
Abstract
Some management strategies for chronic orofacial pain are influenced by models (e.g., Vicious Cycle Theory, Pain Adaptation Model) proposing either excitation or inhibition within a painful muscle. The aim of this study was to determine if experimental painful stimulation of the masseter muscle resulted in only increases or only decreases in masseter activity. Recordings of single-motor-unit (SMU, basic functional unit of muscle) activity were made from the right masseters of 10 asymptomatic participants during biting trials at the same force level and direction under infusion into the masseter of isotonic saline (no-pain condition), and in another block of biting trials on the same day, with 5% hypertonic saline (pain condition). Of the 36 SMUs studied, 2 SMUs exhibited a significant (p < 0.05) increase, 5 a significant decrease, and 14 no significant change in firing rate during pain. Five units were present only during the no-pain block and 10 units during the pain block only. The findings suggest that, rather than only excitation or only inhibition within a painful muscle, a re-organization of activity occurs, with increases and decreases occurring within the painful muscle. This suggests the need to re-assess management strategies based on models that propose uniform effects of pain on motor activity.
Introduction
Our understanding of the effect of pain on muscle activity has been dominated for many years by the proposals of the Vicious Cycle Theory (Travell et al., 1942; Johansson and Sojka, 1991) and the Pain Adaptation Model (Lund et al., 1991). These models propose either excitation or inhibition, but not both, within a painful muscle, and they have influenced some management strategies for chronic pain, e.g., attempts to break a so-called vicious cycle between pain and muscle activity. However, these models do not readily explain all effects of pain on electromyographic (EMG) activity. For example, while some previous studies have demonstrated that single-motor-unit (SMU, the basic functional unit of muscle) discharge rate or multi-unit EMG activity is reduced during noxious muscle stimulation, these models do not explain why reductions in EMG activity can occur while maintaining the same force level during the painful stimulation as in the pain-free condition (Graven-Nielsen et al., 1997; Sohn et al., 2000, 2004; Wang et al., 2000; Farina et al., 2004, 2005; for review, see Murray and Peck, 2007). Explanations for this include changes in activity patterns in other agonist and antagonist muscles, changes in the mechanical properties of SMUs (Sohn et al., 2000, 2004; Farina et al., 2004, 2005, 2008), or recruitment of additional SMUs within the painful muscle (Tucker et al., 2009; Tucker and Hodges, 2009). While there is not good evidence for the former 2 explanations (Farina et al., 2008; Hodges et al., 2008), in recent pain studies in limb muscles, there is good evidence for recruitment of additional motor units in the presence of a reduced discharge rate or cessation of firing of motor units that were active in the pain-free condition (Tucker et al., 2009; Tucker and Hodges, 2009; Hodges and Tucker, 2011).
However, the recruitment of additional motor units to assist in force maintenance, as shown in these limb muscle studies, has not been demonstrated in the jaw motor system. It is important to determine whether re-organization is the response of the jaw motor system in pain, because some management strategies for chronic orofacial pain are based on earlier simplistic models proposing uniform effects of pain on muscle activity (for review, see Murray and Peck, 2007), as well as to provide baseline information assisting future studies guiding prosthodontic management. The aim of this study, therefore, was to determine if experimental painful stimulation of the masseter muscle resulted in only increases or only decreases in activity within the masseter muscle during the generation of the same direction and level of jaw-closing force. The hypotheses were that, during the generation of the same bite force level under pain-free and pain conditions, painful stimulation of the masseter will result in (a) a significant change (i.e., increases and/or decreases) in the discharge rates of some masseter motor units, and (b) the recruitment of additional single motor units to assist in the maintenance of bite force.
Methods
Ten participants without signs and symptoms of temporomandibular disorders or any other health problem were recruited (five men, five women; ages 36.9 ± 10.6 yrs). All participants gave informed consent, and procedures were approved by the Western Sydney Local Health District Human Ethics Committee and the Human Ethics Committee of the University of Sydney. Many of the procedures have been previously described (Phanachet et al., 2001; Sae-Lee et al., 2006, 2008a).
Jaw-muscle EMG Recording
Bipolar fine-wire electrodes were placed within the right masseter muscle via a 25-mm-long needle (Sae-Lee et al., 2006), approximately in the middle of the masseter and to a depth of ~20 mm or until bone was contacted. Then the needle was withdrawn. No participant was in pain several seconds after electrode placement. The EMG activity was amplified (Model DBA-S, World Precision Instruments Ltd, Hertfordshire, UK; 5,000 – 10,000x), filtered (bandwidth 100 Hz–10 kHz), digitized (sampling rate: 20,000 samples/sec; Model micro1401, Cambridge Electronic Design [CED], England), and displayed for the experimenter only.
Bite-force Recording and Jaw Tasks
Bite force was measured with a one-dimensional force transducer (Fig. 1, left panel). The lower plate was parallel to the occlusal plane, and the jaw would slide horizontally with non-orthogonal forces. We confirmed stability of the contact position between the ball and the lower plate by marking the plate with articulating paper. The transducer force signal was digitized (sampling rate 1,000 samples/sec, bandwidth 0–500 Hz) and displayed for participant feedback. Participants initially increased bite force until a target force level where a single SMU or a small number of SMUs could be discriminated by the experimenter. In each trial, participants were instructed to match bite force as closely as possible to the target force level identified for that participant and to do this for all biting force trials under both blocks. At 2 to 5 sec after the beginning of each biting task trial, participants increased bite force rapidly to the target force level for ~ 5 sec.
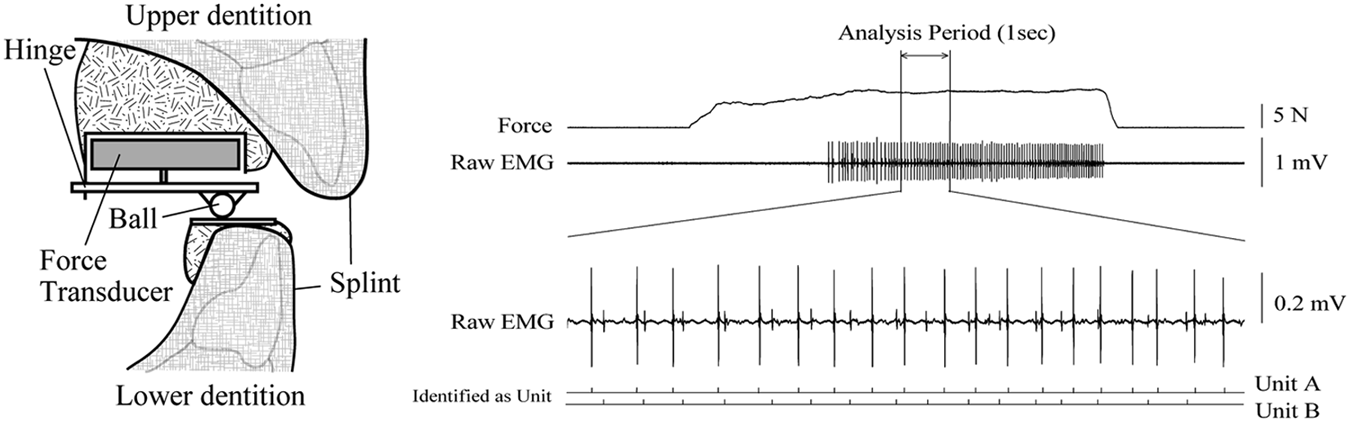
Induction and Assessment of Jaw-muscle Pain
Experimental pain was induced by 5% hypertonic saline (Phebra Pty. Ltd., Lane Cove, NSW, Australia) infused into the deep central region of the right masseter muscle. In a separate block of biting force trials on the same day, isotonic saline was infused through the same indwelling catheter as a control. A disposable 24-gauge (19-mm length) needle integrated IV catheter was inserted about 2 cm posterior to the SMU electrode. An initial bolus infusion of 0.2 mL 5% hypertonic saline was infused over 20 sec, followed by continuous infusion maintained by an infusion pump (IVAC Model P2000, IVAC Medical Systems Inc, San Diego, California, USA) at an infusion rate of 2 to 9 mL/hr. Pain intensity was recorded on a 100-mm visual analog scale (VAS; 0 mm, no pain at all; 100 mm, the worst imaginable pain) prior to catheter insertion, at every 30 sec after commencement of infusion until pain ratings were ~50 mm, and then after each trial. Manual changes in infusion rate maintained pain intensity ~50 mm.
A block of 3 to 6 trials of the biting task was performed in a repeated-measures design: test 1 (hypertonic saline infusion, “pain block”), 10-minute rest, and test 2 (isotonic saline infusion, “no or minimal pain block”, hereafter termed the “no-pain block”). The order of solutions injected was alternated between and among participants. Before the hypertonic or isotonic saline blocks, all participants completed a baseline block of 3 to 6 trials of the biting task to accustom the participants to the task. At the end of each infusion, each participant was asked to outline, on a lateral profile drawing of the head, the maximum extent of pain perception.
Data Analysis
From each trial, 1 second’s worth of data with the lowest variability (i.e., where the SD was the lowest) from the no-pain and pain blocks in force was selected (Tucker and Hodges, 2009; see Table 1). Fig. 1 (right) shows representative data for 2 discriminated SMUs (units A, B) during a no-pain block. We used the statistical methods of Bland and Altman for assessing agreement of force levels between pain and no-pain blocks (Bland and Altman, 1986). A gross measure of muscle activity was provided by the number of crossings of all SMUs by an arbitrary level above baseline noise that was a constant within a participant (Tucker and Hodges, 2009) (Origin Pro 8.0 software, Light Stone, Tokyo, Japan). These crossings were compared between blocks (paired t tests, two-tailed). SMUs were identified based on shape (Spike2 software, CED, UK), and abnormally short or long inter-spike intervals were omitted. Instantaneous firing frequencies of each SMU were compared between the pain and no-pain blocks (independent sample t tests, two-tailed). For all tests, significance was p < 0.05 (IBM SPSS Statistics 20.0, IBM Corporation, Tokyo, Japan).
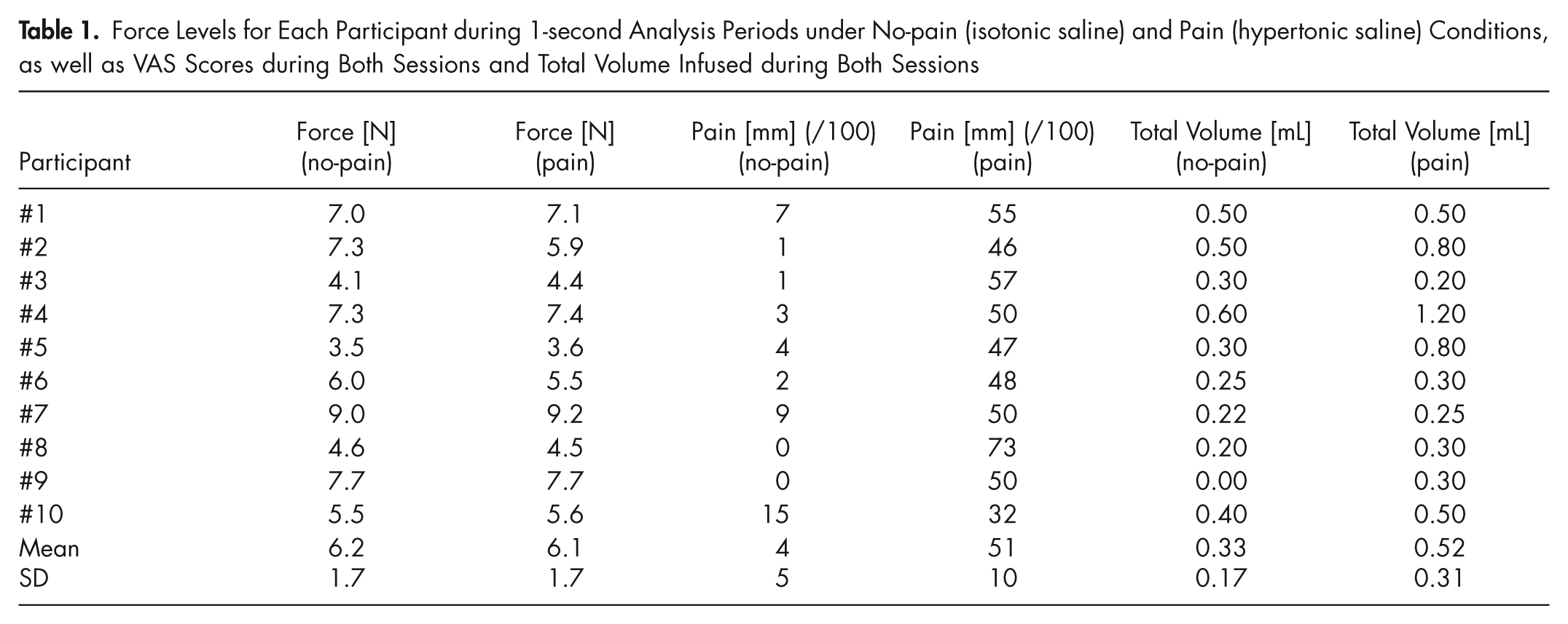
Force Levels for Each Participant during 1-second Analysis Periods under No-pain (isotonic saline) and Pain (hypertonic saline) Conditions, as well as VAS Scores during Both Sessions and Total Volume Infused during Both Sessions
Results
Pain Scores
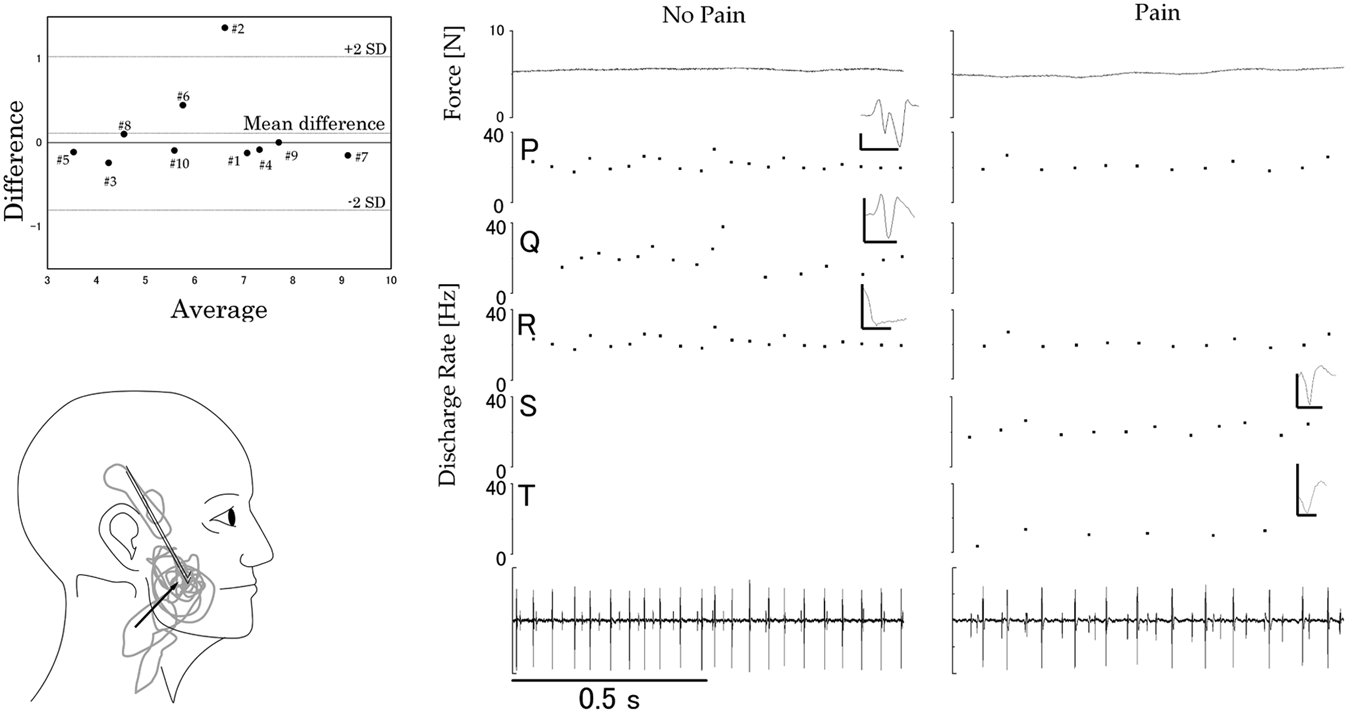
The mean (SD) pain ratings reported during all trials following the injection of hypertonic saline [pain block; 51 (10) mm] were significantly different (p < 0.001) from the scores reported during isotonic saline infusion [no-pain block; 4 (5) mm)] (Table 1). Hypertonic saline pain maps were generally centered at the injection site, although two individuals exhibited referral sites to the temple region or the angle of the mandible/neck region (Fig. 2, lower left panel).
Number of Appearances and Discharge Rates of All Identified Single Motor Units
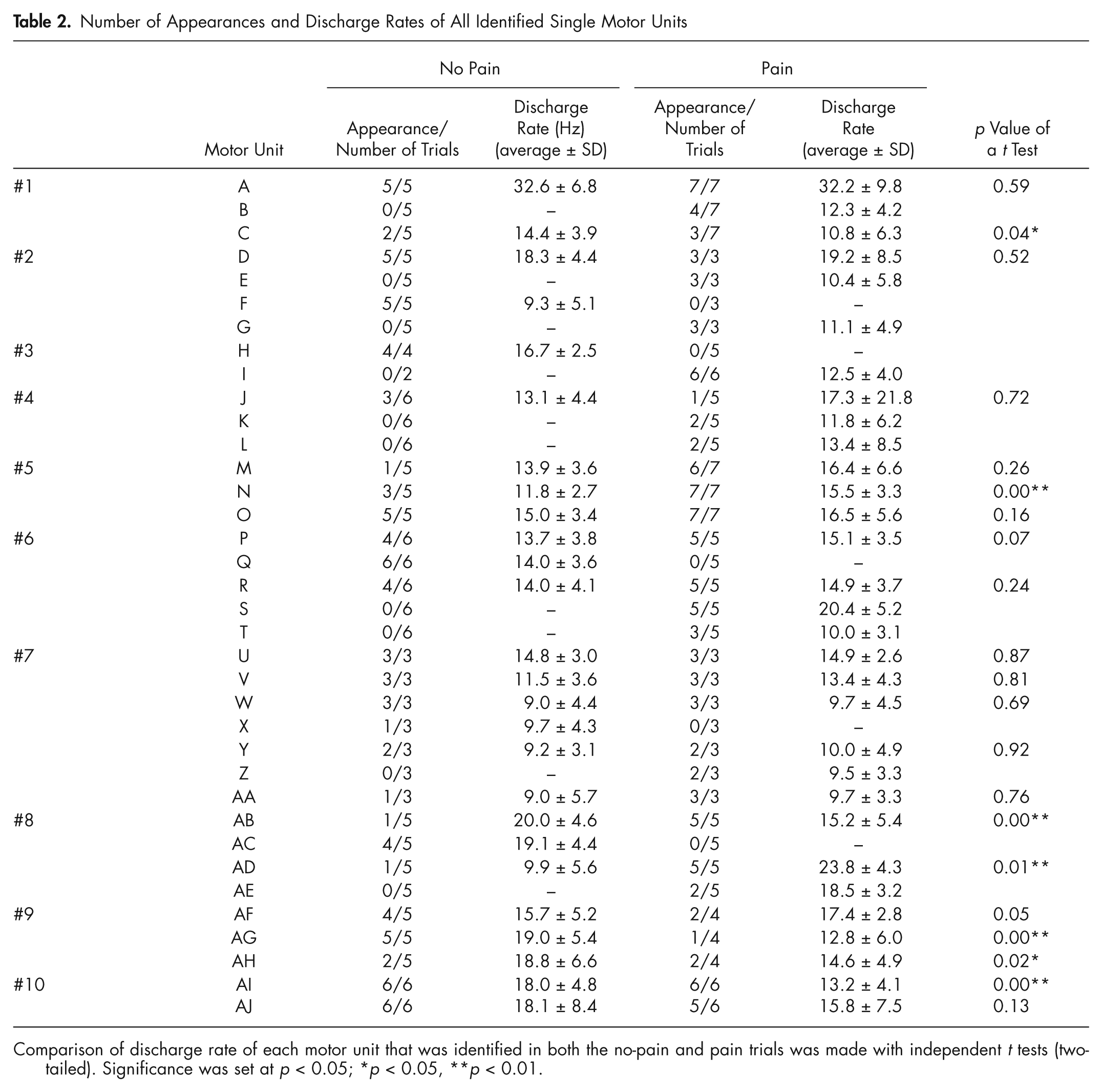
Comparison of discharge rate of each motor unit that was identified in both the no-pain and pain trials was made with independent t tests (two-tailed). Significance was set at p < 0.05; *p < 0.05, **p < 0.01.
Force Levels
The mean (SD) force during the most stable 1-second analysis period during the contraction without pain [6.2 (1.7) N] was not significantly different (p = 0.51) from that during the 1-second analysis period during pain [6.1 (1.7) N; Table 1]. Bland and Altman diagrams (difference between the pain and no-pain block, divided by the average of these 2) (Fig. 2, upper left) confirm that the mean difference between the 2 blocks was not significantly different from zero (see Fig. 2 legend).
Single Motor Unit Recordings
In total, 36 SMUs were discriminated (Table 2). Of these, the same 21 were present during one or more trials of both the no-pain block and the pain block. Of these 21, there was no significant difference (p = 0.6) in mean firing rates between the no-pain [14.7 (5.1) Hz] and the pain [15.3 (5.9) Hz] blocks. Of these 21 SMUs, in four participants 6 SMUs were present in every force trial of both pain and no-pain blocks, and there was no significant change in firing rates of these 6 units between blocks. However, of the 21 SMUs present during the pain and no-pain blocks, there was a significant increase in firing rates for 2 units (Table 2, units N and AD), a significant decrease for 5 units (Table 2, units C, AB, AG, AH, AI, p < 0.05), and no significant change for 14 units.
Of the remaining 15 SMUs of the total of 36 units, 5 were present during the no-pain block only (Table 2, units F, H, Q, X, and AC; e.g., Fig. 2, right panel). Ten units were present in the pain block and not in the no-pain block (Table 2, units B, E, G, I, K, L, S, T, Z, and AE; e.g., Fig. 2, right). In five of the participants (Table 2, participants #2, #3, #6, #7, and #8), the de-recruitment of one SMU in the pain block coincided with the recruitment of at least one SMU that was not present in the no-pain block. There was no significant difference (p = 0.47) between the no-pain [mean (SD): 280 (369)] and pain [309 (428)] blocks in the number of threshold crossings.
Discussion
Reorganization of Activity Within a Painful Muscle
The first hypothesis—that painful masseter stimulation will result in a significant change in the discharge rates of some masseter motor units—was accepted. The second hypothesis was also accepted, that is, during painful stimulation of the masseter muscle, additional single motor units will be recruited. Thus, 10 SMUs were active in the pain block only and not the no-pain block, and, in five participants, the de-recruitment of at least one SMU coincided with the recruitment of at least one SMU that was not present in the no-pain block. The recruitment of additional motor units during painful stimulation provides a mechanism whereby the jaw-closing force level can be maintained in the presence of a decrease in the firing rates of some motor units.
These new findings provide general support for the view emerging from limb and trunk muscle studies, that pain results in neither inhibition only nor excitation only but rather a re-organization of activity within the painful muscle, that is, a slowing and/or de-recruitment of one population of motor units and a recruitment of a new population of units (Tucker et al., 2009). Analysis of the new data points to the need for re-assessment of management strategies for patients with jaw muscle pain conditions based on models (e.g., Vicious Cycle Theory, Pain Adaptation Model; Travell et al., 1942; Johansson and Sojka, 1991; Lund et al., 1991) proposing generalized reductions or increases in EMG activity within muscles subject to painful stimulation. Rather than supporting these earlier models, the findings lend support to more recent models proposing a re-organization of the recruitment strategy adopted by the brain in the control of motor units in the presence of pain (Murray and Peck, 2007; Hodges and Tucker, 2011).
Deviations from previous studies showing only reductions in EMG activity during noxious stimulation may reflect methodological differences, such as the averaging effect from surface EMG recordings where reductions in SMU firing might mask increases in activity or recruitment. In at least one previous study (Sohn et al., 2000), the pre-set force levels may have precluded the investigators’ ability to discriminate all SMUs in the recording, whereas most SMUs were able to be discriminated in the present study.
Methodology and Limitations
Although only a uniaxial force transducer was used, changes in force direction during closing other than perpendicular to the occlusal plane would have resulted in the upper ball slipping off the lower plate when applied lateral forces exceeded frictional forces. No such positional changes were detected in the present study. Further, in four participants (participants #1, 2, 5, 7) showing significant changes in firing rates and/or recruitment or de-recruitment of SMUs, 6 SMUs were present in every force trial of both pain and no-pain blocks, and there were no significant changes in firing rates of these units between blocks. Future studies could utilize a 3-dimensional force transducer to determine if pain induces subtle changes in force direction, as has recently been demonstrated in the leg muscle system (Tucker and Hodges, 2010). There are several reasons ruling out possible direct effects of hypertonic saline or the injected volume of the solution on motor units. First, hypertonic saline does not have an effect on muscle fiber electrophysiological properties (Farina et al., 2004, 2005). Second, there was evidence for stability of firing of some motor units where changes were noted in other motor units recorded with the same electrode. Third, isotonic saline was used as a control. Finally, our findings are entirely consistent with recent findings in the spinal literature (Tucker et al., 2009). Fatigue is also unlikely a factor, given the short duration and number of the holding trials and the absence of significant changes in firing rates of 21 of the recorded SMUs between control and pain conditions.
Footnotes
Acknowledgements
The first author is now at the Department of Removable Partial Prosthodontics, Tokyo Medical and Dental University. We acknowledge assistance from Paul Gezelle Meerburg and Willemien Molenaar, visiting students from the Academic Centre for Dentistry Amsterdam (ACTA), and Mr. Kamal Wanigaratne.
This research was supported by the NHMRC of Australia (No. 512309) and the Australian Dental Research Foundation, Inc.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.