
Abstract
Nitric oxide (NO) and reactive oxygen species (ROS) are key molecules in resistance to pathogens. Little is known about their role in pathogenesis of periapical lesions. To address this issue, we induced periapical lesions in mice lacking nitric oxide synthase (iNOS-/-) or phagocyte oxidase (PHOX-/-). iNOS-/- mice expressed higher levels of IL-1β, TNF-α, RANK, RANKL, and MCP-1 than C57BL/6 and PHOX-/-. Apical thickening of the periodontal ligament was also greater in iNOS-/- compared with other groups. Interestingly, ROS production did not interfere in periapical lesion progression, but seemed to be essential for the appearance of multinucleated TRAP-positive cells. Thus, periapical lesion progression in iNOS-/- was associated with an imbalance of pro-inflammatory cytokines (IL-1β and TNF-α), bone-resorptive modulators (RANK and RANKL), and MCP-1. We conclude that NO, but not ROS, controls progression of bone resorption in a murine experimental model of apical periodontitis.
Keywords
Introduction
Apical periodontitis is characterized by periapical bone destruction and occurs as a result of host defense reactions primarily caused by root canal infection. Progression of periapical lesion depends on bone resorption induced by interleukin (IL)1-β, tumor necrosis factor (TNF)-α, and the receptor activator for NF-κB ligand (RANKL) (Vernal et al., 2006; Kawashima et al., 2007).
RANKL plays an important role in bone resorption around tooth apex. Since this cytokine is required for differentiation and activation of osteoclasts, RANKL-deficient mice are osteopetrotic (Kong et al., 1999). Osteoprotegerin (OPG) is a natural decoy receptor for RANKL. Therefore, a balance between OPG and RANKL regulates osteoclast differentiation (Hofbauer and Heufelder, 2001).
Monocyte chemoattractant protein-1 (MCP-1) is one of the inflammatory factors that mediate extravasation and accumulation of leukocyte subsets during inflammation. MCP-1 recruits osteoclast monocyte precursors and synergizes with RANKL to induce osteoclastogenesis, particularly enhancing cell fusion (Li et al., 2007). Thus, MCP-1 is one of the inflammatory mediators responsible for periapical bone resorption.
Nitric oxide (NO) is a free radical produced by inflammatory macrophages and other cell types. NO plays an important role in protection against infection, but may cause tissue damage (Leonard et al., 1998). There is a correlation between development of periodontal disease and activation of inducible nitric oxide synthase (iNOS), which is induced by interferon (IFN)-γ, IL-1β, TNF-α, and bacterial products (Xie et al., 1992; Lappin et al., 2000). NO production plays an important role in periapical lesion development (Takeichi et al., 1998). In periapical lesions, macrophages and polymorphonuclear leukocytes are the main sources of NO (Hama et al., 2007). Moreover, osteoblasts and osteoclasts can express iNOS, which may indicate that NO is important for periapical lesion progression.
In adult bone, the effects of iNOS induction appear to be complex. NO produced endogenously or supplied by NO donors has antagonistic actions that profoundly affect the recruitment, proliferation, differentiation, activity, and survival of osteoclasts and osteoblasts (Kasten et al., 1994; Holliday et al., 1997). Additionally, some investigators have shown that high concentrations of exogenously administered NO can induce apoptosis in osteoclasts and osteoblasts (Armour et al., 2001). However, endogenous NO can stimulate osteoclasts, and in vivo experiments indicate that NO is required for proper osteoclast and osteoblast function (Cuzzocrea et al., 2003).
NO has inhibitory effects on RANKL expression (Fan et al., 2006). Skeletal loading promotes inhibition of bone resorption by suppression of RANKL, while concurrently causing the generation of NO (Rubin et al., 2006). Moreover, RANKL suppression was dependent on NO production (Rahnert et al., 2008). Importantly, NO has a potent inhibitory effect on osteoclast differentiation by down-regulating RANKL signaling (Fan et al., 2004). Also, RANKL-induced iNOS functions as a negative feedback signal to limit osteoclastogenesis (Zheng et al., 2006).
NADPH oxidase (NOX2) is a multi-subunit enzymatic complex that generates superoxide anion (O2 .-), composed of 2 membrane subunits (gp91 and p47) and 3 cytoplasmatic subunits (Babior, 2004). The generation of superoxide anion by NOX2 is crucial to the host defense, especially the antimicrobial activity of professional phagocytes. High levels of superoxide anion induce other reactive oxygen species (ROS) that are responsible for antimicrobial activity (Babior, 2004). Low concentrations of ROS can act as second-messenger molecules in signaling pathways other than those involved in microbicidal activity (Lander, 1997; Rhee, 1999). One of these signaling pathways is related to the transcriptional factors involved in osteoclast differentiation. Hence, some authors have demonstrated the importance of NOX2 activation and ROS production in bone resorption and osteoclast differentiation by showing that osteoclastogenesis is inhibited by the treatment of pre-osteoclast cells with superoxide dismutase and catalase (Yang et al., 1998). In support of this finding, other researchers showed the importance of ROS in RANK-mediated signaling (Koh et al., 2006; Sasaki et al., 2009a). For this reason, ROS production would likely be involved in osteoclast differentiation and bone resorption.
To investigate the role of NO and ROS in the development of periapical lesions, differentiation of osteoclasts, and cytokine production at the site of infection, we examined the effects of root canal infection in wild-type, iNOS-/-, and PHOX-/- mice.
Materials & Methods
Animals
C57BL/6 mice aged 6 to 8 wks were purchased from CEBIO (UFMG, Belo Horizonte, MG, Brazil). Knockout mice for NADPH oxidase (PHOX-/-, B6.129S6-Cybb tm1Din /J) and for nitric oxide synthase 2 (iNOS-/-, B6.129P2-Nos2 tm1Lau /J) in the C57BL/6 background were purchased from Jackson Laboratories (Bar Harbor, ME, USA). Mice were kept in a conventional animal house with barriers and a controlled light cycle. The experimental protocol was approved by the animal ethical committee (protocol 234/08, CETEA, UFMG).
Periapical Lesion Induction
Experimental procedures were carried out with the animals under general anesthesia, 100 mg/kg of ketamine hydrochloride (Dopalen, Division Vetbrands Animal Health, Jacareí, SP, Brazil) and 10 mg/kg of xylazine (Anasedan, Agribrands do Brasil Ltda, Paulínia, SP, Brazil). Animals were fixed on a table that permitted direct visualization of the upper first molar. The pulp of the upper right first molar was accessed with the use of an endodontic operative microscope (Alliance, São Paulo, SP, Brazil) and a ½ carbide bur (KG Sorensen, Barueri, SP, Brazil) coupled to a controlled rotation handpiece (Driller, São Paulo, SP, Brazil). The pulp chamber was opened until the entrances of the canals could be visualized and probed with a no. 8 endodontic K file. There were no surgical interventions on the upper left first molar, which was our control. Exposed teeth were left open to the oral environment for 14 days (n = 6 mice).
Histomorphometric Evaluation
All animals were sacrificed by cervical dislocation, and the hemi-maxillas were immersed in 10% buffered neutral formalin for 72 hrs at room temperature. Demineralization and histological routine were performed as described (Silva et al., 2008). The apical region was identified with the apical foramen as reference. The samples were randomized and blindly examined. Quantification of periapical bone resorption was evaluated as the size of the apical region. Images were obtained from the periapical region through a 10x objective and a JVC TK-1270-RGB adapted to a microscope, and traced and analyzed with KS300 software built into a Kontron Elektronick/Carl Zeiss image analyzer. We analyzed unexposed teeth to determine the periodontal ligament area. The periapical lesion surrounding the apical foramen was traced and measured.
TRAP Analysis
Tartrate-resistant acid phosphatase (TRAP) activity was detected with the use of a leukocyte-specific acid phosphatase kit (Sigma Aldrich, St. Louis, MO, USA). The sections were stained with TRAP solution for 1 hr at 37°C and counter-stained with Mayer’s hematoxylin. Multinucleated cells on the bone superficies that were stained dark red to purple were considered TRAP-positive.
Real-time Polymerase Chain-reaction Analysis
Quantitative polymerase chain-reaction analysis (RT-PCR) of IL-1β, TNF-α, RANKL, RANK, OPG, and MCP-1 mRNA expression was performed in tissue samples. Primer sequences are listed in the Table. The periapical tissue surrounding root apices of the right and left molars was extracted together with the surrounding bone. Gingiva, oral mucosa, and crown were removed and discarded. Samples were treated with TRIZOL (Invitrogen, Carlsbad, CA, USA). Reverse transcription was conducted as described (Silva et al., 2008). RT-PCR was performed in an ABI PRISM® 7900HT Sequence Detection System (Applied Biosystems, Foster City, CA, USA). We used Syber-Green system detection (Applied Biosystems) to assay amplification. We used Sequence Detection Software version 1.3 (Applied Biosystems) to analyze data after amplification. Results were obtained as threshold cycle (Ct) values. Ct values were normalized with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene expression, following the equation 2-ΔΔCt (Schmittgen and Livak, 2008). Mean values of 3 independent measurements are reported.
Sequences of the Primers Used for Analysis of IL-1β, TNF-α, RANK, RANKL, OPG, and MCP1 mRNA Expression by Real-time PCR

FW, forward primer; RV, reverse primer.
Amplicon length in base pairs.
Statistical Analysis
The SPSS statistical program was used for analysis (SPSS Inc., Chicago, IL, USA). We applied the Kolmogorov-Smirnov test to determine normal distribution. The mean values obtained from morphometric analysis were compared between and among strains, with the independent t test. We used Levene’s test to assay the equality of variance. Means were considered different if P < 0.05.
Results
C57BL/6 control and experimental periapical areas were similar 14 days after the procedure (Figs. 1A, 1D, P > 0.05). Exposed teeth from iNOS-/- mice presented intense inflammatory infiltration and bone resorption compared with the other strains (P < 0.05, Figs. 1B, 1D). Periapical lesions were attenuated in PHOX-/- mice compared with iNOS-/- (P < 0.05) (Figs. 1C, 1D). Tissues from C57BL/6 and iNOS-/- mice presented TRAP-positive cells (Figs. 1E, 1F), but an increase in TRAP-positive cells due to the experimental procedure was found only in iNOS-/- exposed teeth (P < 0.05) (Fig. 1H). Remarkably, TRAP-positive cells were not clearly detected in the periapical lesion or in the periodontal tissue of PHOX-/- mice (Figs. 1G, 1H).
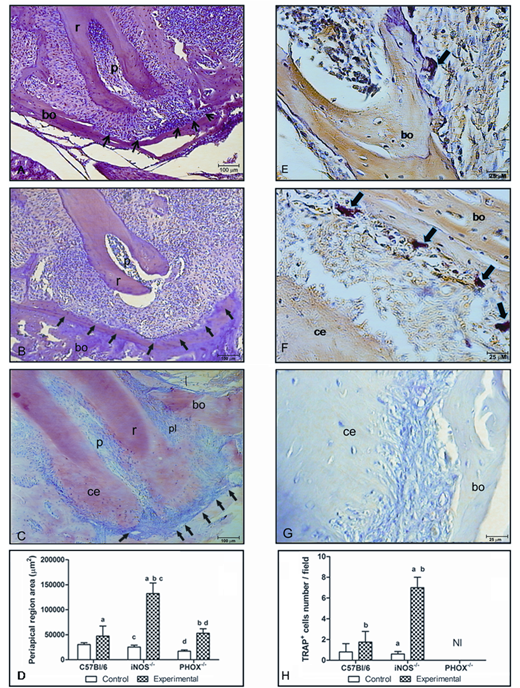
Photomicrograph of periapical inflammatory lesions, delimited by arrows, and TRAP-stained cells, indicated by large arrows, in C57BL/6
IL-1β, TNF-α, RANKL, RANK, and MCP-1 were up-regulated in the apical tissue of exposed teeth from all groups (P < 0.05), and were higher in iNOS-/- mice than in C57BL/6 and PHOX-/- (P < 0.05) (Figs. 2A, 2B, 3A, 3B). OPG expression showed a broad variation among and inside the groups, leading to high standard deviation values (Fig. 3C). MCP-1 mRNA expression was higher in exposed apical tissue from iNOS-/- mice, compared with C57BL/6 and PHOX-/- (Fig. 3D). Also, MCP-1 mRNA expression was 10-fold higher in iNOS-/-- exposed periapical tissues than in control tissues (P < 0.05). No difference in MCP-1 expression between control and experimental C57BL/6 and PHOX-/- mice was found (P > 0.05).
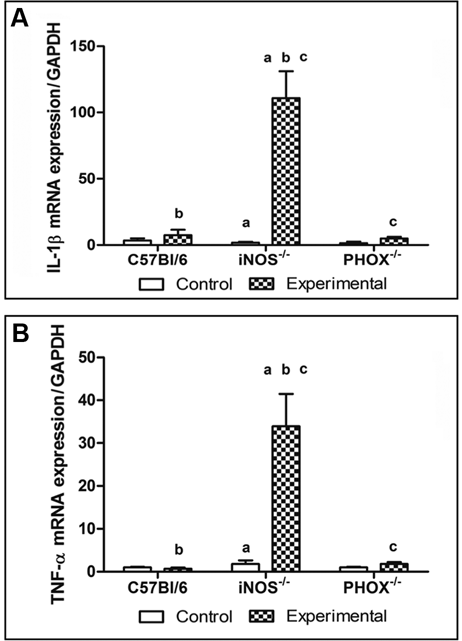
IL-1β and TNF-α mRNA expression in the periapical lesion of C57BL/6, iNOS-/-, and PHOX-/-. Data are expressed as the mean ± SE for 3 independent experiments, each with 6 mice per group.
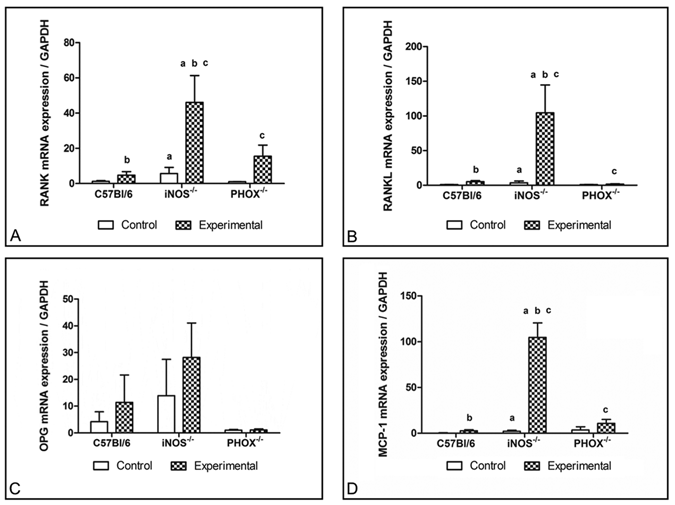
RANK, RANKL, OPG, and MCP-1 mRNA expression by cells in the periapical lesions of C57BL/6, iNOS-/-, and PHOX-/- mice. Data are expressed as the mean ± SE for 3 independent experiments with 6 mice per group.
Discussion
We evaluated the role of NO and ROS in the pathogenesis of periapical disease in wild-type, iNOS-/-, and PHOX-/- mice. iNOS-/- mice, but not PHOX-/- mice, displayed aggravated and increased extension of apical lesions, including bone resorption. Previous studies have shown an increase in iNOS expression in human apical lesions (cysts and granulomas), suggesting that iNOS is one of the many factors responsible for the inhibition of the progression of periapical lesions (Kawashima et al., 2007; Lin et al., 2007). iNOS was expressed by macrophages and endothelial and plasma cells (Hama et al., 2007). There are no data on the role of ROS in periapical lesions.
Progression of the periapical lesion is related to two events: production of the pro-inflammatory cytokines IL-1β, IL-6, and TNF-α; and cellular migration and osteoclast differentiation. We investigated both events. IL-1β and TNF-α mRNA expressions were remarkably up-regulated in iNOS-/- mice compared with wild-type and PHOX. Macrophages and neutrophils exhibit bactericidal activity by iNOS-mediated NO production (Fierro et al., 1999; Gyurko et al., 2003), which would be absent in iNOS-/- mice. Also, it is well known that NO regulates the immune system by down-regulating IL-12 synthesis, which is a potent inducer of Th1 differentiation (Huang et al., 1998), thereby blocking the amplification or the inflammatory circuit created with macrophages and IFN-γ. Consequently, deficiency of NO increases IFN-γ production, thereby inducing osteoclast activity (Gao et al., 2007). In addition, bacterial products such as LPS stimulate IL-1β and TNF-α production, thereby inducing osteoclast activity (Schletter et al., 1995). Therefore, the absence of iNOS and the reduction of NO production could both cause an increase of bacterial products in the periapical region and an increase in the inflammatory response, leading to the production of higher levels of IL-1β and TNF-α.
NO can either stimulate or suppress osteoclast differentiation and bone resorption in murine cell cultures (Cuzzocrea et al., 2003; Lee et al., 2004). iNOS deficiency or NO pharmacological inhibition increased osteoclast differentiation in murine and rat cell cultures by regulating RANKL expression in bone marrow stromal cells (Ralston and Grabowski, 1996; Holliday et al., 1997; Fan et al., 2004; Zheng et al., 2006). Our present findings show that, in the periapical region, NO is key to bone resorption during the inflammatory process by down-regulating the generation of osteoclasts. This conclusion is supported by the significantly enhanced expression of RANK/RANKL mRNA and osteoclast numbers in iNOS-/- compared with C57BL/6. OPG mRNA expression was not different between iNOS-/- and C57BL/6 mice. However, our findings suggest a tendency for iNOS-/- animals to express more OPG compared with wild-type.
NO modulates the expression of chemokines, including MCP-1, by endothelial cells (Desai et al., 2003). In agreement with these data, iNOS-deficient mice expressed more MCP-1 when compared with wild-type.
ROS production by PHOX is involved with the intracellular signaling mediated by RANKL during osteoclast differentiation (Sasaki et al., 2009a,b). The role of PHOX in oral diseases is currently unknown. Our results demonstrated that periapical lesion development in PHOX-/- mice was similar to that in wild-type with respect to bone resorption. Expression of RANK and RANKL was similar between wild-type and PHOX-/- mice, and, yet, we could find TRAP+ cells only in wild-type tissues. The absence of these cells in PHOX-/- mice may be related to an impairment of RANK-RANKL activation as a result of no ROS production. Our findings are in agreement with those of other studies that demonstrated the participation of ROS in RANKL-mediated osteoclastogenesis (Lee et al., 2005; Koh et al., 2006). In addition, p22 phox and p67 phox gene knockdown inhibited multinucleated TRAP+ cell formation in RAW 264.7 cells (Sasaki et al., 2009a).
NO and superoxide may react in vivo to produce peroxynitrite. Peroxynitrite promotes the nitration of phenolic compounds such as tyrosine, which may lead to molecular (and tissue) damage (Radi et al., 2001). Moreover, peroxynitrite has immunomodulatory effects (Brito et al., 1999). Hence, peroxynitrite could have an effect on the development of periapical lesions. However, since our mouse models exclude either NO or superoxide by the deletion of enzymes key to the production of these species, it is possible that either peroxynitrite plays no role in the development of the periapical lesion or its role is compensated for by unknown mechanisms in the enzyme-deficient strains.
In conclusion, this study points to an important role for iNOS-derived NO in controlling progression of bone resorption in a murine experimental model of apical periodontitis. iNOS deficiency is associated with an imbalance in the pro-inflammatory cytokines (IL-1β and TNF-α), bone-resorptive modulators (RANK and RANKL), and the chemokine MCP-1. Interestingly, ROS production does not appear to interfere in periapical lesion progression and may mediate osteoclast differentiation.
Footnotes
Acknowledgements
We are indebted to Antonio Mesquita Vaz for excellent animal care and Dr. Rosa Esteves Arantes for invaluable contributions. LMAS, APRS, and LQV are CNPq fellows. This study was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq473597/2006-3) and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior). The authors are members of INCT de Processos Redox em Biomedicina-Redoxoma (CNPq/FAPESP/CAPES 573530/2008-4). The authors declare that there is no conflict of interest or funding.