONOO- (Weber et al., 2001; Stasch et al., 2006). An increased level of ROS is associated with oxidation of the β1-subunit prosthetic heme group that converts sGC to an NO-insensitive state (Stasch et al., 2006). Under inflammatory conditions, sGC is not sensible to activation by NO (Meurer et al., 2009). In the presence of excess levels of ROS, NO-dependent S-nitrosylation of a β1-subunit-thiol resulted in decreased responsiveness of sGC to NO (Sayed et al., 2007). The high concentrations of ROS and RNS in inflamed odontoblasts can impair the activity of sGC or induce degradation in the α1-, β1-, and α2-subunits of sGC at the protein level. An inflammation-dependent decrease in the protein levels of the sGC in odontoblasts occurs in a subunit-specific manner. While the decrease of the β1-subunit in inflamed odontoblasts was moderate, the α1- and α2-subunits were much more decreased in odontoblasts of the inflamed dentin-pulp complex. This may contribute to the reduced biological availability of the possible α1β1- and α2β1-heterodimer of sGC as receptor enzymes to transmit NO effects in odontoblasts. Analysis of the present data gives further evidence that sGC could also be regulated at the transcriptional level.
There was a significant discrepancy between clinical and histological diagnoses for the inflamed and non-inflamed teeth used in the present study. In addition to the H&E staining, in the inflamed dentin-pulp complex, CD11b and CD3 were used as markers for cells of inflammation in the dentin-pulp complex. The CD11b- and CD3-positive cells were detected only in sections of the inflamed dentin-pulp complex. In the case of irreversible dental pulp inflammation, migration of dendritic cells to the odontoblast layer and accumulation of T-lymphocytes, neutrophils, and B-lymphocytes in the local inflammation areas are associated with the development of the inflammatory processes in the dentin-pulp complex (Goldberg et al., 2008). It has been reported that cytokines—for example, interleukin-1α, tumor necrosis factor-α (Takata et al., 2001), interleukin-1β, and bacterial LPS (Pedraza et al., 2003; Glynos et al., 2007; Baltrons et al., 2008)—decrease sGC in an NO-dependent but also NO-independent manner (Sardon et al., 2004). In addition to the ROS and RNS, under inflammatory conditions, the decreased protein levels of the α1-, β1-, and α2-subunits of sGC in odontoblasts may also be caused in part by inflammatory mediators.
Here, we have shown that odontoblasts of human molars contain α1-, β1-, and α2-subunits of sGC. We have found that inflammation of the dentin-pulp complex induces a decrease at the protein levels of the α1-, β1-, and α2-subunits of sGC in odontoblasts. Inflammation-dependent production of ROS, RNS, and inflammatory mediators may be involved in the down-regulation of the α1-, β1-, and α2-subunits of the sGC in odontoblasts. It is concluded that sGC is a critical enzyme to mediate the different biological effects of NO in healthy odontoblasts.
Footnotes
Acknowledgements
The study was supported by the Forschungskommission of the Heinrich-Heine-University of Düsseldorf.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.