
Abstract
Mesenchymal stem cells are present in the dental pulp. They have been shown to contribute to dentin-like tissue formation in vitro and to participate in bone repair after a mandibular lesion. However, their capacity to contribute efficiently to reparative dentin formation after pulp lesion has never been explored. After pulp exposure, we have identified proliferative cells within 3 zones. In the crown, zone I is near the cavity, and zone II corresponds to the isthmus between the mesial and central pulp. In the root, zone III, near the apex, at a distance from the inflammatory site, contains mitotic stromal cells which may represent a source of progenitor cells. Stem-cell-based strategies are promising treatments for tissue injury in dentistry. Our experiments focused on (1) location of stem cells induced to leave their quiescent state early after pulp injury and (2) implantation of pulp progenitors, a substitute for classic endodontic treatments, paving the way for pulp stem-cell-based therapies.
Keywords
Dental pulp trauma or infections are classically treated by pulp capping, intended to promote pulp healing when the lesion involves a limited part of the coronal pulp while a large part of the pulp is still alive. This treatment normally leads to the formation of a reparative dentinal bridge (Fitzgerald et al., 1990). However, the presence of tunnels with persisting pulp remnants and the fact that a poorly mineralized osteodentin is generally formed explain the high percentage of failures observed with this therapy (Barthel et al., 2000). In cases of pulp degradation and necrosis, endodontic treatment of the root canal is needed. Despite an apparent initial success, treated teeth become susceptible to re-infection and fracture. The high failure rate of both treatments (up to 18.8%) (Willershausen et al., 2005) ends with tooth loss. This has a strong economic impact for public health. Therefore, active research into alternative treatments is ongoing.
The ability of the dentin-pulp complex to respond to a variety of pathological conditions and injuries by deposition of a reparative dentin by pulp “progenitors” has long been recognized (Tziafas et al., 2000). However, the cellular and molecular events underlying this repair capacity of dental pulp cells are still largely unknown. In particular, the origin and precise identity of the progenitors have remained elusive. Rouget’s pericytes (perivascular cells), undifferentiated mesenchymal/mesectodermal cells, or fibroblasts have all been proposed as potential progenitors mediating pulp repair after destruction of the odontoblasts and the Hoehl’s sub-odontoblastic cell layer (Fitzgerald et al., 1990). The first descriptions, in the early 21st century, of the presence of cell populations displaying stem cell properties in the perivascular area of the human adult tooth (Gronthoset al., 2000; Miura et al., 2003), have opened new paths for the consideration of more conservative therapies. A formal demonstration that the “stem” cells present in the pulp are the cells performing the dentin repair is still lacking. An involvement of stem cells carried by the circulation to the lesion site cannot be excluded. Identifying cells mobilized in response to pulp injury is a prerequisite to the design of alternative strategies for capping and endodontic treatment, with stem cells. Two strategies can be envisioned, depending on the types of pulp lesions. When parts of a damaged pulp remain alive, a pulp-capping procedure performed with a biomolecule able to recruit quiescent stem cells would permit their proliferation and differentiation into odontoblast-like cells capable of forming a dentinal bridge. In more severe lesions, where the pulp is irreversibly damaged, direct implantation of exogenous “stem” cells into the remaining pulp would be performed, with the aim of regenerating a dental pulp leading to the formation of a mineralized matrix.
Over the past few years, our laboratories have isolated pulpal progenitor cell lines (Priam et al., 2005) and have been involved in characterizing the cellular and molecular mechanisms implicated in these different strategies. In this paper, we will (i) review the data on localization of proliferative cells recruited within the pulp 48 hrs after pulp exposure, and (ii) analyze how implanted pulp-derived stem cells have the ability to promote tooth repair.
Localization of Proliferative Cells After Pulp Exposure
Our aim was to trace cells which enter the cell cycle 48 hrs after pulp exposure, with or without the concomitant introduction of biomolecules. This approach is driven by the well-established capacity of biomolecules to improve dental repair by acting on dentinogenic cells. Indeed, extracellular matrix (ECM)-associated molecules may play promiscuous roles in dentin formation. By participating in cell-ECM-dependent signaling (Bornstein and Sage, 2002), biomolecules, such as amelogenin splice products (Gruenbaum-Cohen et al., 2009), may control odontoblast functions and/or influence stem cell recruitment. Our studies with the implantation of agarose beads loaded with amelogenin small peptides, A-4/LRAP and A+4, after a pulp exposure of the first molar in rats, have shown that these molecules were capable of promoting, within a month, the formation of a dentinal bridge. Therefore, they can be considered as good candidates to be used as new capping agents (Goldberg et al., 2009). These experiments have shown that the repair process is initiated through a phase of cell proliferation during the first week following implantation. Thereafter, odontoblast cell differentiation, as revealed by DSP expression, occurs near the pulp exposure, leading to reparative dentin formation (Six et al., 2004). The nature of the “recruited” proliferative cells, however, has not been determined.
The simplest scheme would assume that mitotic cells correspond to “stem” cells recruited to perform the repair process in response to injury. Forty-eight hrs after pulp exposure, proliferative cell nuclear antigen (PCNA)-positive cells were localized in 3 “zones” (Figs. 1B, 1C, 1E, 1F). In the crown, zone I was located in the mesial part of the pulp near the exposure site (Fig. 1A), whereas zone II was located at the junction between the mesial and central parts of the pulp (Fig. 1A). Zone I is adjacent to the inflammation area generated by the surgical procedure, where the repair process will take place after 15-28 days, between the coronal pulp and the upper pulp part of the root (Six et al., 2004). Zone II is where an osteodentin structure will be formed, between the mesial and central parts of the pulp after the same period of time. However, this junction is a specificity of the rat molar (Weber and Ashrafi, 1981; unpublished observations). In the mesial root, zone III was localized within the pulp, close to the apex (Fig. 1D).
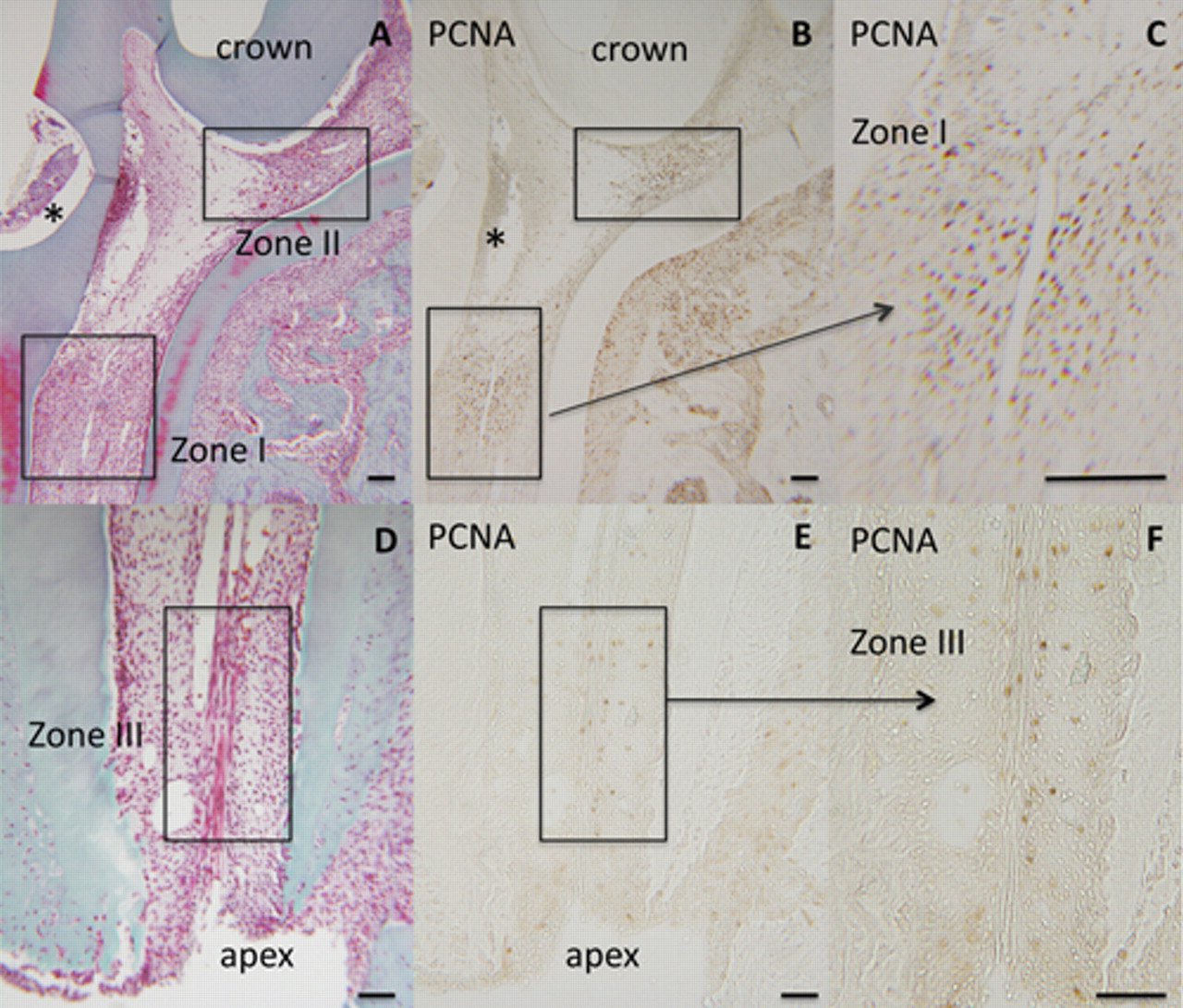
Localization of proliferative cells in the pulp of a rat first molar 48 hrs after exposure, as revealed by proliferative cell nuclear antigen (PCNA) immunostaining. Light microscope images of Masson’s trichrome-stained paraffin sections show the coronal (
Interestingly, when amelogenin small peptides (A-4/LRAP)-loaded beads were implanted into the exposed pulp of the rat first molar, no significant difference in the distribution and number of the PCNA-positive cells was observed at 48 hrs as compared with controls (pulp exposure with or without the implantation of unloaded beads). Thus, A-4 does not mobilize subsets of progenitors within the pulp other than those recruited by the trauma. This suggests that A-4 has no impact on the early events related to the recruitment of mitotic cells, but acts as a bioactive molecule promoting the differentiation potential of odontogenic cells and improving reparative dentin formation.
What is the nature of these proliferating cells? Odontogenic stem cells, pulpoblasts, or inflammatory cells? PCNA-positive cells defining zones I and II, close to the exposure site, may be mobilized by inflammatory molecules and/or mechanotransduction pathways generated by the lesion. Detection of PCNA-positive cells close to the apex in zone III reveals that a recruitment of mitotic cells occurs at a distance from the lesion. Of note, mitosis is not observed in this zone in the absence of pulp exposure. Whether these proliferative cells, distributed along or near the apical vessels, may have a perivascular origin (Shi and Gronthos, 2003) or whether they emanate and migrate from the periapical niche (Sonoyama et al., 2006) is still an open question.
We performed a preliminary immunohistochemical analysis on sections of injured teeth to analyze whether we could distinguish between “odontogenic stem cells” and “inflammatory cells” within the 3 zones of PCNA-positive cells.
We used antibodies against MHC IIB, identifying mainly dendritic cells, and against CD68, labeling monocytes/macrophages, whereas antibodies against α-SMA (smooth-muscle actin) and CD90 were used to screen non-inflammatory cells. CD90 (also named THY-1) can be used as a marker for a variety of stem cells, including mesoblastic and mesectodermal cells. A small number of precursor cells expressing CD 90 have been recently detected within the pulp of mouse molars (Balic et al., 2010). Accordingly, few CD90-positive cells can be visualized throughout the coronal and radicular pulp, and no change in their number and/or distribution could be observed 48 hrs after pulp exposure (not shown).
Cells labeled with anti-CD68 and MHC IIB antibodies were mainly visualized in the inflammatory area beneath the exposure site (Figs. 2D, 2E, 2G, 2H), adjacent to the proliferative zones I and II. Of note, only a few CD68- and MHC IIB-positive cells are present among the population of PCNA-positive cells (Figs. 2A, 2B), indicating that dividing cells did not have an inflammatory origin. In addition, α-SMA labeling was restricted to sparse perivascular cells in zones I and II (Figs. 2J, 2K) and also did not overlap with the PCNA-positive cells (Figs. 2A, 2B). Moreover, 48 hrs after lesion creation, we did not observe any increase in the distribution of α-SMA-positive cells as compared with that in control teeth, implying that mitotic cells do not arise from the vascular compartment.
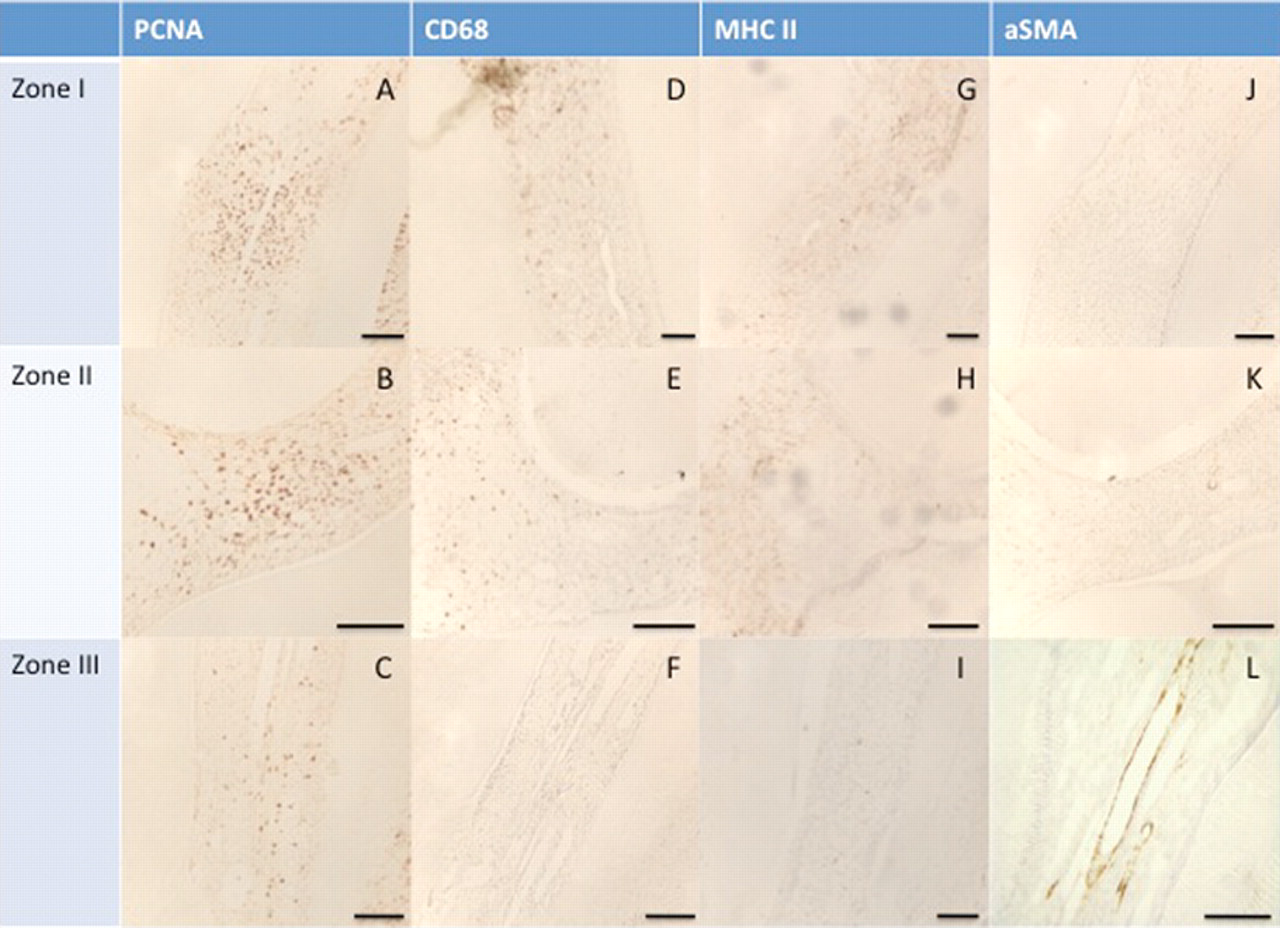
Comparative immunohistochemical analysis of the cell proliferation zones 48 hrs after pulp exposure. Proliferating cells were detected by PCNA immunostaining in zone I (
These observations lend support to the idea that the inflammatory process promotes a local secretion of cytokines and growth factors that induces rapid recruitment of progenitor cells resident in the pulp stroma in the vicinity of the lesion. These dividing cells are likely the odontogenic cells that elaborate, upon differentiation, into a mineralized matrix 15-28 days after pulp exposure. The restricted location of these mitotic cells within the crown precisely overlaps the area where the dentinal bridge is formed.
Importantly, no labeling of zone III with the CD68 and MHC IIB antibodies could be visualized (Figs. 2F, 2I), indicating that PCNA-positive cells in the root canal are neither monocytes/macrophages nor dendritic cells. This also indicates that there is no recruitment of inflammatory cells in the root at this stage. What we observed in this zone 48 hrs after the pulp exposure was that PCNA-labeled cells were localized in the pulp stroma (Fig. 2C), contrary to the α-SMA-labeled cells lining the blood vessel walls (Fig. 2L). According to some authors, α-SMA-positive cells, presumed to be pericytes, have been considered as odontoblast progenitors (Alliot-Licht et al., 2001). We cannot exclude the possibility that perivascular cells may have odontogenic potential, but analysis of our data provides strong evidence that another subset of cells in the radicular pulp displays stem cell properties. This provides a foundation for determining whether these cells entering mitosis at a distance from the lesion are radicular pulp resident cells or whether they migrate from the apex.
Until now, the location of stem cells induced to leave their quiescent state in response to injury was unknown. The identification of 3 well-delimited zones of proliferation paves the way for better characterization of the identity of pulp progenitors.
Dental Pulp Stem Cells: A Potential Cell Therapy for Pulp Lesions
Mesenchymal stem cells (MSCs) present in the dental pulp of deciduous and adult teeth (Balic et al., 2010; Menicanin et al., 2010) share similarities with MSCs isolated from other tissues, particularly with MSCs of the bone marrow (BMSCs). The dental pulp stem cells (DPSCs) may display specific characteristics due to their neural crest origin (Huang, 2008). They are thought to contribute to reparative dentin formation, but are still poorly characterized. It appears that the term “DPSCs” represents either heterogeneous populations of progenitor cells and/or distinct differentiation stages along the odontogenic lineage of pulp stem cells. As inferred from flow cytometric analysis performed on primary pulp cultures, the “dental-MSC-like population” expresses different panels of surface molecules such as STRO-1, CD 45, CD 90, CD 117, CD 146, and Sca-1 (Miura et al., 2003). Based on cell-surface antigen trials, distinct precursor cell sub-populations have been isolated. In vitro studies have permitted us to assign to these pulp precursor-enriched populations either a multi-lineage differentiation potential or a more restricted commitment toward the odonto-osteogenic lineage (Gronthos et al., 2000; Laino et al., 2006). Besides, when in vitro expanded STRO-1-positive DPSCs mixed with hydroxyapatite particles were implanted subcutaneously into immunocompromised mice, they were shown to form a pulp-dentin-like tissue complex (Gronthos et al., 2000). Thus, the in vivo dentinogenic potential of DPSCs has been clearly established in ectopic sites, but their involvement in reparative dentinogenesis has not been explored.
To characterize the potential of dental pulp progenitors toward odontogenesis and other mesoblastic fates (osteogenic, chondrogenic, adipogenic), we have established clonal pulp precursor cell lines from the first molar of embryonic ED18 mouse (Laoide et al., 1996; Priam et al., 2005). Among the clones obtained, one, named A4, is multipotent and behaves as a genuine mesenchymal stem cell, with the ability to undergo odontogenic, osteogenic, chondrogenic, or adipogenic differentiation in vitro, depending on the nature of the inducers (unpublished observations).
We exploited this pulp-derived cell line to evaluate whether the implantation of multipotent A4 cells may improve tooth repair in a rat molar after pulp exposure. When 105 A4 cells were implanted into the lesion site, inflammation was observed during the first week in both control (pulp exposure only) and implanted teeth. At day 15, the initial formation of a dentinal barrier could be visualized in the implanted teeth but not in the controls, indicating that A4 cell implantation favors the onset of mineralized matrix formation (not shown). At day 28 in the control teeth, a fibrous matrix was formed without closure of the exposure site (Figs. 3A, 3B), and such poorly mineralized matrix still persisted in the controls after 3 months. Conversely, 28 days after A4 cell implantation, the exposure site was completely occluded. A robust osteodentin bridge filling the mesial part of the pulp has been elaborated (Fig. 3C). Moreover, the impact of A4 cells on the reparative process appears to be spatio-temporally controlled. Indeed, mineralization has not expanded to other parts of the pulp, and no abnormal focus of stem cell proliferation can be detected within the teeth. Obviously, the introduction of A4 “stem” cells within the exposed pulp creates proper conditions for the formation of a well-mineralized osteodentin barrier. The marked calciotraumatic line, which separates the native tubular orthodentin from the induced atubular reparative osteodentin, could be visualized and permits the efficacy of the reparation process to be highlighted (Fig. 3D).
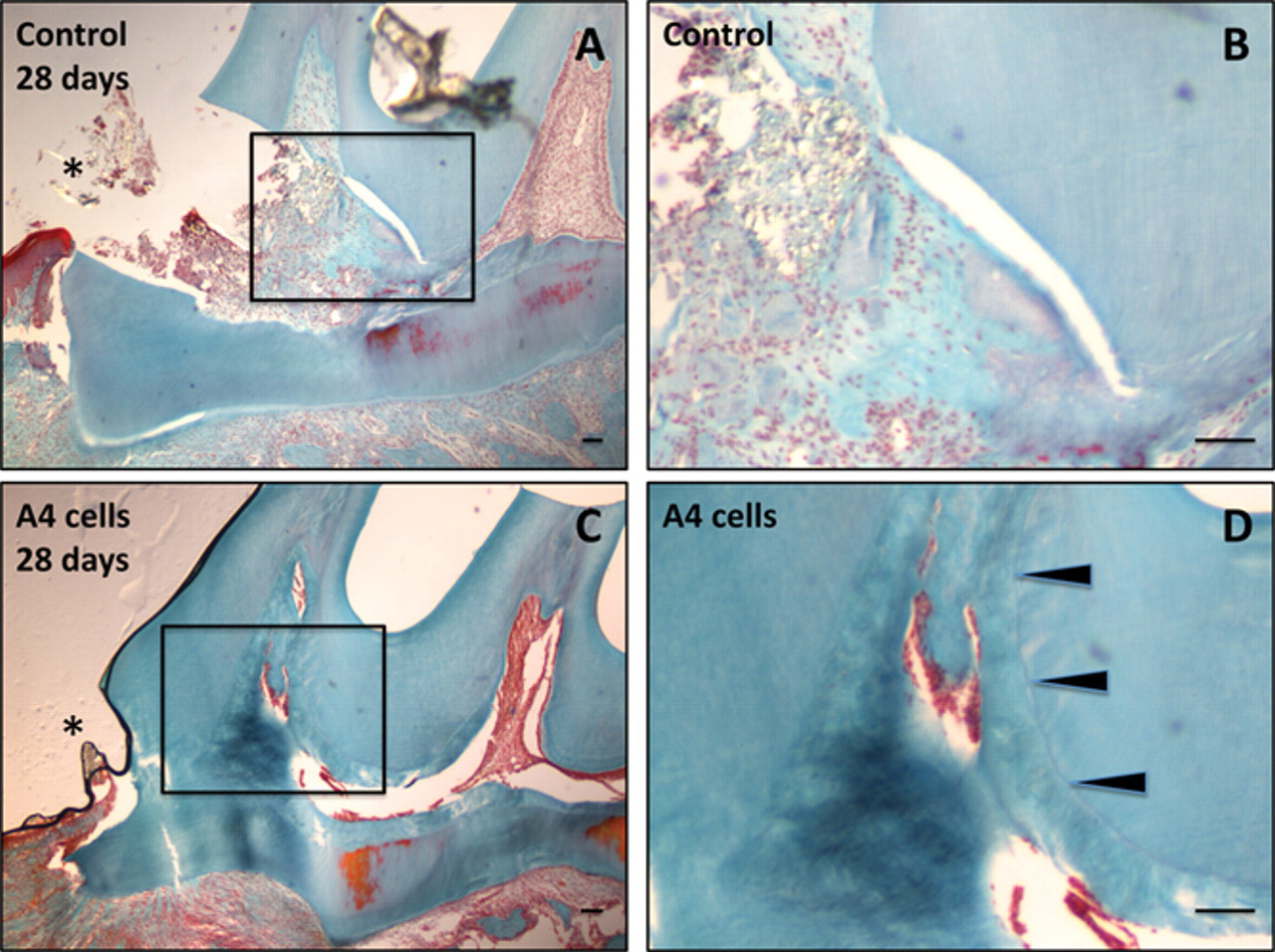
Formation of reparative osteodentin 28 days after implantation of pulp-derived clonal precursor cells in the rat molar. Twenty-eight days after a pulp exposure alone, the mesial pulp chamber is still exposed and filled by a heterogeneous fibrous matrix (
Analysis of these data suggests that the implanted A4 multipotential stem cells convert into odonto-osteogenic cells, are directly involved in reparative mineralized matrix formation, and/or indirectly act by providing paracrine signals for the recruitment and differentiation of resident progenitors.
Conclusion
At present, the feasibility of stem-cell-based bioengineering of teeth is still confronted by methodological and conceptual issues. Better knowledge related to the location and identity of odontogenic precursor cells participating in reparative dentin formation is required. The localization of 3 well-delimited zones of cell proliferation early after pulp exposure constitutes a first step toward the identification of genuine progenitor cell markers and toward the investigation of the signals and the molecular pathways allowing for stem cell recruitment. Finally, our implantation experiments using a pulp-derived precursor cell line provide the prime evidence that, in the absence of any carrier or biomolecule, exogenous stem cells have the capacity to promote efficient tooth repair.
Footnotes
Acknowledgements
This research was partially supported by grants from the Fondation de l’Avenir (2007-2009), by “Institut Français pour la Recherche Odontologique” (IFRO), and by INSERM. Y. Harichane is supported by an IFRO grant, and S. Dimitrova-Nakov is supported by a grant awarded by STEM-Pôle Ile-de-France. We thank C. Willig, EA 2496, Montrouge, for his technical support.