
Abstract
We investigated the roles of osteocytes in osteoclastic bone resorption during orthodontic tooth movement using the transgenic mice in which osteocytes can be specifically ablated. Because these transgenic mice express the receptor for diphtheria toxin on the cell surfaces of osteocytes, the injection of diphtheria toxin can ablate their osteocytes in vivo. Injection of diphtheria toxin into the transgenic mice significantly increased the number of ablated osteocytes in alveolar bone compared with that in wild-type mice with or without diphtheria toxin injection. Increased numbers of ablated osteocytes were observed from day 4 to day 12 after the injection in alveolar bones as well as in cortical bone of the tibiae. We applied the orthodontic force 4 days after the injection of diphtheria toxin, and the distance of tooth movement on day 12 was significantly smaller in transgenic mice than that in control mice. The numbers of osteoclasts and the quantity of eroded bone surface at the compression site were significantly reduced in the transgenic mice injected with diphtheria toxin than in control mice. These results provide in vivo demonstration of osteocyte involvement in osteoclastic bone resorption during orthodontic tooth movement.
Introduction
Orthodontic tooth movement is achieved by a repeated process of alveolar bone remodeling by bone resorption and bone formation. Many studies exploring the mechanism underlying orthodontic tooth movement have focused on the roles of osteoclasts and osteoblasts during tooth movement (Waldo and Rothblatt, 1954; Storey, 1973; Proff and Römer, 2009).
Osteocytes are considered to play a central role in mechanical sensing in bone (Noble, 2008; Bonewald, 2010). Osteocytes regulate the differentiation of osteoblasts and osteoclasts during bone remodeling induced by mechanical loading (Yamashiro et al., 2001; Fujihara et al., 2006; Robling et al., 2008). Recently, 2 research groups revealed that the receptor activator of NF-κB ligand (RANKL) synthesized by osteocytes plays a crucial role in osteoclastogenesis (Nakashima et al., 2011; Xiong et al., 2011). Although increasing evidence suggests that osteocytes play a crucial role in bone remodeling induced by mechanical loading, there have been no reports that directly demonstrated the impact of osteocytes on orthodontic tooth movement.
Tatsumi et al. established the osteocyte-ablated mice using the diphtheria toxin receptor (DTR)-mediated cell knockout (TRECK) system (Tatsumi et al., 2007). They expressed TDR in osteocytes using the 9.6-kb Dmp1 promoter (DTR-Tg mouse), which is specifically activated in osteocytes and odontoblasts (Lu et al., 2007). After injection with diphtheria toxin (DT) into these mice, empty lacunae increased markedly in femoral cortical bones. Interestingly, these osteocyte-ablated mice were resistant to unloading-induced bone loss, indicating that osteocytes play a crucial role in bone remodeling induced by mechanical loading (Tatsumi et al., 2007).
In the present study, we showed that DT-injection in DTR-Tg mice successfully ablated osteocytes in alveolar bones and significantly inhibited tooth movement. The present study provides the first evidence showing that osteocytes are involved in orthodontic tooth movement.
Materials & Methods
Experimental Animals
Wild-type mice (C57BL6/J) were purchased from the Jackson Laboratory (Bar Harbor, ME, USA), and C57BL/6Cr-Tg (Dmp1-HBEGF) transgenic (DTR-Tg) mice were obtained from RIKEN BRC (The National Bio-Resource, Tsukuba, Japan). DTR-Tg mice express human HB-EGF as the DTR on the cell surfaces of osteocytes under the control of the Dmp1 promoter (Tatsumi et al., 2007). Eight-week-old male mice were used in all experiments. In total, 96 mice (32 wild-type mice and 64 DTR-Tg mice) were used. We injected DT (Sigma-Aldrich Japan K.K., Tokyo, Japan) once intraperitoneally into mice 4 days before the application of orthodontic force. Injection of PBS alone was used as a vehicle control. All experimental procedures were approved by the Experimental Animal Committee of Tokyo Medical and Dental University (No. 0090252), and the experiments adhered to ARRIVE guidelines.
Tooth Movement
Orthodontic tooth movement with the 10gf Ni-Ti closed-coil spring, which works continuously by the force of 10gf between about 0.1 mm and 5.0 mm, was achieved as described previously (Yoshimatsu et al., 2006, 2012) (Fig. 1A). In brief, we inserted this appliance between the upper bilateral incisors and the left first molar of mice. The appliance was fixed, with a stainless steel wire (Φ0.09 mm), to a hole drilled through each of the 2 incisors at the alveolar bone level. The posterior end of this appliance was tied to the first molar with the wire. Although easier and more stable methods are available, these procedures prevented the removal of the appliance during tooth movement experiments, and there were no cases of detachment of the orthodontic appliance during the experiments. The right maxillary molars were used as the control in each mouse.
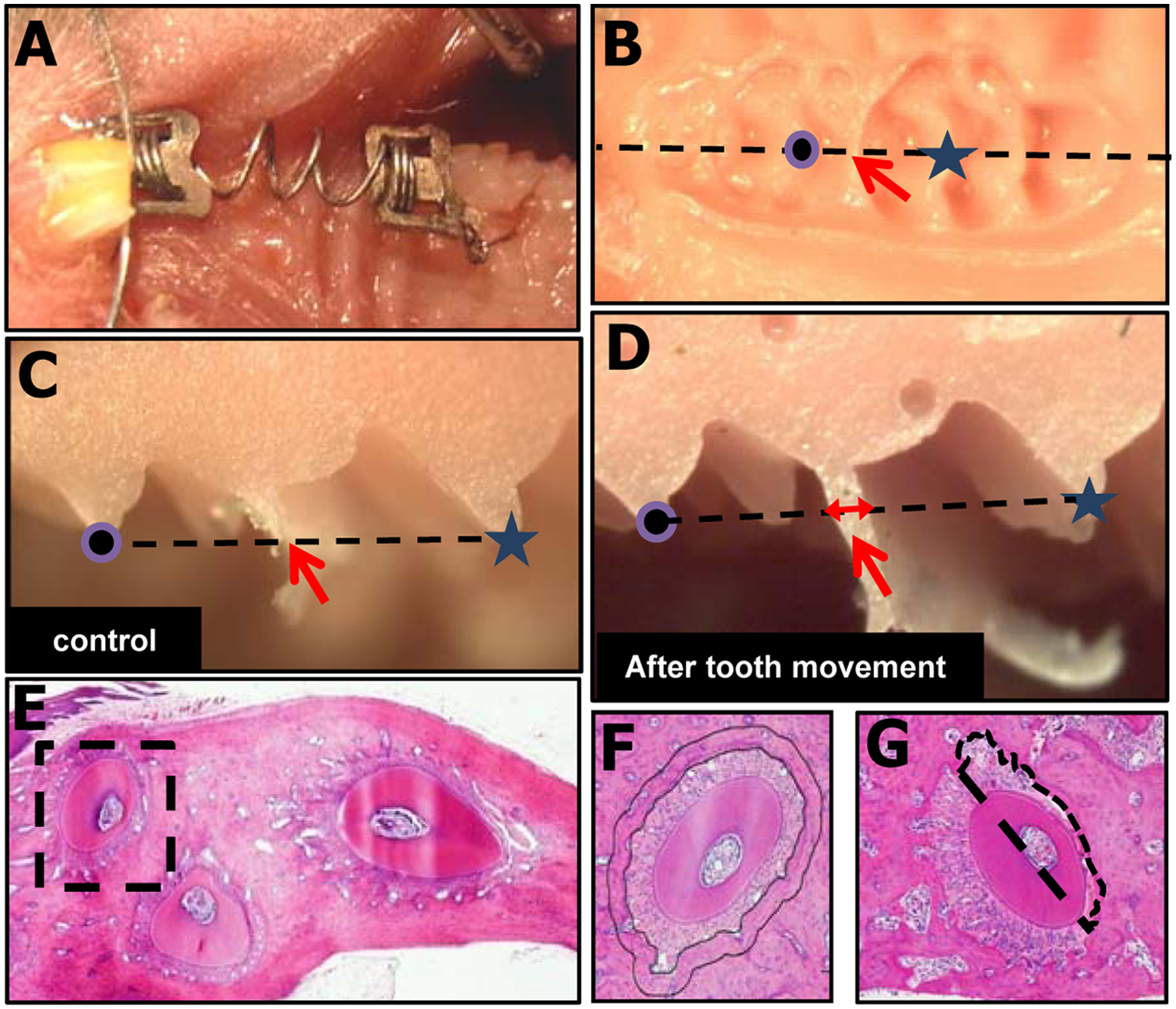
An orthodontic appliance and measurement of tooth movement.
Measurement of Tooth Movement
The distance of tooth movement was measured on days 4, 8, and 10 after the application of the tooth movement appliance in each experimental group (n = 4). After dissection of the maxillary region, impressions of the teeth and maxillae were obtained with the use of individual trays containing hydrophilic vinyl polysiloxane dental impression material (Exafine, GC Co., Tokyo, Japan) (Fig. 1B). The samples were fixed in 4% paraformaldehyde after the impressions were taken. The impression was cut on the line connecting the middle of the distal fissure in the first molar and the middle of the mesial fissure in the second molar (Fig. 1B). The distance of tooth movement was evaluated by the thickness of the dental impression material between the first and second molars in a cross-sectional surface of the impression observed under a digital microscope (Figs. 1C, 1D). MicroCT images were taken by a micro-CT system (SMX-100CT; Shimadzu, Japan) in DTR-Tg-mice with or without DT injection. The images consisted of 1,200 slices with a voxel size of 8 μm in all 3 axes.
Tissue Preparation
The fixed teeth and maxillae were decalcified in 20% ethylene-diaminetetraacetic acid at 4°C. After dehydration, the samples were embedded in paraffin, and 50 serial transverse sections of the first molar region were cut from the first molar bifurcation to 300 µm toward the root apex (Kobayashi et al., 2000). The distal buccal root of the upper first molar was used for the following histomorphometric analysis (Fig. 1E). We evaluated 10 sections at 30-µm intervals in each animal, and the histomorphometric values are expressed as the average of these 5 sections.
Measurement of Ablated-osteocyte and Osteoclast Numbers
To estimate the number of ablated osteocytes, we counted empty lacunae by using ImageJ software. The empty lacunae were counted in the alveolar bone around the distal buccal root within a 50-µm width (Fig. 1F), and the number of empty lacunae was expressed by a percentage of the total lacunae number at the measured area. The osteoclast surface (N.Oc/BS; osteoclast number per bone surface) and eroded surface (ES/BS; eroded bone surface per bone surface) were counted at the compression site of the mesial half area of alveolar bone (Fig. 1G) by identification of tartrate-resistant acid phosphatase (TRAP)-positive cells that contained more than 3 nuclei, as described previously (Ishikuro et al., 2008; Kayamori et al., 2010). These measurements were conducted on days 8, 12, and 16 after injection with PBS or DT (corresponding to days 4, 8, and 12 after tooth movement) (n = 4 in each experimental group). The histomorphometric parameters were measured with ImageJ software. Nomenclature and units used in the histomorphometric analyses are those recommended by the Nomenclature Committee of the American Society for Bone and Mineral Research (Dempster et al., 2013).
Staining for Actin Filaments with Fluorescein-conjugated Phalloidin
To visualize the osteocyte lacunocanalicular system, we stained the specimens for cytoskeletal actin by using fluorescein-conjugated phalloidin as described previously (Cao et al., 2011). In brief, decalcified samples were embedded in OCT compound, and the cryosections (10-20 μm thick) were incubated with phalloidin-AlexaFluor 488 (Invitrogen, Carlsbad, CA, USA) and BOBO-3 (Invitrogen) at room temperature for 2 hrs.
Statistical Analyses
We used analysis of variance with the F-test, followed by the t test. Any p values less than 0.05 were considered significant. The data are presented as mean ± standard deviation values of independent replicates.
Results
Ablation of Osteocytes in Alveolar Bone
We investigated the effects of DT injection on the ablation of osteocytes at alveolar bone in 4 experimental groups: wild-type mice with PBS-injection (WT;PBS), wild-type mice with DT-injection (WT;DT), DTR-Tg mice with PBS-injection (DTR-Tg;PBS), and DTR-Tg mice with DT-injection (DTR-Tg;DT). Before conducting orthodontic tooth movement, we tested the effective doses of DT injections to induce osteocyte ablation at alveolar bone in DTR-Tg mice. The injection of 50 μg/kg DT significantly increased the number of empty lacunae after 4 days, but 25 μg/kg DT injection or PBS injection did not (Fig. 2A). Thus, we applied a dose of 50 μg/kg for DT injection in the following experiments.
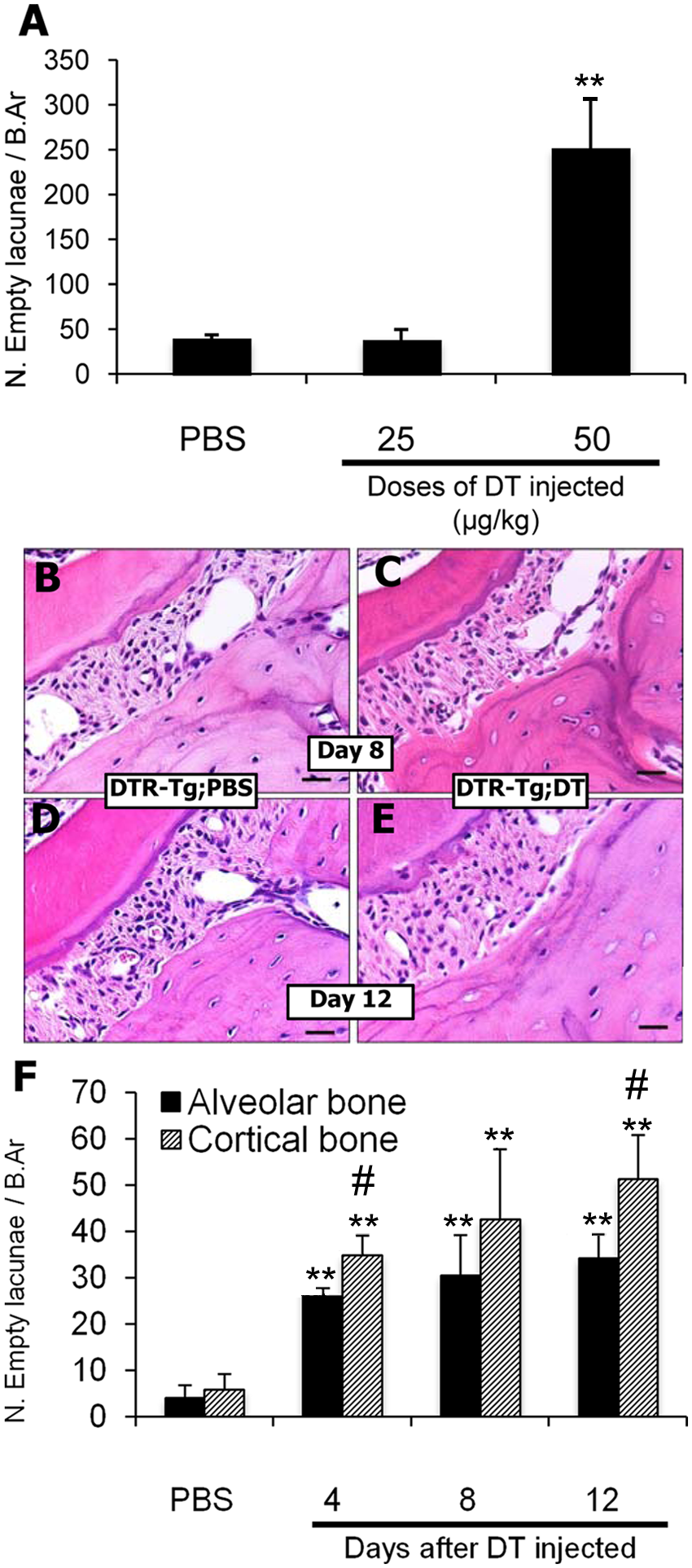
Effect of DT on ablation of osteocytes.
DTR-Tg;DT mice showed an increased number of empty lacunae than did the DTR-Tg;PBS mice (Figs. 2B-2F). On days 4 and 8 after the injection, the empty lacunae were localized at the superficial layer of the alveolar bone adjacent to the periodontium (Fig. 2C). On day 12 after DT injection, many empty lacunae were localized at a deeper layer of the alveolar bone (Fig. 2E). In contrast, empty lacunae were rarely observed in alveolar bone in the other groups on days 8 and 12 after injection of PBS or DT (Figs. 2B, 2D). As shown in Fig. 2F, DT treatment increased the percentages of empty lacunae among the total number of lacunae time-dependently in DTR-Tg;DT mice. Time-dependent increases of the empty lacunae were also observed in the cortical bone of tibiae, though values of the percentages were lower in alveolar bone than in tibia on each day examined (Fig. 2F).
Tooth Movement in WT and DTR-Tg Mice with or without DT Treatment
To investigate the effects of osteocyte ablation during orthodontic tooth movement, we injected mice with DT or PBS 4 days before the application of orthodontic force. On day 12 after tooth movement, the μCT analyses clearly showed wider space between the first and second molars in the DTR-Tg;PBS group (Fig. 3A) than in the DTR−Tg;DT group (Fig. 3B). As shown in Fig. 3C, the distances of tooth movement in the DTR-Tg;DT group tended to be smaller than those in the other 3 groups on days 4 and 8, and the distance of tooth movement became significantly smaller in the DTR-Tg;DT group than in the other 3 groups on day 12 (Fig. 3C). These results indicate that tooth movement was disturbed in the osteocyte-ablated mice.
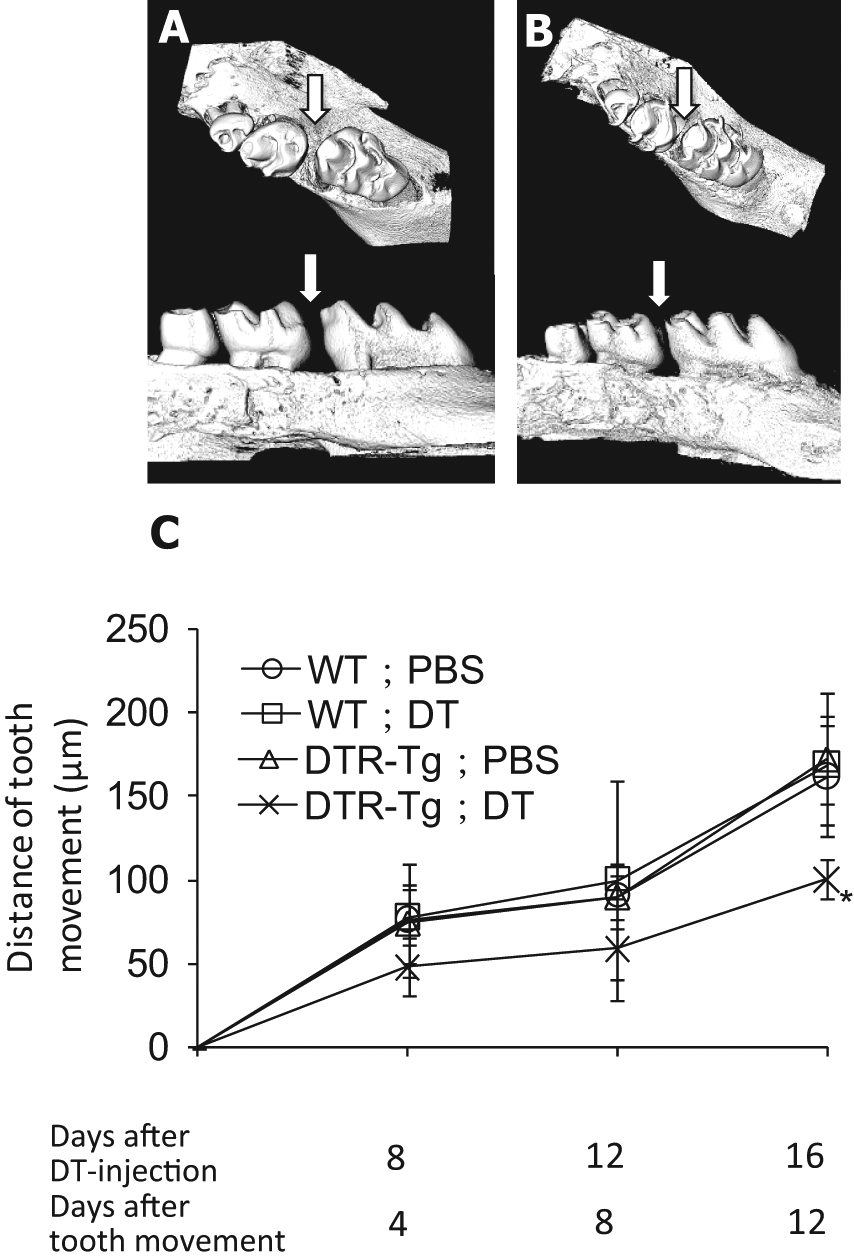
Changes in tooth movement in experimental groups.
Distribution of Osteoclasts at the Compression Site of Alveolar Bone during Tooth Movement
Under a condition without tooth movement, osteoclast numbers at a mesial half area of alveolar bone showed no significant differences among WT;PBS, WT;DT, DTR-Tg;PBS, and DTR-Tg;DT groups on days 4, 8, and 12 after DT injection (Appendix Fig.). In contrast, the numbers of TRAP-positive osteoclasts decreased in the DTR-Tg;DT group (Fig. 4A) compared with that in the DTR-Tg;PBS group (Fig. 4B) at compression sites during orthodontic tooth movement on day 12. Fig. 4C summarizes the time-course of changes in the numbers of TRAP-positive osteoclasts. On day 4, we found no significant differences in the TRAP-positive osteoclast numbers among the groups tested, but on days 8 and 12, the TRAP-positive osteoclast numbers were significantly reduced in the DTR-Tg;DT group compared with the other groups (Fig. 4C). The DTR-Tg;DT group exhibited significantly higher values of eroded bone surface/bone surface (ES/BS), which indicates osteoclast function, than did the other 3 groups on days 4 and 12 after tooth movement (Fig. 4D). These results indicate that osteoclastogenesis and osteoclast function were impaired at the compression site in the DTR-Tg;DT group.
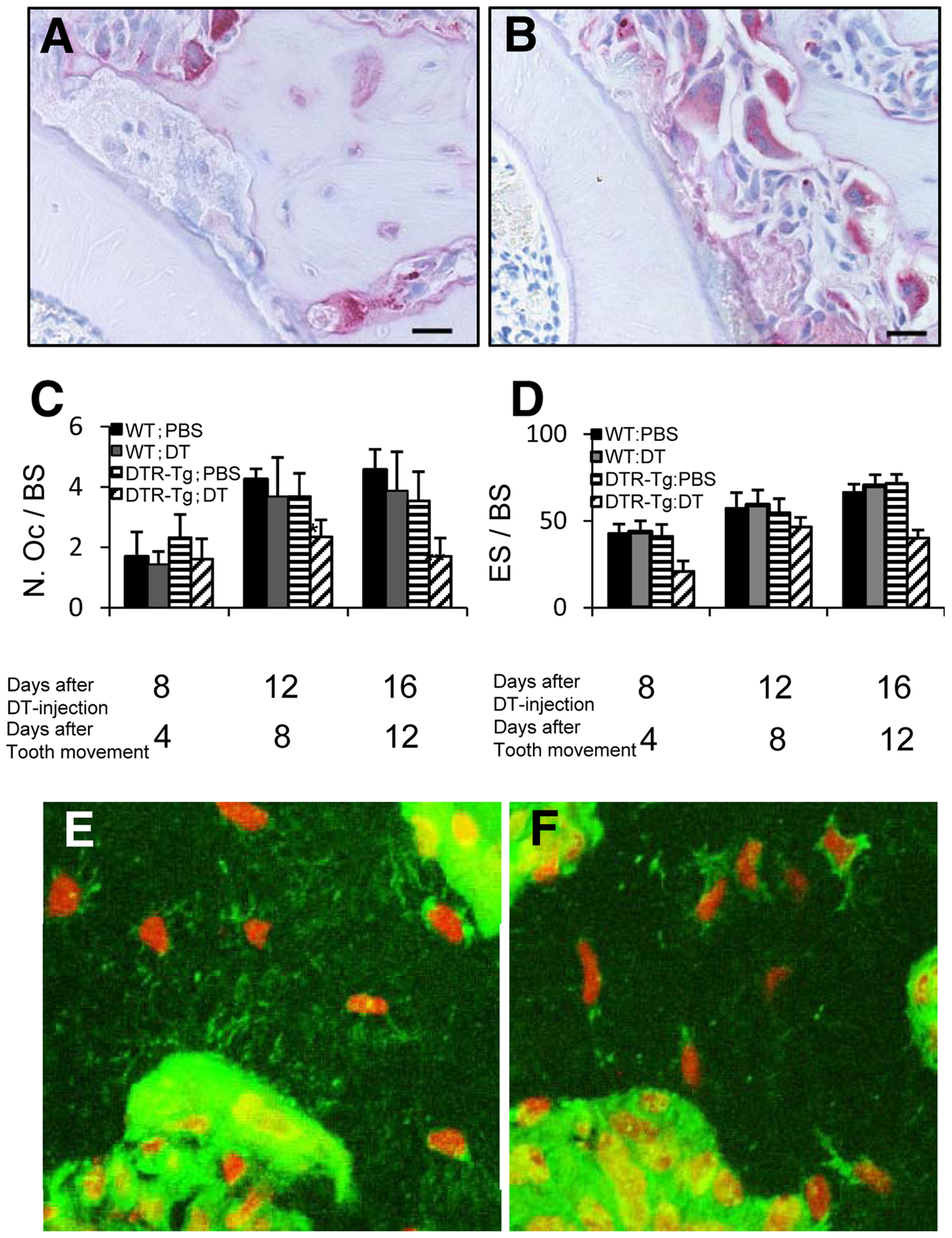
The distribution of TRAP-positive osteoclast and osteocyte canaliculi.
Morphological Interaction between Osteocytes and Osteoclasts at Alveolar Bone during Tooth Movement
Well-developed lacunocanalicular systems of osteocytes were observed in DTR-Tg;PBS mice 4 days after tooth movement (Fig. 4E). At the periphery of the bone adjacent to the bone marrow, several cell processes were in contact with cells on the bone surface, including large osteoclast-like cells (Fig. 4E). These suggest that a direct morphological interaction exists between osteocytes and osteoclasts. In contrast, lacunocanalicular systems of osteocytes were often disrupted in DTR-Tg;DT mice 4 days after tooth movement (Fig. 4F), suggesting the lack of direct contact with osteocytes and osteoclasts in these regions.
Discussion
Osteoclastic bone resorption is an essential event for orthodontic tooth movement. Although no significant differences were observed in osteoclast numbers on day 4 after tooth movement in each group examined, the osteoclast numbers significantly increased on days 8 and 12 compared with those on day 4 in the WT;PBS, WT;DT, and DTR-Tg;PBS groups, indicating that further progression of osteoclastic bone resorption had occurred after day 8 (Fig. 4C). However, DTR-Tg;DT mice showed significantly decreased osteoclast numbers on days 8 and 12 compared with those in other groups on each day (Fig. 4C). Since DT had been injected 4 days before the application of orthodontic force, day 4 after orthodontic tooth movement coincided with day 8 after DT injection. We showed that DT injection induced a substantial increase in the number of empty lacunae from day 4 to day 12 (Fig. 2F), indicating that an adequate amount of osteocyte ablation had occurred when orthodontic force was applied in the DTR-Tg:DT group. However, our experiment demonstrated that the significant inhibition of tooth movement occurred only on day 12, but not on days 4 and 8. This suggests that osteocytes are not critically involved in the early phase of tooth movement, which is considered to represent deformation of investing tissues without active participation of bone remodeling, and that living osteocytes may play an important role in the later stage of orthodontic tooth movement.
Tatsumi et al. (2007) described the increased numbers of osteoclasts at trabecular bone of femurs in DTR-Tg;DT mice by showing TRAP-stained histology without a histomorphometric analysis. In the present study, we showed no significant changes in osteoclast numbers in alveolar bones between the DTR-Tg;PBS and DTR-Tg;DT groups (Appendix Fig.). These results suggest that osteocyte ablation exerts a different action on osteoclast numbers between the femur and alveolar bones in conditions of physiological mechanical loading. We showed decreased numbers of osteoclasts at the compression site in DTR-Tg;DT mice during orthodontic tooth movement (Figs. 4A-4D), and Tatsumi et al. (2007) described how mechanical unloading induced decreased numbers of osteoclasts in the femur. These results suggest that osteocyte ablation decreases osteoclast numbers depending on mechanical stimuli, regardless of unloading or overloading.
Recent studies have demonstrated the importance of osteocytes in osteoclastogenesis (Nakashima et al., 2011; Xiong et al., 2011). Nakashima et al. (2011) reported that osteocytes are the major source of RANKL in bone remodeling. These findings are consistent with our findings, because osteocyte ablation resulted in reduced numbers of osteoclasts during orthodontic tooth movement. In addition, our examination of osteocytes by phalloidin staining showed a direct interaction between osteocytes and osteoclasts. The 9.6-kb mouse Dmp1 promoter is strongly activated in osteocytes and odontoblasts (Lu et al., 2007), but a recent study (Kalajzic et al., 2012) reported that this promoter is activated in osteoblasts, bone marrow cells, muscle tissues, brain, and kidney as well as osteocytes. Thus, it is important to distinguish the role of RANKL synthesized by osteocytes or osteoblasts during orthodontic tooth movement in different animal models.
It has been suggested that not only healthy but also dying osteocytes can recruit osteoclasts to sites of remodeling (Bonewald, 2011). Apoptosis and necrosis are different forms of cell death. Osteocyte apoptosis can trigger bone remodeling by stimulating osteoclastogenesis, because the apoptotic bodies derived from osteocytes are associated with potent osteoclastogenic factors (Kogianni et al., 2008). Although the targeted ablation of osteocytes used in this study was thought to occur through necrosis (Bonewald, 2011), the original reports describing DTR-Tg mice (Tatsumi et al., 2007) and analysis of our data clearly show that osteocyte ablation in these mice caused changes in osteoclastogenesis in response to mechanical stimuli. The mechanisms underlying osteocyte necrosis-related osteoclastogenesis should be investigated in more detail to clarify the mechanisms by which osteocytes regulate osteoclast recruitment.
Footnotes
This work was supported by Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science to K. M. and A. Y., and by the Global Center of Excellence (GCOE) Program; International Research Center for Molecular Science in Tooth and Bone Diseases, Tokyo Medical and Dental University.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.