
Abstract
Members of the ephrin/Eph family have recently been shown to be involved in the regulation of bone homeostasis in a murine model. The activation of the EphB4 receptor on osteoblasts by its ligand ephrin-B2 led to stimulation of osteoblastogenesis and therefore to bone formation. The activation of ephrin-A2-EphA2 signaling on osteoblasts inhibited the activation of osteoblast-specific gene expression, leading to bone resorption. Fibroblasts within the periodontal ligament periodontal ligament may be one of the first responders to orthodontic forces. Periodontal ligament fibroblasts (PDLF) are mechanoresponsive. Members of the ephrin/Eph family might link mechanical forces received by PDLF with the regulation of osteoblastogenesis on osteoblasts of the alveolar bone. To study whether ephrin-A2 is modulated upon compression, we subjected human primary PDLF to static compressive forces (30.3 g/cm2). Static compressive forces significantly induced the expression of ephrin-A2, while the expression of ephrin-B2 was significantly down-regulated. Moreover, osteoblasts of the alveolar bone stimulated with ephrin-A2 in vitro significantly suppressed their osteoblastogenic gene expression (RUNX2, ALPL) and decreased signs of osteoblastic differentiation, as demonstrated by a significantly reduced ALP activity. Together, these findings establish a role for this ligand/receptor system linking mechanical forces with the regulation of osteogenesis during orthodontic tooth movement.
Introduction
Fundamental biochemical alterations of the dental pulp, the periodontal ligaments (PDL), the alveolar bone, and the gingiva are required for the process of orthodontic tooth movement (OTM). Of pivotal importance during OTM is the conversion of mechanical forces to biological signaling causing the profound changes allowing for OTM (Meikle, 2007; Wise and King, 2008; Krishnan and Davidovitch, 2009). During OTM, the applied forces immediately lead to strain in the PDL matrix and change the fluid flow in the PDL and bone. As a consequence, one side of the PDL is compressed (“compression sites”), and the opposite side of the PDL is stretched (“tension sites”). Because of the mechanosensitive properties of osteocytes, periodontal ligament fibroblasts (PDLF), and osteoblasts (OB), bone resorption is occurring at compression sites, and bone formation is evident at tension sites, allowing the tooth to be moved through the alveolar bone. How the applied mechanical loads are linked to the biological response is not fully understood.
Ephrin ligands and Eph receptors have been characterized initially for their roles in developmental processes during embryogenesis (Holder et al., 2000). Eph receptor-dependent pathways are critically involved in neural development, cell morphogenesis, tissue patterning, and angiogenesis (Pasquale, 2005). Ephrin-Eph signaling is tightly connected with alterations of the cytoskeleton mediating cell migration and cell adhesion. As exemplified on endothelial cells, mechanical forces (e.g., strain) which alter the interaction of the cells with the ECM and/or influencing the organization of the cytoskeleton have an impact on the function and expression of members of the ephrin/Eph family (Korff et al., 2008; Obi et al., 2009). Members of the ephrin/Eph family have recently been shown to be involved in the regulation of bone homeostasis. Zhao and colleagues reported an NFATc1 (nuclear factor of activated T-cells, cytoplasmic 1)-dependent Ephrin-B2 expression during the RANKL (receptor activator of nuclear factor kappa-B ligand)-induced differentiation of osteoclasts (Zhao et al., 2006). Ephrin-B2-dependent activation of the EphB4 receptor on osteoblasts together with ephrin-B2-dependent reverse signaling on osteoclasts led to a suppression of osteoclast differentiation with a concurrent stimulation of osteoblastogenesis and therefore to bone formation (Mundy and Elefteriou, 2006; Zhao et al., 2006). Irie et al. (2009) demonstrated that the activation of the EphA2 receptor on osteoblasts by its ligand ephrin-A2 expressed on osteoclasts inhibited the activation of osteoblast-specific genes. Simultaneously, the activation of the EphA2 receptor on osteoclasts stimulated osteoclast differentiation and activation which led to bone resorption. Together, both systems might play an important role in the regulation of bone homeostasis.
Eph receptors, like receptor tyrosine kinases (RTK), initiate signal transduction through autophosphorylation after ligand-receptor engagement, referred to as “forward signaling”. However, in contrast to soluble ligands for RTKs, the ephrins display unique features in that they are membrane-bound and capable of receptor-like active signaling (“reverse signaling”), resulting in bidirectional cell-to-cell communication (Kullander and Klein, 2002).
Ephrin-Eph interactions frequently modulate cellular repulsion or attraction. In contrast, the expression and activity of ephrin/Eph family members can be modulated by mechanical forces, indicating a causal relation among mechanical forces, the ECM, the cytoskeleton, and the ephrin-Eph-system. Fibroblasts within the PDL are among the cellular recipients of compressive forces during OTM. Therefore, we hypothesized that PDLF might be able to modulate their ephrin-A2 expression upon the application of compressive forces, and that ephrin-A2 on PDLF via the interaction with the EphA2 receptor on osteoblasts can lead to suppression of osteoblastogenesis at compression sites during OTM.
Materials & Methods
Please see the Appendix for further details.
Primary Cell Cultures
Primary PDL fibroblasts were obtained from juvenile patients (12-20 yrs old) following premolar extraction indicated during orthodontic treatment. Alveolar bone tissue was obtained from patients following osteotomy of third molars. The Ethics Committee (Medical Faculty, University of Heidelberg; Vote S147/2010) approved the harvest of the tissues. Informed consent was obtained from the patients following explanation of the study.
Small tissue fragments were established as explant cultures in DME medium (Invitrogen, Karlsruhe, Germany) supplemented with 10% fetal calf serum (FCS), 2 mM L-glutamine, antibiotics, and antimycotics. Primary PDL cells and primary osteoblasts were used between passages 3 and 9 for the experiments. PDL cells from two different patients (PDL donor, male, age 12 yrs; PDL II donor, female, age 12 yrs) were included in this study. Osteoblasts from the alveolar bone of three different patients were included in this study (OB donor, male, age 17 yrs; OB II donor, male, age 16 yrs; OB III donor, female, age, 12 yrs). A time-course of the mRNA expression of ALPL, IBSP, and BGLAP during the induction of mineralization with “mineralization medium”(DMEM) supplemented with 50 mg/mL ascorbic acid, 10 mM ß-glycerophosphate, and 10−7 M dexamethasone (all from SIGMA-Aldrich, Steinheim, Germany) is given in Appendix Fig. 2.
For stimulation experiments, cells were starved for 24 hrs in culture medium, supplemented with 2% FCS (starvation medium). Ephrin-A2-Fc was pre-clustered with anti-human IgG-Fc (1:10 stoichiometry) in starvation medium for 30 min at room temperature. Anti-human IgG-Fc alone in starvation medium served as a control.
Application of Static Compression
Static compression to PDLF was simulated via centrifugation. Confluent cultures of PDLF were centrifuged in cell culture dishes (60 mm) at 127 g in a swing-out rotor (Beckman GH3.7) for 1, 4, or 6 hrs, respectively. The compressive force applied to the cells was 30.3 g/cm2, simulating clinical orthodontic forces (Davidovitch, 1991). The applied force was calculated according to the following formula: P = (m * r * rpm2 * π2)/A * 9.8 *900), P = pressure [kg/cm2], m = mass (of medium) [g], r = radius [m], and A = surface area of culture dish [cm2]. The application of compressive forces on PDLF has been established previously (Redlich et al., 2004). Centrifugal forces to simulate compression in PDLF have been successfully used by others (Kook et al., 2009).
Quantitative RT-PCR Analysis
Total RNA was isolated from cells by means of the RNeasy-Kit (Qiagen, Hilden, Germany). RNA integrity was monitored by capillary electrophoresis (Experion System, Bio-Rad, Munich, Germany).
Total RNA was subjected to reverse transcription with poly-dT-Primers. Single-stranded cDNA was used for qPCR analyses. Quantitative PCR was performed with TaqMan chemistry or SYBR Green chemistry on an iCycler Instrument (Bio-Rad). To ensure equal amplification efficiencies, we used predesigned TaqMan Gene Expression Assays (Applied Biosystems, Darmstadt, Germany) or RT2; qPCR Primer Assays (Qiagen). See Appendix Methods for assay IDs. The relative gene expression was determined by the delta-delta CT method.
Statistics
Results are presented as mean ± standard deviation (SD). Differences between and among groups were compared by one-way analysis of variance (ANOVA) followed by the appropriate post hoc test. All statistics were performed with SigmaStat software (SPSS Inc., Chicago, IL, USA). Results were considered significant with a p value < 0.05.
Results
Ephrin-A2 and EphA2 Expressions Are Up-regulated and Ephrin-B2 and EphB4 Expressions Are Down-regulated in PDLF Exposed to Compressive Forces
To determine if ephrin-A2 is up-regulated in PDLF, we analyzed the expression of ephrin-A2 by means of quantitative RT-PCR (qRT-PCR) on 2 primary PDLF lines (PDL, PDL II). Compression (30.3 g/cm2) was applied for 1, 4, or 6 hrs. QRT-PCR analysis revealed a significant up-regulation, as compared with static control cells, of ephrin-A2 expression in PDL and PDL II 4 and 6 hrs after the application of compressive forces (Figs. 1A, 1E). Ephrin-B2 was significantly down-regulated after 4 and 6 hrs in compressed PDL and PDL II (Figs. 1C, 1G). The receptors EphA2 and EphB4 were also tested. EphA2 was significantly up-regulated (Figs. 1B, 1F), and EphB4 revealed a trend toward down-regulation in PDL (Fig. 1D) or was significantly down-regulated in PDL II (Fig. 1H).
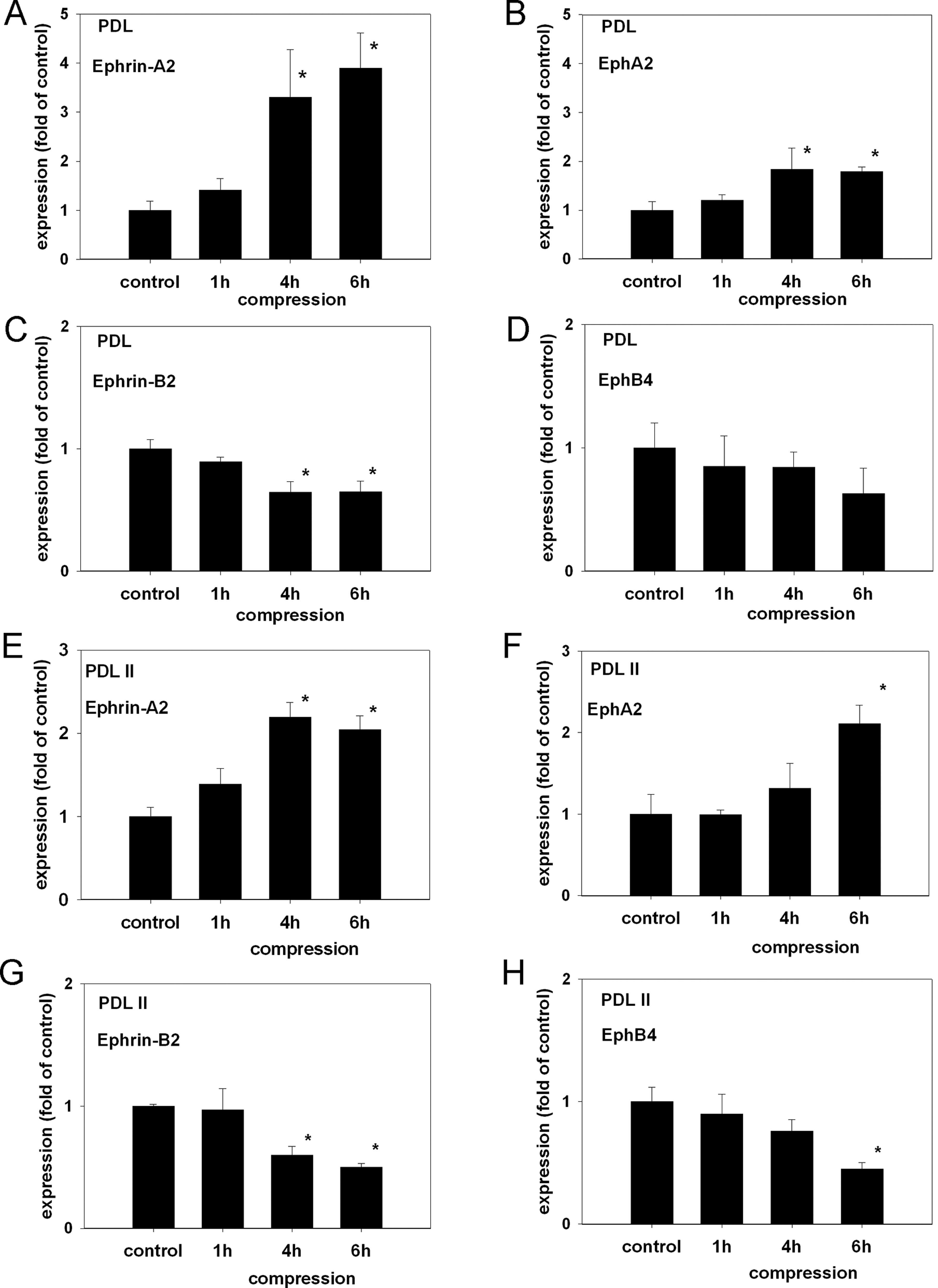
The application of compressive forces induces the expression of ephrin-A2 and EphA2 and reduces the expression of Ephrin-B2 and EphB4 in PDLF. Two PDLF lines (PDL, PDL II) were subjected to compression (30.3 g/cm2) for 1, 4, or 6 hrs. The expressions of ephrin-A2, ephrin-B2, EphA2, and EphB4 were determined by means of qRT-PCR with TaqMan-chemistry.
Ephrin-A2 Attenuates the Transcription of RUNX2 and ALPL in Osteoblasts of the Alveolar Bone
Based on our studies on PDLF, we next sought to investigate the function of ephrin-A2-dependent signaling in osteoblasts of the alveolar bone. First, we analyzed osteoblastogenic gene expression in ephrin-A2-stimulated osteoblasts.
QRT-PCR revealed a significant down-regulation of RUNX2 at a concentration of 1 µg/mL ephrin-A2 (Fig. 2A). The expression of ALPL was likewise significantly attenuated (Fig. 2B). To substantiate the potential impact of ephrin-A2 on RUNX2 and ALPL expression in osteoblasts, we performed QRT-PCR analysis on 2 additional primary osteoblast lines (OB II, OB III) and obtained comparable results (see Appendix Figs. 3A-3D).
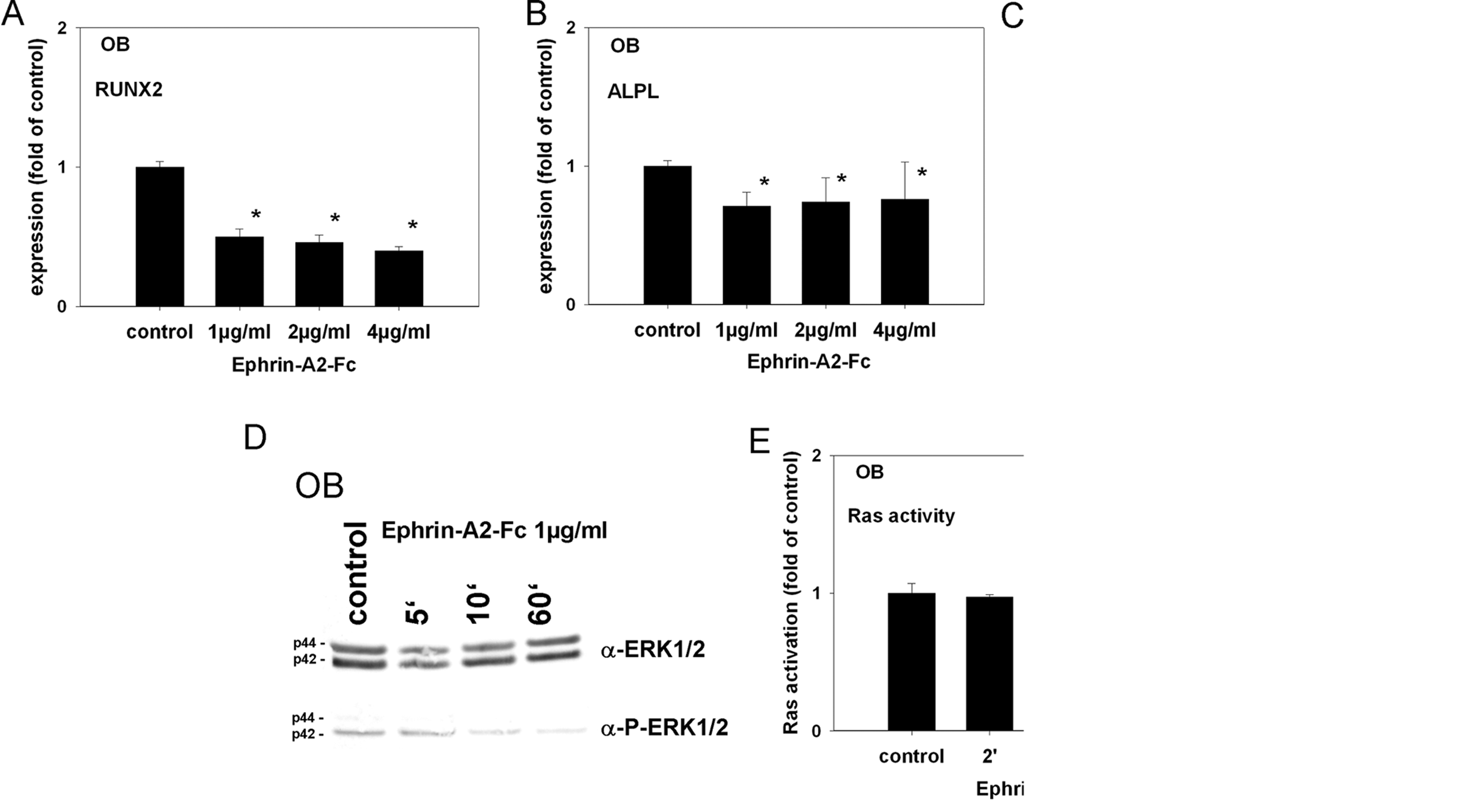
Ephrin-A2 attenuates the transcription of RUNX2 and ALPL in osteoblasts (OB) of the alveolar bone. Hallmarks of osteoblast differentiation are the expression of RUNX2, the pivotal transcription factor in osteoblast differentiation and alkaline phosphatase (ALPL), an early osteoblastic marker and a downstream target of RUNX2. To test for consequences of ephrin-A2-EphA2 signaling in osteoblasts of the alveolar bone, we stimulated osteoblasts with 1, 2, and 4 µg/mL of ephrin-A2-Fc for 48 hrs and analyzed the transcription of RUNX2 and ALPL by means of qRT-PCR using SYBR Green chemistry.
Ephrin-A2-Fc Causes EphA2 Receptor Phosphorylation in Osteoblasts of the Alveolar Bone
To gain more insight into the regulation of the ephrin-A2-dependent EphA2 activation in osteoblasts of the alveolar bone, we analyzed the signaling pathway in more detail. First, we tested whether ephrin-A2-Fc is able to activate the EphA2 receptor on osteoblasts. Tyrosine phosphorylation of the EphA2 receptor was detected after 5 min and sustained throughout the course of the experiment (Fig. 2C).
Ephrin-A2-Fc Suppresses ERK1/2 Activation in Osteoblasts of the Alveolar Bone
To determine whether ephrin-A2-Fc causes the suppression of ERK1/2 activation, we stimulated alveolar bone osteoblasts with 1 µg/mL of ephrin-A2-Fc (preclustered with anti-human IgG-Fc antibodies) for 5 to 60 min. Lysates were probed with antibodies against ERK1/2 and pERK1/2. A decrease in ERK1/2 phosphorylation was observed 10 min after stimulation. Phosphorylation of ERK1/2 remained suppressed during the course of the experiment (Fig. 2D). The analysis of ERK1/2 phosphorylation on OB II and OB II cells confirmed the results obtained for OB (see Appendix Figs. 3E, 3F).
Ephrin-A2-Fc-dependent Suppression of ERK1/2 Is Mediated via a Ras-dependent Pathway
The ERK1/2 (p44 MAPK /p42 MAPK) pathway can be inhibited via the suppression of a Ras/Raf/MEK/MAPK-dependent pathway or through the activation of MAPK phosphatases (MKPs). The regulation of MKPs occurs with slow kinetics (Keyse, 2000). The observed rapid kinetics of ERK1/2 suppression in osteoblasts of the alveolar bone suggested a Ras-dependent mechanism. A Ras activation assay revealed Ras inhibition 5 min after stimulation (Fig. 2E), which was consistent with the onset of ERK1/2 suppression. Ras inhibition was also evident on OB II and OB III (see Appendix Figs. 3G, 3H)
Ephrin-A2-Fc Forfeits ALP Activity on Osteoblasts of the Alveolar Bone
To check for possible functional consequences of ephrin-A2-Fc stimulation-dependent down-regulation of RUNX2 and ALPL, we performed an ALP activity assay on lysates of stimulated osteoblasts. Stimulation with ephrin-A2-Fc led to a significant decrease of ALP activity (Fig. 3A). Comparable results were obtained for OB II and OB III (see Appendix Figs. 4A, 4B).
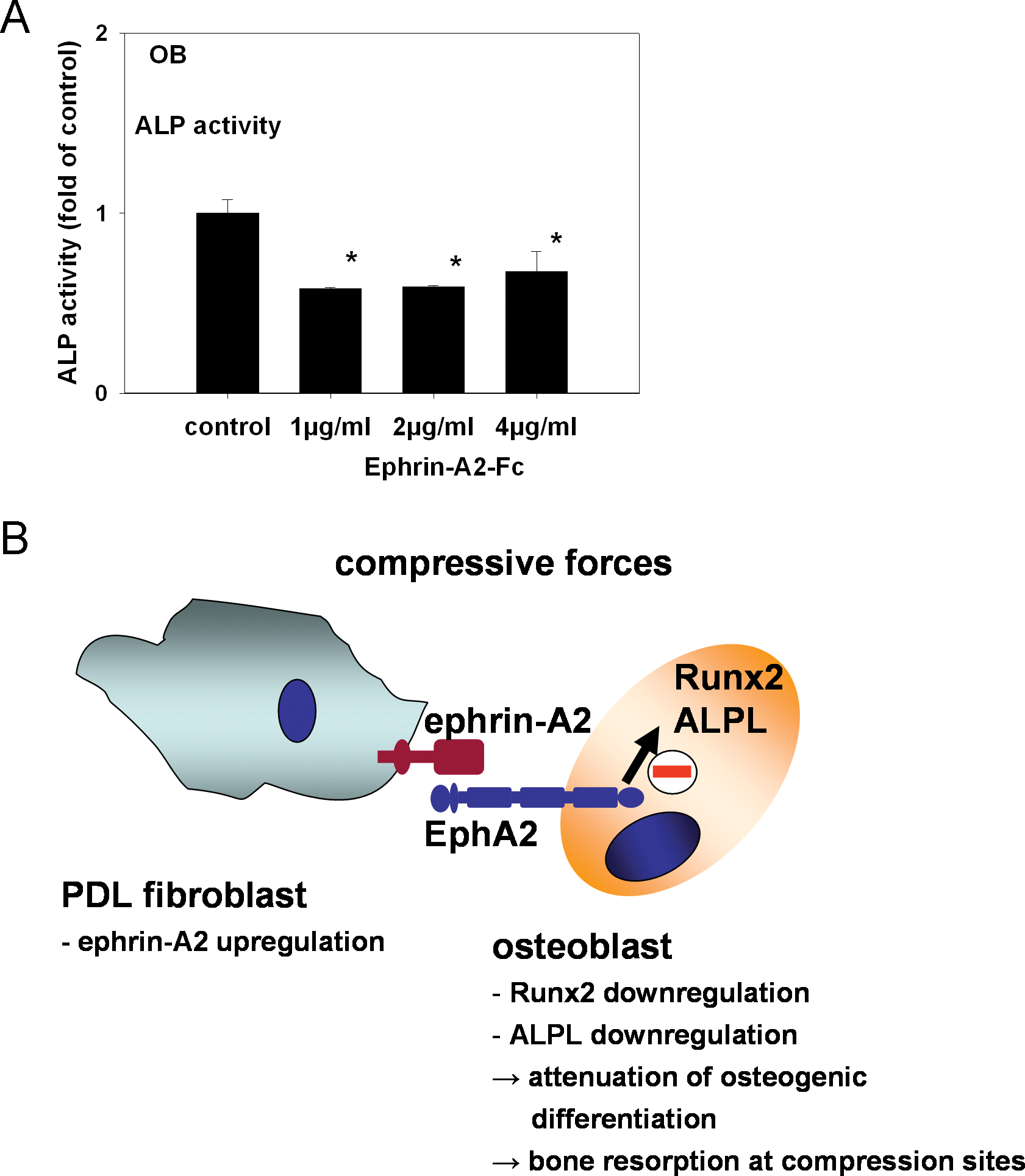
Ephrin-A2-Fc decreased signs of osteoblastic differentiation. To test for functional consequences of the decreased RUNX2 and ALPL transcription, we performed an ALP activity assay on lysates of stimulated osteoblasts. Stimulation with 1 µg/mL ephrin-A2-Fc led to a significant decrease of ALP activity in osteoblasts of the alveolar bone. Stimulation with 2 or 4 µg/mL did not exert an additional effect
Ephrin-A2-Fc Suppresses ALPL and RUNX2 Transcription in PDLF
Different stimuli have been shown to induce osteoblastogenic gene expression in PDLF (Wescott et al., 2007; Heo et al., 2010), and might contribute to osteogenesis during tooth movement. Analysis of the data obtained in this study revealed an anti-osteoblastogenic effect of ephrin-A2-Fc in the osteoblastic lineage. To test the effect of ephrin-A2-Fc stimulation in PDLF, we stimulated PDLF with ephrin-A2-Fc and monitored the transcription of RUNX2 and ALPL by means of qRT-PCR. Expression of both RUNX2 and ALPL at the transcriptional level was significantly down-regulated (Figs. 4A, 4B). PDL II cells were also analyzed for RUNX2 and ALPL expression, and comparable results were obtained (see Appendix Figs. 5A, 5B). These results suggest that ephrin-A2-Fc attenuates osteoblastogenic gene expression synergistically in osteoblasts and PDLF.
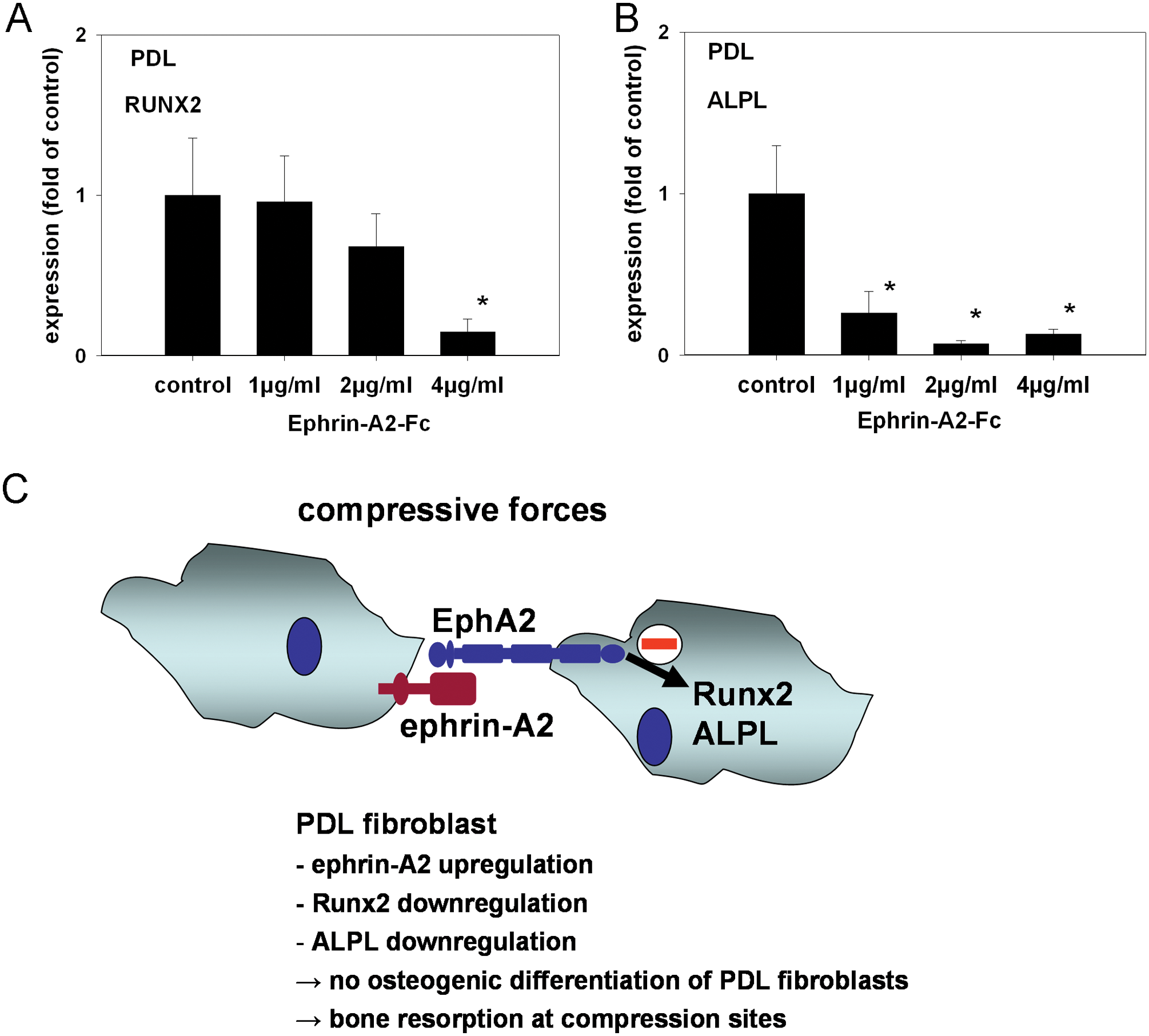
Ephrin-A2-Fc suppresses ALPL and RUNX2 transcription in PDLF. At tension sites during OTM, PDLF have been reported to contribute to osteoblastogenesis. Analysis of the data presented in this study indicates an anti-osteoblastogenic role for ephrin-A2 during OTM at compression sites. To test whether ephrin-A2-Fc stimulation of PDLF might contribute to the observed anti-osteoblastogenic activity, we stimulated PDL with 1, 2, and 4 µg/mL of ephrin-A2-Fc for 48 hrs and determined the expression of RUNX2 and ALPL by means of qRT-PCR using SYBR Green chemistry.
Discussion
During OTM, the application of adequate orthodontic forces allows teeth to be moved through the alveolar bone. Of pivotal importance during OTM is the conversion of mechanical forces to biological signaling, causing the profound changes allowing for OTM.
Ephrins and Ephs can be modulated by mechanical forces in different tissue types, including bone (Xing et al., 2005), and specific members are involved in the regulation of bone homeostasis. Therefore, we hypothesized that the ephrin/Eph family might link mechanical forces with biochemical signaling regulating bone remodeling.
Compression-induced Ephrin-A2 Expression in PDLF
Although it is known that the ephrin ligand ephrin-B2 in endothelial cells, endothelial progenitor cells, and embryonic vessels (le Noble et al., 2004; Korff et al., 2008; Obi et al., 2009) is regulated by cellular strain caused by fluid shear stress, the mechanoresponsive pathway leading to ephrin ligand regulation is not understood. Data on the transcriptional regulation of ephrin-A2 are rare. During the development of the cerebellum in chick embryos, a Ras-MAPK-dependent regulation of ephrin-A2 has been reported: A constitutively active Ras mutant led to MAPK activation and subsequently to an increased expression of ephrin-A2. A functionally inactive Ras mutant had the opposite effect (Vennemann et al., 2008). In breast cancer cells, a tamoxifen-inducible Raf fusion protein caused the activation of the Raf/MAPK pathway, leading to increased EphA2-RNA and protein levels upon induction (Macrae et al., 2005). Analysis of these data suggests an involvement of the Ras/Raf/MAPK-pathway in the regulation of ephrin-A2 and EphA2 expression. The structure of the ephrin-A2 promoter region has not yet been fully characterized. Nevertheless, ephrin-A2 has been identified as a transcriptional target of c-fos (Irie et al., 2009). Fos- and Jun family members together form dimeric transcription factors: AP-1 (activator-protein-1). AP-1 transcription factors are regulated in a mechanoresponsive manner (Hipskind and Bilbe, 1998), depending on the activation of MAPK (Kletsas et al., 2002). PDLF compressed by centrifugal forces showed increased AP-1 activity, which was inhibited by the administration of specific ERK-inhibitors (Kook et al., 2009). Ephrin-A2 seems to be regulated via a mechanoresponsive pathway involving Ras, ERK1/2, and members of the AP-1 family of transcription factors.
We have analyzed compressed PDLF and found a transient activation of the Ras GTPase and an activation of ERK1/2 (see Appendix data), suggesting an involvement of the Ras/ERK1/2-pathway in the compression-dependent regulation of ephrin-A2 in our experimental model. However, the signal transduction pathways linking cellular compression and downstream signaling events leading to this effect remain to be fully elucidated.
Interestingly, the application of compressive forces on PDLF not only led to an increase of ephrin-A2 expression but also attenuated the ephrin-B2-EphB4 pathway, which has been shown to induce osteoblastogenesis, by decreasing the transcription of ephrin-B2 and EphB4 in PDLF. The simultaneous reciprocal regulation of both ephrin-Eph signaling pathways strengthens their possible role in linking mechanical forces applied to PDLF and signal transduction events regulating osteogenesis in osteoblasts of the alveolar bone.
Ephrin-A2 Up-regulation in PDLF Attenuates Osteogenesis in Osteoblasts of the Alveolar Bone
Only recently have ephrin-B2-EphB4 signaling and ephrin-A2-EphA2 signaling been shown to be involved in the regulation of bone homeostasis. While ephrin-B2-dependent stimulation of EphB4 receptors on osteoblasts led to an induction of osteoblastogenesis, activation of the EphA2 receptor on osteoblasts by its ligand ephrin-A2 expressed on osteoclasts inhibited the activation of osteoblast-specific gene expression (Zhao et al., 2006; Irie et al., 2009).
We have identified ephrin-A2 as being up-regulated on the transcriptional level in primary human PDLF upon the application of compressive forces. In their anatomical setting, PDLF are in physical contact with osteoblasts of the alveolar bone, allowing the cell-to-cell interactions required for ephrin-Eph signaling. To test for putative functional consequences of ephrin-A2-dependent EphA2 receptor stimulations, we have used ephrin-A2-Fc chimeras. For appropriate signaling, soluble ephrin ligands require preclustering with anti-Fc antibodies (Egea and Klein, 2007).
Stimulating osteoblasts of the alveolar bone with ephrin-A2-Fc resulted in a decreased expression of RUNX2, the pivotal transcription factor of osteoblastogenic differentiation. ALPL, a target gene of RUNX2, was also down-regulated. Further functional consequences manifested in a reduced ALP activity. These findings are in line with previous observations. Irie et al. (2009) demonstrated that calvarial osteoblasts transfected with a functionally inactive EphA2 receptor variant showed a stronger reaction to ascorbic acid and ß-glycerophosphate with respect to the expression of RUNX2, ALPL, ALP activity, and calcification, indicating that the intact EphA2 pathway inhibits osteoblastogenesis. Because mechanical stresses lead to cellular messages and outcomes through similar mechanoreceptors and signaling effectors in many types of cells, it is likely that common signaling mechanisms are involved in mechanotransduction pathways. In particular, ERK1/2 is the most prominent kinase activated by mechanical stimuli in most cells examined (Liedert et al., 2006). Many studies have shown that Ras GTPases are upstream effectors that induce ERK1/2 phosphorylation in stress-exposed cells (Basdra et al., 1995). Ephrin-A-dependent EphA activation inhibits the Ras/MAPK cascade, identifying EphA receptors as negative regulators of the Ras/MAPK pathway (Miao et al., 2001). Our present findings demonstrate that Ras and ERK1/2 are central mediators of ephrin-A2-dependent EphA2 signaling in osteoblasts of the alveolar bone, leading to a reduced expression of osteoblastogenic markers and forfeiting signs of osteoblastogenic differentiation. At tension sites during orthodontic tooth movement, Wescott and colleagues (2007) have shown that PDLF might contribute to osteoblastogenesis by initiating osteoblastic differentiation themselves. At compression sites, one would expect the opposite. Stimulation of PDL with ephrin-A2-Fc revealed a significant down-regulation of RUNX2 and ALPL, suggesting that the effects of the up-regulation of ephrin-A2 are not limited to the osteoblast lineage but affect PDLF as well. Ephrin-A2 might, therefore, synergistically attenuate osteogenesis at compression sites in the osteoblast lineage and in PDLF.
During OTM, the tooth is moved through the alveolar bone, which requires bone resorption at sites where the applied forces lead to a compression of the PDL and adjacent mineralized and non-mineralized tissues. At the resorption side, soluble factors such as inflammatory cytokines, RANKL, osteoprotegerin, and bone morphogenic proteins (BMPs) are known to regulate osteoclast differentiation (Meikle, 2007; Wise and King, 2008; Krishnan and Davidovitch, 2009). Specific members of the ephrin/Eph family are involved in the regulation of bone homeostasis, and ephrins and Ephs can be mechanically modulated. At compression sites during OTM, ephrins and Ephs might be additional integrators of mechanical forces. We demonstrated that ephrin-A2 expression is up-regulated in compressed PDLF, and that ephrin-A2 has an anti-osteoblastogenic effect on osteoblasts of the alveolar bone. In a physiological setting, ephrin-A2 modulation in PDLF might support bone resorption by attenuating osteoblastogenesis in neighboring osteoblasts.
In conclusion, the results of our study support an important role for ephrin-A2-dependent EphA2 signaling between PDLF and osteoblasts at compression sites of bone remodeling. The results of our study suggest a novel concept in the regulation of bone remodeling during orthodontic tooth movement.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.