
Abstract
Chitosan is a naturally derived polymer with antimicrobial and anti-inflammatory properties. However, studies evaluating the role of chitosan in the control of periodontal pathogens and the responses of fibroblasts to inflammatory stimuli are lacking. In the present study, we analyzed whether chitosan particles may inhibit the growth of periodontal pathogens and modulate the inflammatory response in human gingival fibroblasts. Chitosan particles were generated through ionic gelation. They inhibited the growth of Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans at 5 mg/mL. Conversely, IL-1β strongly stimulated PGE2 protein levels in gingival fibroblasts, and chitosan inhibited this response at 50 µg/mL. IL-1β–stimulated PGE2 production was dependent on the JNK pathway, and chitosan strongly inhibited this response. IL-1β stimulated NF-κB activation, another signaling pathway involved in PGE2 production. However, chitosan particles were unable to modify NF-κB signaling. The present study shows that chitosan exerts a predominantly anti-inflammatory activity by modulating PGE2 levels through the JNK pathway, which may be useful in the prevention or treatment of periodontal inflammation.
Introduction
Oral micro-organisms are involved in the pathogenesis of periodontal diseases (Pihlstrom et al., 2005). Nevertheless, the host response that includes the production of cytokines and prostanoids plays a key role in periodontitis (Paquette and Williams, 2000).
Chitosan corresponds to a polysaccharide derived from chitin (Domard, 2011). It has been studied for its applications as a biomaterial for tissue regeneration, drug delivery, antibacterial, and anti-inflammatory purposes (Francesko and Tzanov, 2011). Studies have reported that chitosan may inhibit the growth of Porphyromonas gingivalis (Pg) and Aggregatibacter actinomycetemcomitans (Aa), among others (Choi et al., 2001; Ikinci et al., 2002; Sarasam et al., 2008; Costa et al., 2012). However, it is interesting to note that no previous studies have evaluated the antimicrobial and anti-inflammatory properties of one single chitosan formulation. This is important, since chitosan is prepared from a natural source, and the above-indicated biological activities may show substantial variations between and among different formulations (Domard, 2011).
Chitosan may exert an anti-inflammatory activity in astrocytoma cells (Kim et al., 2002) and macrophages (Chou et al., 2003; Yoon et al., 2007). However, these effects may vary significantly depending on the different chitosan formulations and the tissue under study (Huang et al., 2005). In this regard, no data are available concerning the biological effects and the mechanism of action of chitosan in fibroblasts.
The inflammatory mediator PGE2 plays a prominent role in the pathogenesis of periodontitis (Noguchi and Ishikawa, 2007), and gingival fibroblasts stimulated with IL-1β produce large amounts of PGE2 (Yucel-Lindberg et al., 1999). In the present study, we evaluated the antibacterial activity of chitosan particles against Pg and Aa and their capacity to modulate PGE2 production in fibroblasts stimulated with IL-1β. Moreover, we analyzed some of the signaling pathways modulated by chitosan in gingival fibroblasts.
Materials & Methods
Preparation of Chitosan Nanoparticles
Nanoparticles were prepared by the ionic gelation of chitosan (chitosan, low-molecular-weight; Sigma-Aldrich, St. Louis, MO, USA) with tripolyphosphate (TPP) anions (Mohammadpourdounighi et al., 2010). Briefly, a 3.0-mg/mL quantity of chitosan was dissolved in acetic aqueous solution. A 5-mL chitosan solution was added dropwise under constant stirring to 2 mL of a 1.0 mg/mL TPP solution. Nanoparticles were separated by centrifugation at 13,000 rpm at 14°C for 30 min, freeze-dried, and stored at 4°C. Particles were milled in a mortar and sieved through a 400 mesh. The morphologic and physicochemical features of these particles have been described elsewhere (Silva et al., 2013).
Antibacterial Effects of Chitosan Particles
Aggregatibacter actinomycetemcomitans (Aa) (ATCC 29522) and Porphyromonas gingivalis (Pg) (ATCC BAA-308) cultures were established in BHI broth medium (Thermo Scientific, Oxoid, Sunnyvale, CA, USA) supplemented with 40 µg/mL NaHCO3 (Aa) and 10 μg/mL vitamin K, 5 µg/mL hemin, and 0.5% yeast extract (Pg). Both types of bacteria were grown in the presence, or not, of 10 μg/mL, 50 μg/mL, 100 μg/mL, 150 μg/mL, 200 μg/mL, 500 μg/mL, 1 mg/mL, 5 mg/mL, and 10 mg/mL chitosan. BHI broth medium was used as a negative control. Chlorhexidine (Sigma) (0.12%) was used as a positive control. Aa cultures were grown at 37°C under micro-aerophilic conditions. Pg cultures were grown in anaerobiosis in an anaerobic jar (Thermo Scientific). To evaluate the bacterial concentration in suspension, we measured the optical density (OD) of the medium spectrophotometrically.
Primary Cultures of Human Gingival Fibroblasts
Primary cultures of human gingival fibroblasts (GF) were performed by the explant method. Tissue explants were obtained from the masticatory mucosa at the retromolar area from four adults at the School of Dentistry, Pontificia Universidad Católica de Chile. Second molars were examined, and tissue samples with local inflammation were discarded. None of the examined tissues was from sites showing bleeding on probing, probing depths greater than 3 mm, or clinical attachment level greater than 1 mm. Tissue samples were harvested with the informed consent of the patients. The protocol for tissue acquisition was approved by the Ethical Committee, Faculty of Medicine, Pontificia Universidad Católica de Chile. Patients reported no relevant medical or drug histories during the preceding 6 mos. Cells were cultured in α-Minimal Essential Medium (α-MEM) (Gibco BRL, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS) (Hyclone Laboratories Inc., Logan, UT, USA) and Normocin (InvivoGen, San Diego, CA, USA) at 37oC and 5% CO2. All experiments were performed with cells expanded between passages 4 and 10.
Cell Viability Assays
Cell viability of gingival fibroblasts exposed to chitosan particles was evaluated through the release of lactate dehydrogenase (LDH) and the MTS assay (Promega, Madison, WI, USA).
PGE2 Production in Human Gingival Fibroblasts
GF (4,000/cells per well) were seeded in 96-well plates (Orange, Braine-l’Alleud, Belgium). To evaluate the anti-inflammatory potential of chitosan particles, we exposed serum-starved GF to 50, 200, or 400 µg/mL chitosan overnight. After this, we added 300 pg/mL IL-1β (R&D Systems, Minneapolis, MN, USA). This IL-1β concentration was obtained after we performed a pilot study in which cells were stimulated with this cytokine in concentrations ranging between 100 pg/mL and 10 ng/mL. After 16 hrs of stimulus, PGE2 was measured in the cell culture media through ELISA. Conditioned media was centrifuged at 15,000 rpm for 10 min, and PGE2 levels were measured by a competitive ELISA assay (Thermo Scientific). Results were normalized against viable cells by the MTS assay.
Western Blot
After appropriate stimulation, cells underwent lysis as indicated by Arancibia et al. (2009). Cell lysates were centrifuged (19,000 g, 10 min, 4°C), and proteins were resolved through 10% sodium dodecyl sulfate–polyacrylamide gel and transferred to polyvinylidene difluoride membranes (PerkinElmer Life Sciences, Boston, MA, USA). Membranes were exposed to primary antibodies against phosphorylated c-Jun-N-terminal kinase (JNK) (Cell Signaling Technology, Danvers, MA, USA), JNK (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and beta-actin (Sigma), then to secondary antibodies coupled to horseradish peroxidase (Jackson ImmunoResearch, West Grove, PA, USA) and finally developed by means of an enhanced chemiluminescence kit (PerkinElmer Life Sciences).
Immunofluorescence
We plated 4 x 104 cells on glass coverslips. Serum-starved GF were exposed to 500 µL α-MEM supplemented with 200 µg/mL chitosan, 10 µg/mL dexamethasone (Sigma), or 10 µM SP600125 (Biolmol, Plymouth, PA, USA) overnight. After this, cells were stimulated with 300 pg/mL IL-1β. After 3 hrs, cells were fixed with 4% paraformaldehyde and processed for immunofluorescence according to Arancibia et al. (2009). Primary antibodies used were anti-Phospho-c-Jun (Merck-Millipore, Billerica, MA, USA) and anti-NF-κB (Santa Cruz Biotechnology). Cells were then incubated with fluorescent secondary antibodies (Invitrogen Molecular Probes, Carlsbad, CA, USA). Images were collected by immunofluorescence microscopy (Carl Zeiss, Dresden, Germany) and photographed with a digital camera (Carl Zeiss). Nuclei were stained with Hoescht (Invitrogen Molecular Probes). Nuclei stained for phospho-cJun or p65 were counted in 5 random fields. Positive nuclei/total nuclei ratios were calculated for statistical analysis.
Statistical Analysis
For statistical comparison between and among conditions under study, the Mann-Whitney U test was used (Figs. 2, 3A, 4B, 4D). To analyze data in Fig. 3D, we used the Student’s t test. Statistical significance was set at p < .05. SPSS software for Windows was used (version 16.0.2) (SPSS Inc., Chicago, IL, USA).
Results
The antibacterial activity of chitosan particles against periodontal pathogens was evaluated by growth curves for Aa and Pg in the presence of different chitosan concentrations (10 μg/mL to 10 mg/mL) obtained from previous studies (Choi et al., 2001; Ikinci et al., 2002; Sarasam et al., 2008; Costa et al., 2012). Chitosan was able to inhibit the growth of both Aa and Pg at 5 mg/mL (Fig. 1).
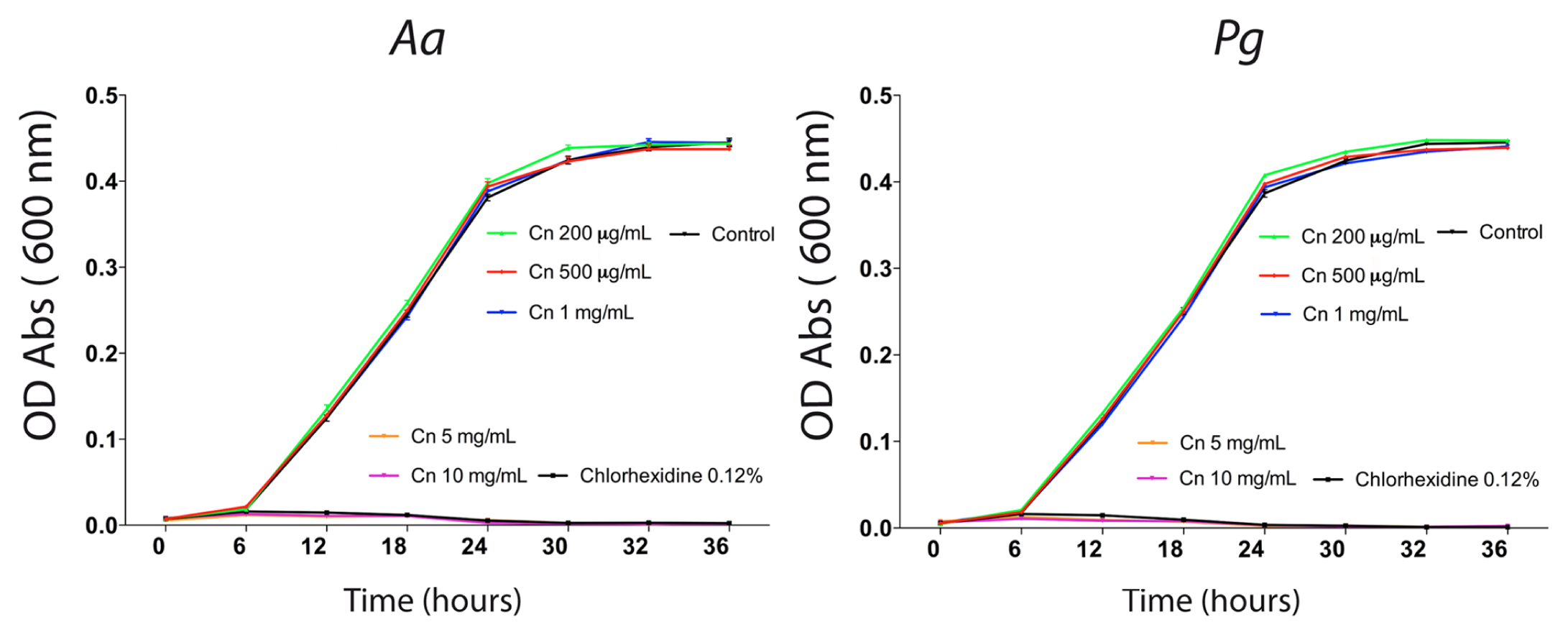
Effects of chitosan (Cn) particles on Aa and Pg. Aggregatibacter actinomycetemcomitans (Aa) and Porphyromonas gingivalis (Pg) were grown in BHI broth medium in the presence, or not, of 10 μg/mL, 50 μg/mL, 100 μg/mL, 150 μg/mL, 200 μg/mL, 500 μg/mL, 1 mg/mL, 5 mg/mL, and 10 mg/mL of chitosan. BHI broth medium was used as a negative control. Chlorhexidine digluconate (0.12%) was used as a positive control. Bacterial concentration in suspension was measured through detection of optical density (OD) of the medium by spectrophotometry at 0, 6, 12, 18, 24, 30, 32, and 36 hrs. Graphs represent optical density means plus standard deviation.
To evaluate the anti-inflammatory potential of chitosan particles, we exposed serum-starved GF to a range of chitosan concentrations overnight. After this, a 300-pg/mL quantity of IL-1β was added for 16 hrs. PGE2 was measured in the cell culture media through ELISA. IL-1β induced a significant stimulus in PGE2 protein levels, and all the chitosan concentrations tested showed significantly diminished PGE2 levels (Fig. 2). We also observed that chitosan alone was unable to induce changes in PGE2 levels in the absence of other stimuli (data not shown). Moreover, IL-1β did not affect cell viability as determined through the MTS assay. However, 5 mg/mL chitosan induced a decrease in the MTS assay and an increase in the release of LDH into the cell culture media (Appendix Fig.).
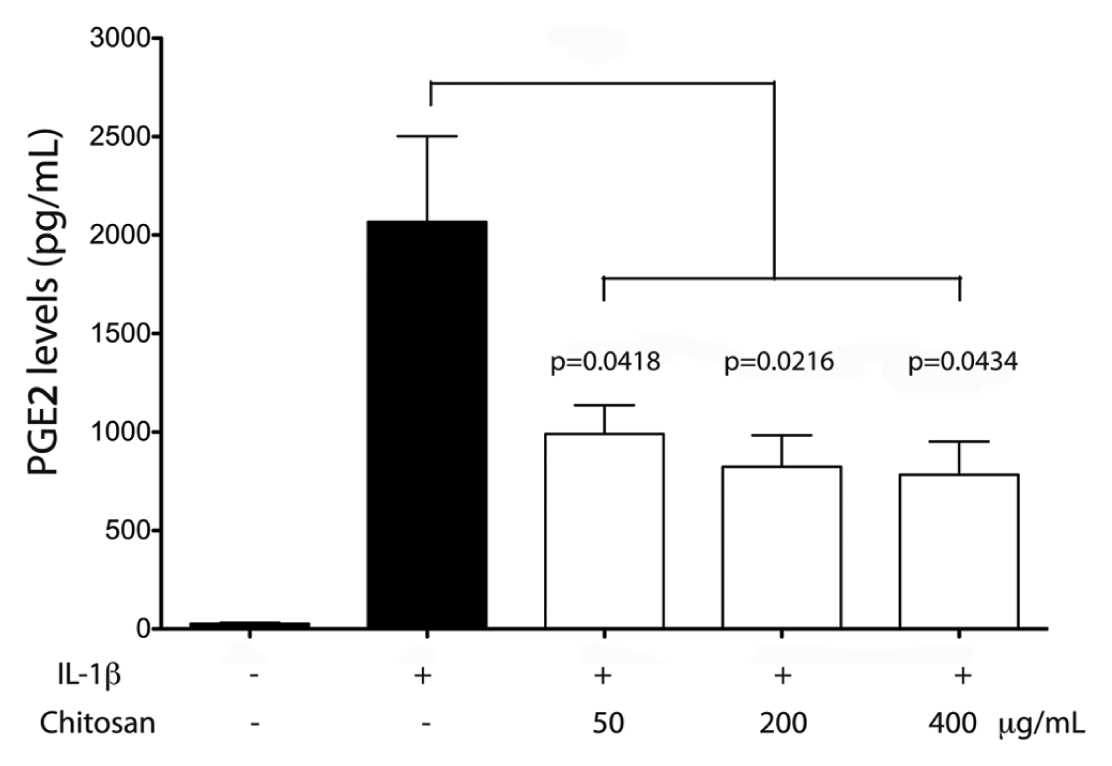
Effect of chitosan on PGE2 levels in gingival fibroblasts. Serum-starved gingival fibroblasts were exposed to chitosan (50, 200, or 400 μg/mL) overnight and then stimulated with 300 pg/mL of IL-1β. An ELISA assay was used to detect PGE2 levels in the cell culture media. Data were obtained from 5 independent experiments. Data were normalized against viable cells at the end of the experiment through an MTS assay. Graph represents PGE2 mean values plus standard error. Brackets indicate statistically significant differences between and among the different experimental conditions.
To explore some of the signaling pathways involved in the regulation of PGE2 by chitosan, we incubated serum-starved GF in the presence of 10 µM SP600125, a selective inhibitor of the JNK signaling pathway implicated in PGE2 production (Båge et al., 2010). Dexamethasone was used as a positive control of cytokine-stimulated PGE2 inhibition (Yucel-Lindberg et al., 1999). Cells were exposed overnight to the above-indicated drugs, or to 200 µg/mL chitosan, and then stimulated with 300 pg/mL IL-1β. After 16 hrs, PGE2 levels were measured through ELISA. All the agents tested effectively diminished IL-1β–stimulated PGE2 protein levels, suggesting that the JNK pathway was involved in PGE2 production (Fig. 3A). We then analyzed whether IL-1β was able to activate the JNK pathway. IL-1β–stimulated GF showed an increase in JNK phosphorylation, with a maximum at 30 min of treatment (Fig. 3B). To analyze whether chitosan was able to modulate IL-1β–stimulated JNK phosphorylation, we stimulated serum-starved GF with IL-1β in the presence, or not, of chitosan particles. Chitosan significantly diminished JNK phosphorylation (Figs. 3C, 3D). We then tested whether phosphorylated c-Jun, the product of JNK (Davies and Tournier, 2012), was modulated by chitosan. To this end, serum-starved GF were exposed to 200 µg/mL chitosan, 20 µg/mL dexamethasone, or 10 µM SP600125 prior to IL-1β stimulation. The nuclear location of phosphorylated-c-Jun was evaluated through immunofluorescence. Chitosan, dexamethasone, and SP600125 effectively diminished the abundance of nuclear phosphorylated-c-Jun (Figs. 4A, 4B).
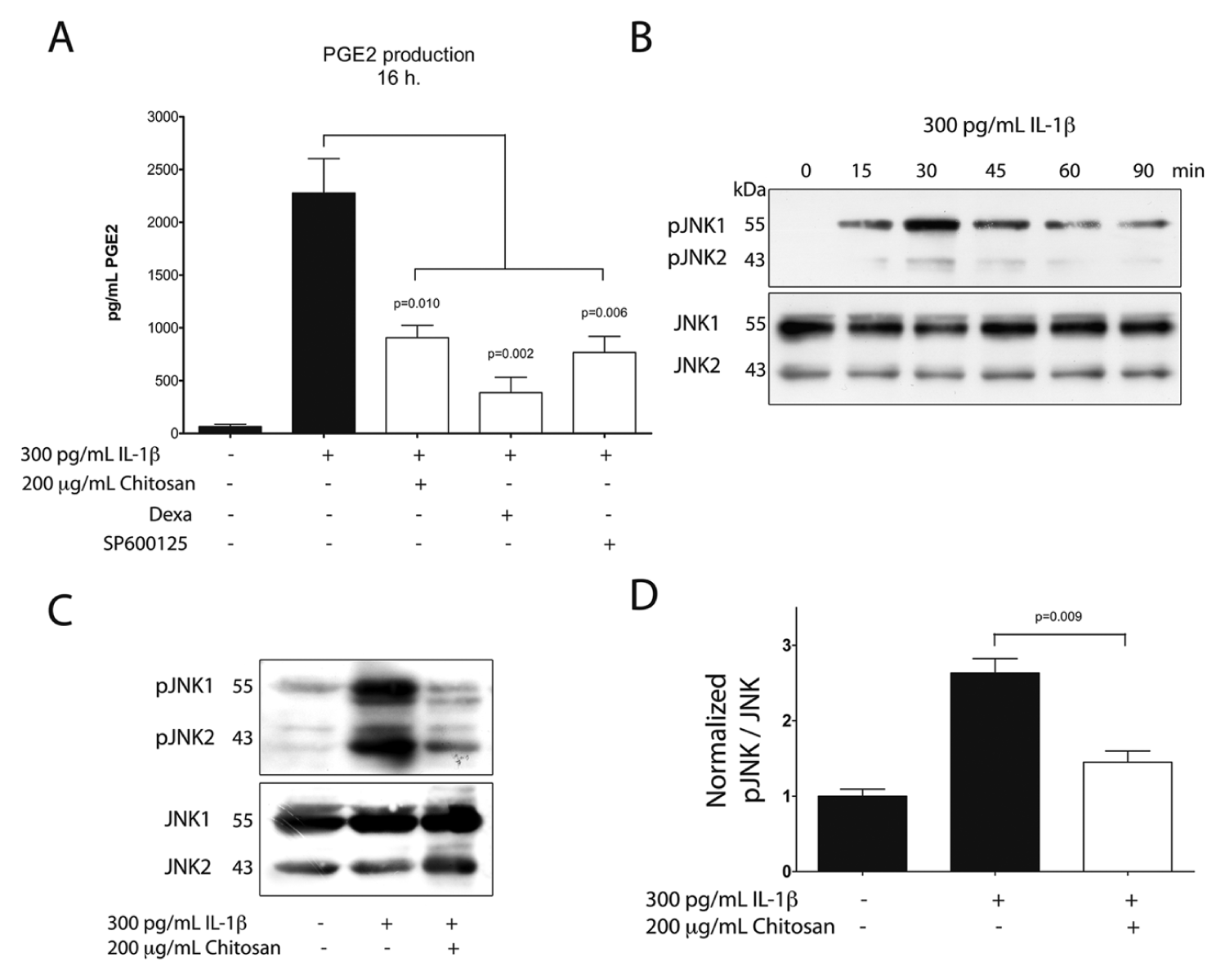
Role of JNK in IL-1β–stimulated PGE2 production.
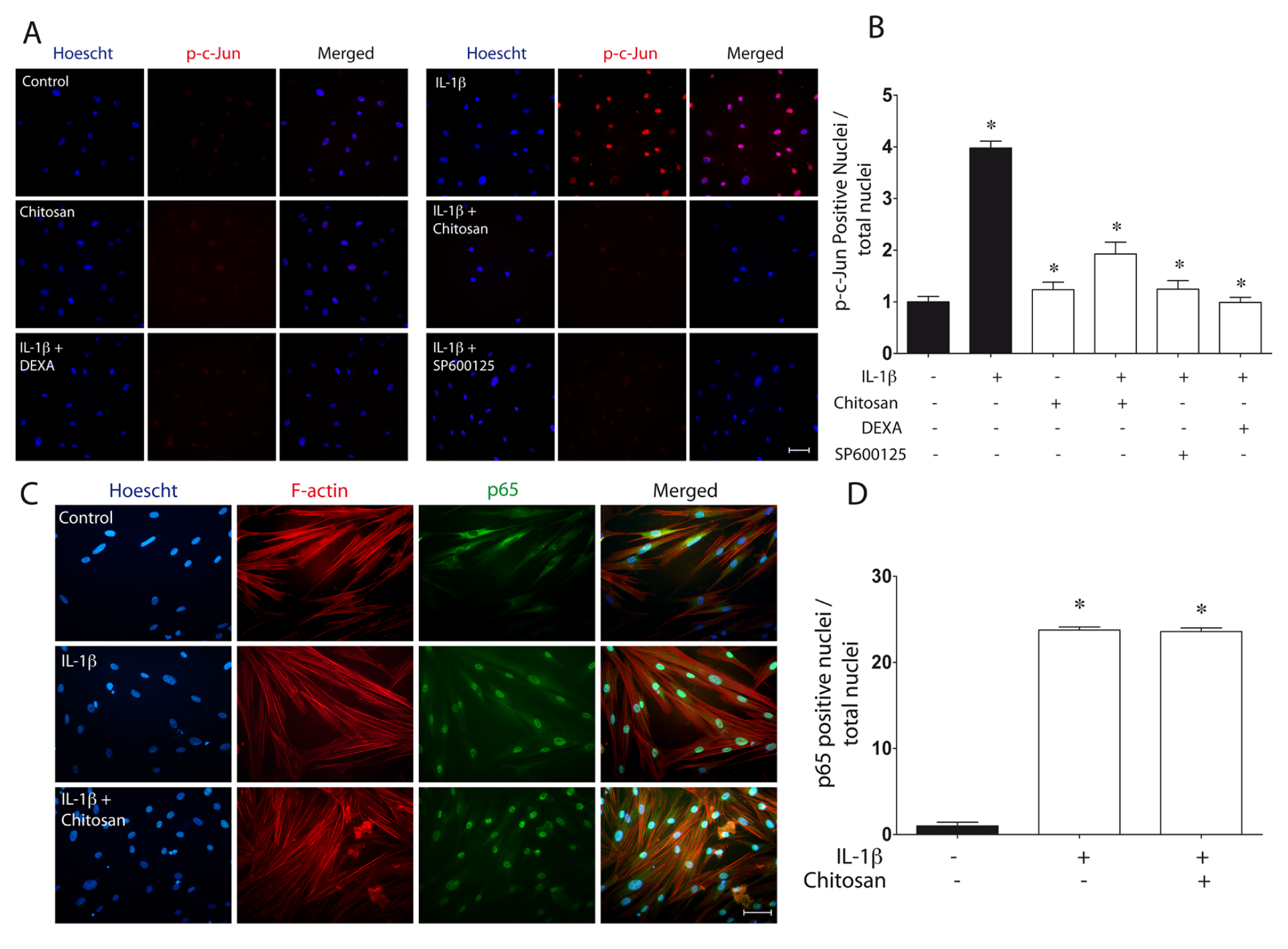
Regulation of c-Jun and p65 translocation by chitosan.
Given the prominent role played by the NF-κB pathway in the regulation of PGE2 production (Båge et al., 2010), we evaluated whether NF-κB was modulated by chitosan. Using immunofluorescence, we monitored the nuclear translocation of the p65 subunit of NF-κB as an indicator of the activation of this pathway. IL-1β stimulated NF-κB nuclear translocation (Figs. 4C, 4D). However, chitosan was unable to reverse this response, suggesting that this polymer does not affect or modulate the NF-κB signaling pathway.
Discussion
Periodontal disease involves the interplay of infecting periodontal pathogens and the host inflammatory response, leading to tissue destruction (Pihlstrom et al., 2005). Chitosan is an attractive polymer that may have both antibacterial and anti-inflammatory properties. However, the great variability observed in chitosan samples, including degrees of deacetylation, molecular weight, viscosity, and pKa, leads to difficulties when different studies are analyzed (Domard, 2011). To our knowledge, this is the first study that evaluates the antibacterial and anti-inflammatory properties of the same chitosan particles. The present results show that chitosan may reduce the IL-1β–stimulated PGE2 protein levels at 50 µg/mL and inhibit the growth of Aa and Pg at 5 mg/mL. Cell viability assays also demonstrated that chitosan may induce a cytotoxic effect at 5 mg/mL, but not at 1 mg/mL, in gingival fibroblasts. Therefore, chitosan may induce cell damage in both bacterial and connective tissue cells at this concentration. Further studies are needed to determine a chitosan concentration that may inhibit bacterial growth without affecting the survival of gingival cells.
The results of the present study show that chitosan is able to reduce the protein levels of PGE2, an important inflammatory mediator involved in tissue destruction in periodontitis (Offenbacher et al., 1993; Preshaw and Heasman, 2002). In addition, selective inhibition of PGE2 by anti-inflammatory drugs decreases periodontal disease progression (Williams et al., 1989; Salvi and Lang, 2005). GFs constitute the predominant cell type in gingival connective tissue and are involved in the production of PGE2 in diseased gingiva (Yucel-Lindberg et al., 1999; Båge et al., 2011). Therefore, the present results suggest that chitosan may be considered as a therapeutic tool for the modulation of inflammation in the periodontium. Future studies should evaluate whether these anti-inflammatory effects are also achieved in vivo.
PGE2 production involves the sequential activity of different groups of enzymes, including phospholipase A2, cyclooxygenases-2, and prostaglandin E synthases (PGEs) (Båge et al., 2011). COX-2 and mPGES-1 have been proposed as the main enzymes involved in PGE2 production in IL-1β−stimulated GF (Båge et al., 2011). Our experiments did not explore the level at which chitosan modulated PGE2 production. Future studies should evaluate whether chitosan affects COX-2 or mPGES-1, among others.
Our study showed that chitosan was able to reduce JNK phosphorylation in GF stimulated by IL-1β. In addition, GFs demonstrated an increase in phosphorylated-c-Jun, and chitosan also diminished this response. c-Jun is a component of the transcription factor AP-1, which is phosphorylated by JNK (Davies and Tournier, 2012). Therefore, the present results strongly suggest that the JNK pathway, stimulated by IL-1β, can be modulated by chitosan. Previous studies have reported the modulation of mitogen-activated protein kinase (MAPK) pathways by chitosan. However, it is not clear whether these effects are anti- or pro-inflammatory. For instance, in osteoblasts, chitosan oligomers may stimulate MAPK signaling pathways (Ganno et al., 2007). Chitosan plays an anti-inflammatory role in human astrocytoma cells by modulating the stimulus induced by amyloid β peptide or IL-1β (Kim et al., 2002). In microglia cells, chitosan may inhibit the inflammatory response by down-regulating MAPK signaling (Pangestuti et al., 2011). In RAW264.7 macrophages, chitosan oligosaccharides inhibit LPS-stimulated PGE2 production through the inhibition of COX-2 (Chou et al., 2003; Yoon et al., 2007). However, in a macrophage-like cell line, stimulation of these cells with chitin induced tumor necrosis factor and nitric oxide production (Shimojoh et al., 2010). Moreover, chitosan may even stimulate inflammation in the lung (Huang et al., 2005). To our knowledge, this is the first report that describes the role of chitosan as an anti-inflammatory agent in fibroblasts and provides evidence that activation of the JNK pathway can be a target for this anti-inflammatory agent. However, it cannot be ruled out whether other MAPKs, such as extracellular signal regulated kinase (ERK) or p38, may be involved in IL-1β–stimulated PGE2 production as well.
The nuclear factor-kappa B (NF-κB) transcription factor exerts a key role in the regulation of inflammation by modulating COX-2 and mPGES-1, among other mediators (Båge et al., 2010). Glucocorticoids are potent NF-κB inhibitors, and this may partly explain the inhibitory effect of dexamethasone on IL-1β–stimulated PGE2 production (Murphy et al., 2011). Contradictory studies have shown that chitosan may stimulate or inhibit NF-κB signaling. These divergent responses may be due to differences in chitosan formulations, cell types under study, and the nature of the inflammatory stimuli (Yu et al., 2004; Wu and Tsai, 2007; Wei et al., 2012). Our results showed that the NF-κB pathway was not modulated by chitosan particles. Further studies are needed to understand the roles of, and interplay between, NF-κB and JNK in the regulation of inflammatory responses and the effect of chitosan in this response.
The results of the present study contribute to our understanding of the mechanism of action of chitosan particles on the growth of periodontal pathogens and the control of inflammatory responses in human gingival fibroblasts. These findings are relevant for the design and future testing of chitosan-based formulations for the prevention and treatment of periodontal diseases.
Footnotes
This study was financed by
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.