
Abstract
Lymphatic vessels return extravasated fluid, proteins, and cells back into the circulation and are important in immune cell trafficking. In the gingiva, lymphatic vessels are located in the lamina propria and travel over the external surface of the alveolar bone. The gingival lymphatics are important for fluid drainage, since lack of lymphatics has been shown to increase interstitial fluid pressure and fluid volume. Maintenance of gingival lymphatic vessels requires continuous signaling by the growth factors VEGF-C and -D via their receptor VEGFR-3. The growth factors are expressed in the gingival epithelium and also in immune cells in the lamina propria. VEGF-C seems to be crucial for lymphangiogenesis induced during periodontal disease development. The lymphatic vessels protect against periodontitis in mice, probably by clearing bacteria and bacterial products and by promoting humoral immune responses. Down-regulation of CCL21, a ligand important for dendritic cell migration, has been demonstrated in lymphatics from patients with periodontitis. High enzymatic activity in the gingiva of these patients may also contribute to impaired lymphatic function, due to the loss of structural components in the interstitium influencing lymphatic function. So far, knowledge is limited in this field because of the dearth of studies on the role of lymphatic vessels in periodontal disease.
Keywords
Introduction
The gingiva is frequently challenged by oral bacteria that are able to induce inflammatory responses, which results in increased fluid filtration into the interstitial space and edema formation in the tissue. Chronicity of the inflammatory response may be followed by the development of periodontitis, characterized by bone resorption. The lymphatic vasculature forms a vessel network in the interstitium and plays an important homeostatic role in the body. This network drains filtered fluid and proteins from the tissue and returns them to the blood. In addition, it serves an important role in the body’s immune defense, since activated antigen-presenting cells, which enter the blind-ended lymphatic capillaries, are transported to lymph nodes and elicit immune responses. The lymphatic endothelial cells interact with immune cells and participate in inflammatory responses, thereby acting as active partners in the peripheral tissue’s interplay (Alitalo et al., 2005). Lymphatic research has developed rapidly since the first specific lymphatic markers were detected, but still we have limited information about their role in normal tissue homeostasis in the oral cavity and their role when the tissue is challenged by inflammatory insults. Here we review the role of lymphatic vessels in tissue fluid balance and also their known functions as part of the body’s immune system. The lymphatic responses in inflammation are also included. Furthermore, we give an overview of the recent research in this field in periodontal tissues. The dental pulp and pulpal disease with development of apical periodontitis are not described, since lymphatic vessels are not found in these locations. This review also points to the direction for future research in this field, a field in which insight remains limited because of the lack of investigations thus far.
Structure and Function of Lymphatics
The lymphatic system consists of lymphatic vessels and lymphoid organs. Lymphatic vessels return extravasated fluid, proteins, and cells to the circulation and are important in immune cell trafficking and the absorption of dietary fat (Alitalo, 2011; Wiig and Swartz, 2012) (Fig. 1). Some avascular tissues, such as epidermis, cartilage, and cornea, and some vascularized organs, like brain, retina, and the dental pulp, are devoid of lymphatics, but all other organs have blind-ended lymphatic capillaries, also known as initial lymphatics. These vessels transport lymph to larger collecting lymphatics, which again return this fluid to the general circulation in lymphatic-vascular junctions in the cervical area (Mumprecht and Detmar, 2009; Alitalo, 2011).
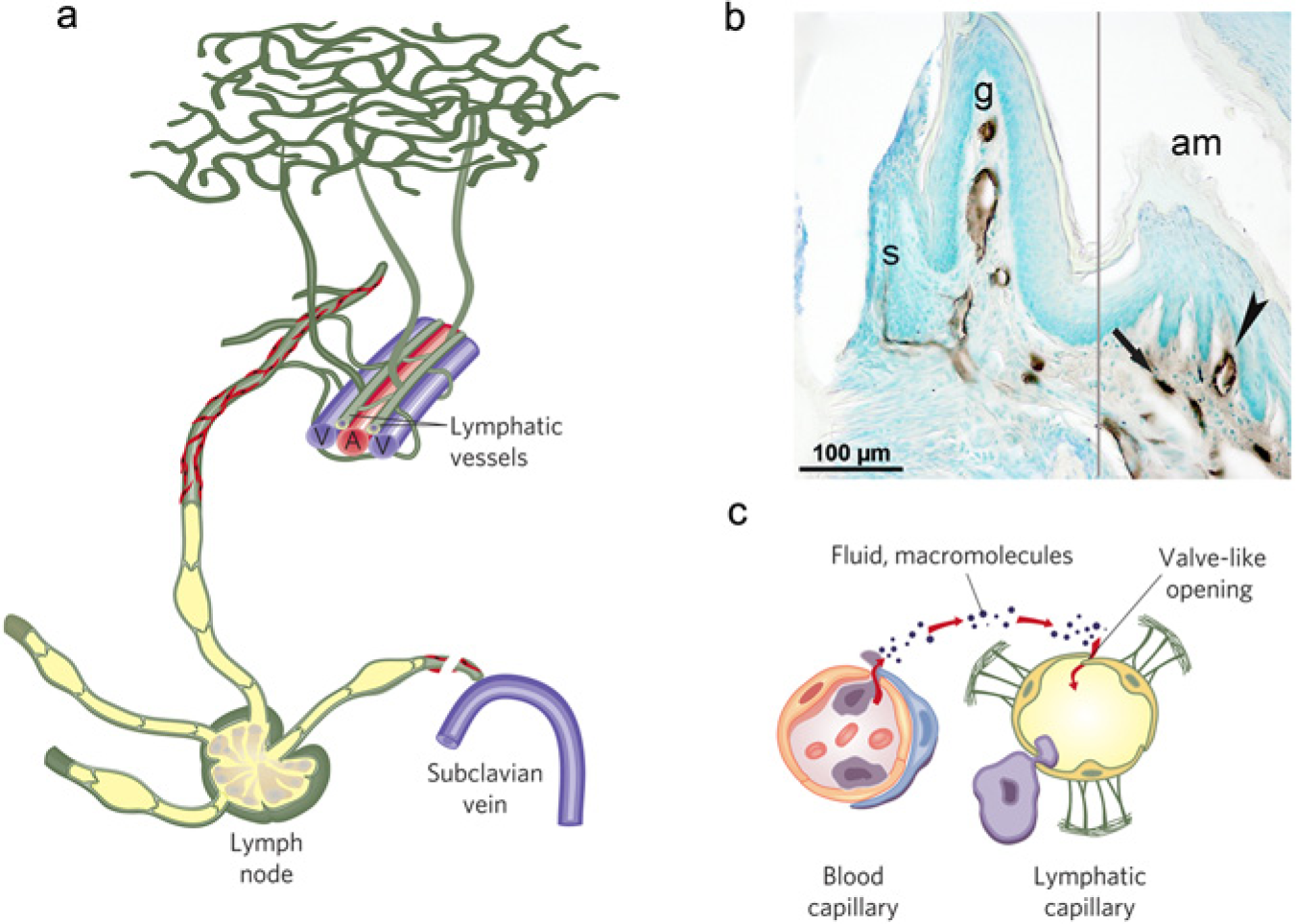
On its way from the tissue to the blood circulation, the lymph travels through the following channels with increasing size: lymph capillaries (also called initial lymphatics), collecting vessels, lymph nodes, trunks, and ducts (Wiig and Swartz, 2012). The initial lymphatics are thin-walled, relatively large vessels (10-60 µm) compared with surrounding blood capillaries (5-10 µm) (Schmid-Schonbein, 1990; Wiig and Swartz, 2012). These vessels are composed of a single layer of endothelial cells called lymphatic endothelial cells (LECs), are not ensheathed by pericytes or smooth-muscle cells, and have little or discontinuous basement membrane. The initial lymphatics are the site of interstitial fluid absorption, and only small and transient pressure gradients are required for the entrance of fluid. A fundamental question concerns the mechanism that causes expansion and compression of the initial lymphatics. It is assumed that the so-called anchoring filaments, connecting elastic fibers in the extracellular matrix (ECM) and the abluminal portion of initial lymphatics (Leak, 1976), as well as tissue stress like tissue expansion due to increased fluid content in the interstitium, contraction of surrounding muscles, and arterial pulsations contributing to initial lymph formation (Schmid-Schonbein, 1990; Wiig and Swartz, 2012), are the mechanisms causing expansion and compression of the initial vessels. The initial lymphatics have overlapping endothelial cells joined together by the junctional protein VE-cadherin in “button-like” patterns (Baluk et al., 2007) that may serve as flaps that allow for one-way absorption of cells, fluid, and proteins, and thus as primary valves in these vessels.
Lymph is formed from fluid filtered across the capillaries that is determined by the imbalance between colloid osmotic (oncotic) and hydrostatic pressure, expressed quantitatively as
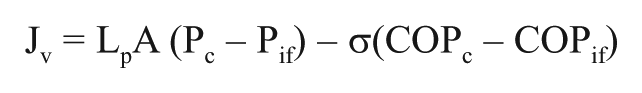
also known as the Starling equation (Levick and Michel, 2010) (Fig. 2). In this equation, Jv is the net capillary filtration, Lp the hydraulic permeability of the capillaries, A the surface area available for filtration, and σ the capillary reflection coefficient. (Pc – Pif) is the hydrostatic pressure difference between plasma in the capillaries (c) and interstitial fluid (Pif), and (COPc – COPif) represents the corresponding difference in colloid osmotic pressures. During control conditions, the lymph flow will be equal to Jv, and thereby the tissue hydration is kept constant, whereas during inflammation, the various parameters will change (Fig. 2), and fluid will accumulate.
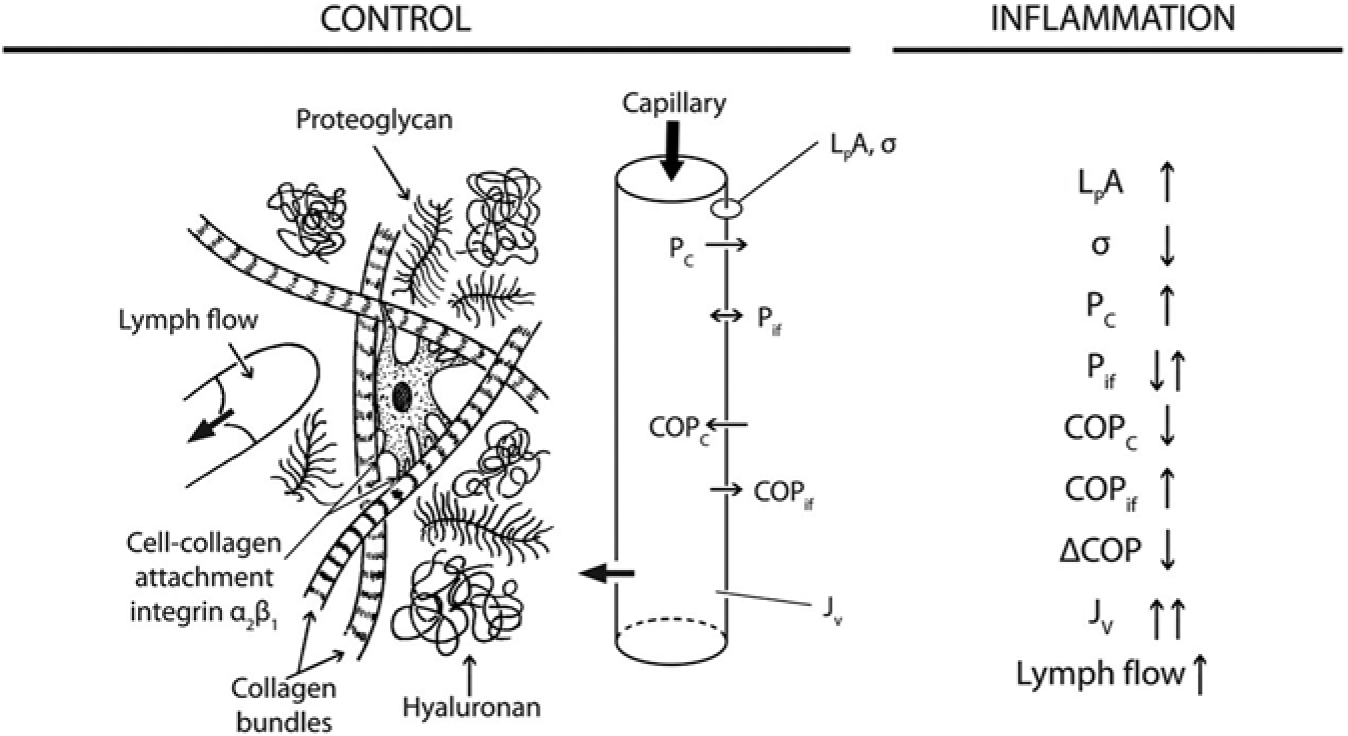
Overview of the transcapillary-interstitial fluid exchange system in a control situation and during inflammation. (
Initial lymph is propagated by tissue stress (Schmid-Schonbein, 1990) from initial to larger collecting lymphatic vessels endowed with smooth-muscle cells and moved centrally by spontaneous contractions. Because of the presence of one-way valves, such transport is unidirectional. Larger collecting vessels are innervated and in addition appear to have pacemaker units (Muthuchamy and Zawieja, 2008; von der Weid and Muthuchamy, 2010). The functional units within the muscular lymphatics are called ‘lymphangions’, which are arranged in series and are delimited by the lymphatic valves (Muthuchamy and Zawieja, 2008). On their central path, lymph in collecting vessels passes through lymph nodes. Lymph may therefore be classified as pre- or post-nodal (or afferent or efferent), depending on whether it is carried to or from the nodes, respectively. This distinction is important, because the lymph composition is changed during its passage through the nodes (Adair et al., 1982), and, moreover, since important immune cell modifications occur in lymph nodes (Randolph et al., 2005).
Since lymphatic vessels are draining cells and fluid from practically all tissues drains into lymph nodes and other lymphoid tissues, lymphatics are ideally suited for immune surveillance (Swartz et al., 2008) (Fig. 1) and thus are an important element of the immune system. Soluble antigens, filtered by antigen-presenting cells residing in the lymph node sinuses and particulate antigens that are presented by dendritic cells traveling to lymph node after phagocytosis (Randolph et al., 2005, 2008), are fundamental for the initiation and propagation of immune responses. These processes are not completely understood, but seem to involve chemokines CCL 21, CCL 19, and CCL 12 released by lymphatic endothelial cells that bind to their cognate receptors CCR7 and CXCR4 on mature dendritic cells to induce mobilization, vessel entry, and trafficking (Forster et al., 1999; Randolph et al., 2008; Jackson, 2009).
Lymphatic Vessel Markers, Lymphangiogenic Factors, and Lymphangiogenesis
In contrast to lymph vessels, which were described at the beginning of the 17th century, growth factors and molecular markers for these vessels have been known for fewer than 20 years, during which lymphatic vascular biology has advanced rapidly through the discovery of lymphangiogenic factors, identification of lymphatic vascular markers, isolation of lymphatic endothelial cells, and the development of animal models for the study of lymphangiogenesis (Tammela and Alitalo, 2010; Alitalo, 2011). These new lymphatic vessel markers, which can be used to distinguish lymphatic from blood vessels, have been crucial for recent progress in the entire lymphatic biology field. Particularly important are homeobox transcription factor Prox1 (Wigle and Oliver, 1999), the lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) (Banerji et al., 1999), podoplanin (Breiteneder-Geleff et al., 1999), and VEGFR- 3 (Kaipainen et al., 1995). These markers are the most commonly used. There are, however, several others that can be used for differentiation of lymph and blood vessels (recently reviewed in, e.g., Jurisic and Detmar, 2009) and thus for isolation of LECs for further studies in vitro.
Factors that induce the growth of lymph vessels are abundant. The first to be identified was VEGF-C (Joukov et al., 1996) and then VEGF-D (Achen et al., 1998). Both bind to VEGFR-3 (flt-4) on LECs and result in downstream signaling. Later, several other lymphatic growth factors have been identified, including hepatocyte growth factor, angiopoietin-1, fibroblast growth factor-1, insulin-like growth factors 1 and 2, platelet-derived growth factors, adrenomedullin, growth hormone, and endothelin, all discussed in more detail in a recent review (Jurisic and Detmar, 2009). Importantly, however, these latter factors may have an indirect lymphangiogenic effect through their induction of VEGF-C and -D (Tammela and Alitalo, 2010). Many of the lymphangiogenic factors are also angiogenic, due to the common embryonic origins of lymphatic and blood vessels (Tammela and Alitalo, 2010). Accumulating evidence indicates a potential significance for ECM in regulating lymphangiogenesis, through integrin- or non-integrin-mediated interactions (Wiig et al., 2010).
Lymphangiogenesis, the formation of new lymphatic vessels from existing ones, takes place during development as well as in wound healing, inflammation, and cancer, occurring in inflamed tissue as well as in the respective draining lymph nodes (Tammela and Alitalo, 2010; Alitalo, 2011; Kim et al., 2012). VEGFR-3-dependent lymphangiogenesis has been found to be involved in the resolution of inflammation and edema in some models of experimental inflammation, including a mouse model of chronic airway inflammation (Baluk et al., 2005) and inflammation in mouse skin (Kajiya et al., 2009). Analysis of more recent data has supported these findings, showing that VEGF-C, -D, and -A are all important for the resolution of inflammation in mouse skin (Kataru et al., 2009; Huggenberger et al., 2010, 2011). The inhibition of lymphangiogenesis via blockade of VEGFR-3 in a mouse model of chronic inflammatory arthritis reduced lymph drainage and aggravated inflammation (Guo et al., 2009), whereas, in the same model, intra-articular delivery of VEGF-C improved lymph drainage and diminished joint tissue damage (Zhou et al., 2011). Together, these studies in mice indicate that inflammatory lymphangiogenesis plays a central role in clearing edema fluid and antigens, and moreover that local delivery of VEGF-C may be used for the treatment of certain types of inflammatory disorders.
Normal Distribution and Function of Lymphatic Vessels in Gingiva
The distribution of lymphatic vessels in gingiva in mice was first described in detail after identification with LYVE-1 as a specific marker (Ushijima et al., 2008). Immunolabeling showed lymphatic vessels in the connective tissue under the epithelium and in the junctional epithelium. The initial lymphatic vessels were adjacent to the epithelial cells, even protruding into the epithelium in the connective tissue papillae (see also Fig. 1b). In gingiva, lymphatics start as blind-ended vessels and travel from their origin along the external surface of the alveolar bone (Ushijima et al., 2008). No lymphatic vessels are detected on the border between the gingiva and the periodontal ligament (Ushijima et al., 2008). Whether there is lymphatic drainage from the periodontal ligament has not been established, since conflicting data have been published. In the rat apical periodontal ligament (Bletsa et al., 2012) and in human apical lesions (Virtej et al., 2013), no lymphatic vessels have been found, whereas in horse periodontal ligament, lymphatic vessels have been shown (Staszyk et al., 2005). It is therefore uncertain if fluid from the periodontal ligament is drained from lymphatic vessels within the tissue or if fluid is transported through the interstitium into adjacent tissue supplied with lymphatic vessels.
Maintenance of lymphatics is dependent on the constant supply of the growth factors VEGF-C and -D binding to VEGFR-3. The importance of such binding was demonstrated in K14-VEGFR-3-Ig mice overexpressing soluble VEGFR-3 (Mäkinen et al., 2001). In this murine model, genetically engineered for studies of primary lymphedema, the growth factors VEGF-C and -D are trapped, resulting in complete regression of dermal (Mäkinen et al., 2001) and gingival lymphatics (Mkonyi et al., 2010) during embryonic development. The blood vessel network remains normal in these mice, and the inhibition appears to be specific to the lymphatics (Mäkinen et al., 2001; Mkonyi et al., 2010).
In normal mice, both growth factors are expressed by gingival epithelial cells. VEGF-C is widely expressed in CD45+ cells in lamina propria and also within the epithelium, whereas VEGF-D is found only in a few immune cells in the connective tissue (Mkonyi et al., 2012a). The activity of the homeobox gene Prox1 is necessary and sufficient to maintain a lymphatic endothelial fate (Johnson et al., 2008), and Prox1 has been shown in gingival lymphatic vessels (Staszyk et al., 2005). Lymphatic endothelial cells are one of the few differentiated cell types that require the constant expression of a particular ‘instructor’ gene (in this case, Prox1) to maintain their phenotype (Oliver and Srinivasan, 2010).
Inflammation and Fluid Transport in Gingiva
In gingiva (as in most other tissues), the lymphatic vessels drain excess interstitial fluid. Moreover, crevicular fluid can be formed due to fluid transport from the tissue into the gingival sulcus area. The crevicular fluid formation increases during gingival inflammation as a result of increased interstitial fluid pressure (Del Fabbro et al., 2001). Several investigators have measured the Pif in gingiva and found a range between -1.0 and +3.5 mm Hg in free gingival and 0.0 and +9.6 mm Hg in attached gingiva under normal conditions (Fjaertoft et al., 1992; Aarli and Heyeraas, 1995; Del Fabbro et al., 2001; Mkonyi et al., 2010). A Pif in gingiva above atmospheric pressure will also tend to promote fluid transport into the sulcus area and contribute to a low, but constant, production of crevicular fluid. The rise in Pif during inflammation varies from around 100% (free gingiva) to 50% (attached gingiva) (Fjaertoft et al., 1992; Mkonyi et al., 2010). The colloid osmotic pressure (COPif, Fig. 2) in gingiva, which is another factor controlling lymph flow in the tissue, has been found to range between 9.6 and 14.1 mm Hg (Aarli and Heyeraas, 1995; Del Fabbro et al., 2001) and to decrease during inflammation because of dilution after increased filtration (Aarli and Heyeraas, 1995).
In the K14-VEGFR-3-Ig mouse model lacking lymphatic vessels in gingiva (see above), Mkonyi et al. (2012b) demonstrated that Pif was elevated in gingiva devoid of lymphatics by approximately 50%. Acute inflammation increased the interstitial fluid pressure to a significantly higher level in K14-VEGFR-3-Ig compared with normal mice, due to more fluid accumulation in the former. Together, these results demonstrate that gingival lymphatics play an important role in tissue fluid homeostasis in gingiva in both the normal and inflamed conditions. In mice lacking gingival lymphatics, more infiltration of CD45+ cells was found in the tissue (Mkonyi et al., 2012b), suggesting that lymphatic vessels normally attenuate an inflammatory process by their draining capacity.
Del Fabbro et al. (2001) further showed that, during inflammation, proteins accumulated in the crevicular fluid derived from inflamed gingiva and/or from dental plaque bacteria, creating a pressure gradient driving fluid from the gingiva interstitium into the sulcus area. Increased capillary pressure (induced by venous stasis) in gingiva is followed by about 100% increase in both Pif and crevicular fluid formation (Aarli and Heyeraas, 1995). It is likely, moreover, that gingival lymph flow increases in these situations, due to the increased Pif representing the filling pressure for initial lymphatics (Wiig and Swartz, 2012).
Lymphangiogenesis in Periodontal Disease
As mentioned above, lymphangiogenesis takes place during inflammation to enhance fluid and immune cell transport from the periphery toward lymph nodes. In gingiva, lymphangiogenesis was recently demonstrated in mice infected with P. gingivalis; a model for murine periodontal disease (Mkonyi et al., 2012a). The increased growth of vessels should ideally contribute in clearing the infection and resolving the inflammation, but lymphangiogenesis has also been shown to actively participate in maintenance of chronic inflammatory diseases (Cueni and Detmar, 2006). As stated previously, the most important growth factors for lymphangiogenesis are VEGF-C and VEGF-D. After induction of gingival inflammation by P. gingivalis, the number of VEGF-C+/CD45+ immune cells increased significantly in gingiva, whereas the number of VEGF-D+/CD45+ cells was low and unchanged compared with control levels (Mkonyi et al., 2012a). Inflammatory cytokines produced by recruited immune cells are known stimulators of VEGF-C and -D production, and in the inflamed gingiva an up-regulation of TNF-α and an insignificant increase in IL-1β were found 10 days post-infection (Mkonyi et al., 2012a). Both IL-1 β and TNF-α are strong promoters of lymphatic vessel growth, as demonstrated after airway infection (Baluk et al., 2009, 2013).
In the same study (Mkonyi et al., 2012a), the lymphatic responses included growth of lymphatic vessel area and numbers of vessel profiles in gingiva (Fig. 3). In addition, proliferating lymphatic endothelial cells were detected in lamina propria 10 days post-infection. In contrast to VEGF-C, changes in VEGF-D protein expression were not observed after infection, and the gene expression was significantly lowered, probably due to a toxic effect of the bacteria upon the epithelial cells, which were immunopositive to VEGF-D. Taken together, it seems as if VEGF-C is mainly responsible for the inflammatory lymphangiogenesis that takes place after onset of a periodontal infection. New evidence on the influence of VEGF-C is also provided in a transgenic mouse model, which shows overexpression of VEGF-C produced by the epithelial cells in gingiva. In this model, a significant increase in lymphatic area (hyperplasia) is found and further supports the important role of VEGF-C for lymphatic growth in gingiva (Fig. 4, unpublished observations, Papadakou et al., 2013).
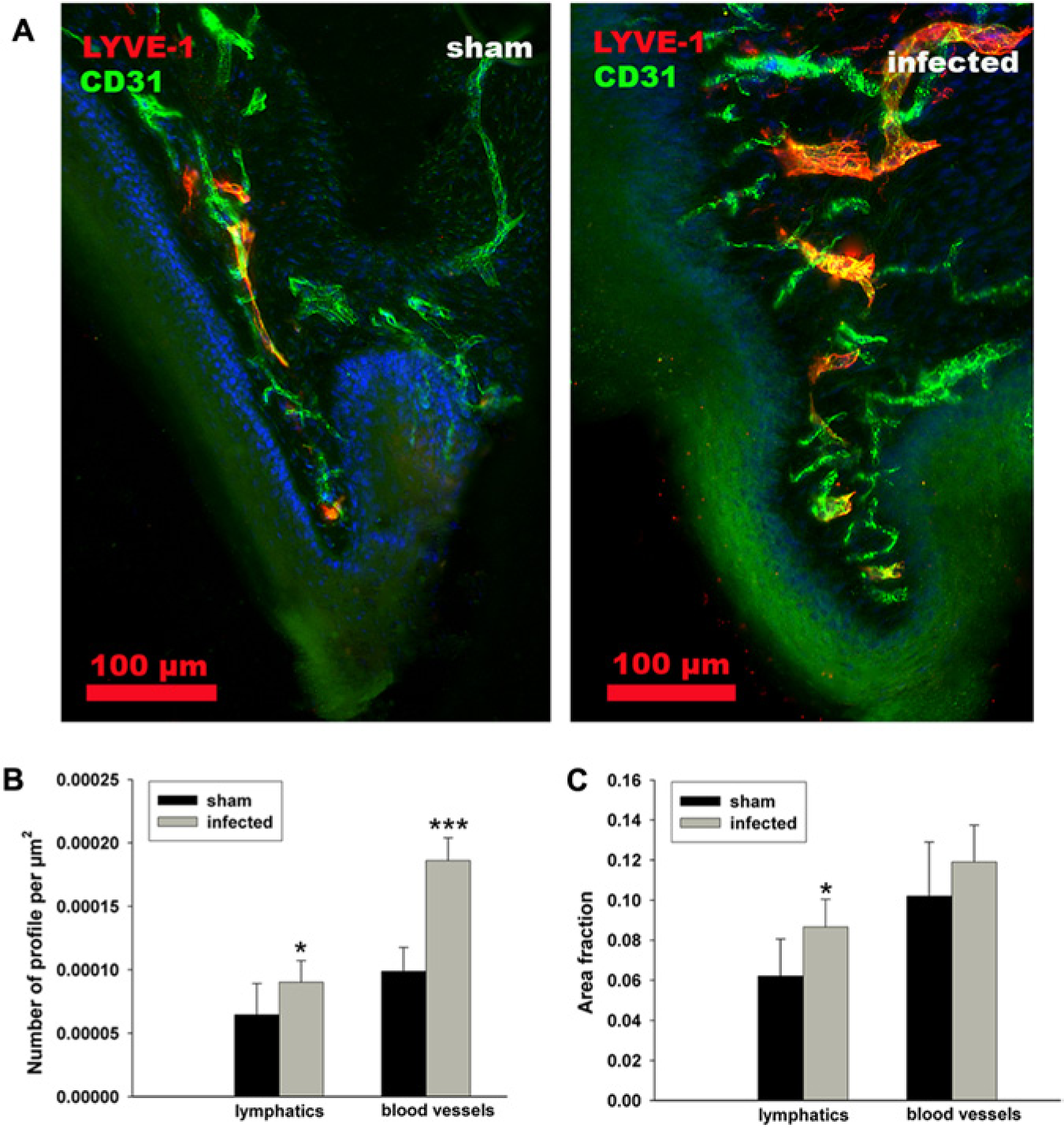
A gingival papillae in a normal situation (left) and after induction of inflammation (right) promoted by P. gingivalis infection 10 days previously
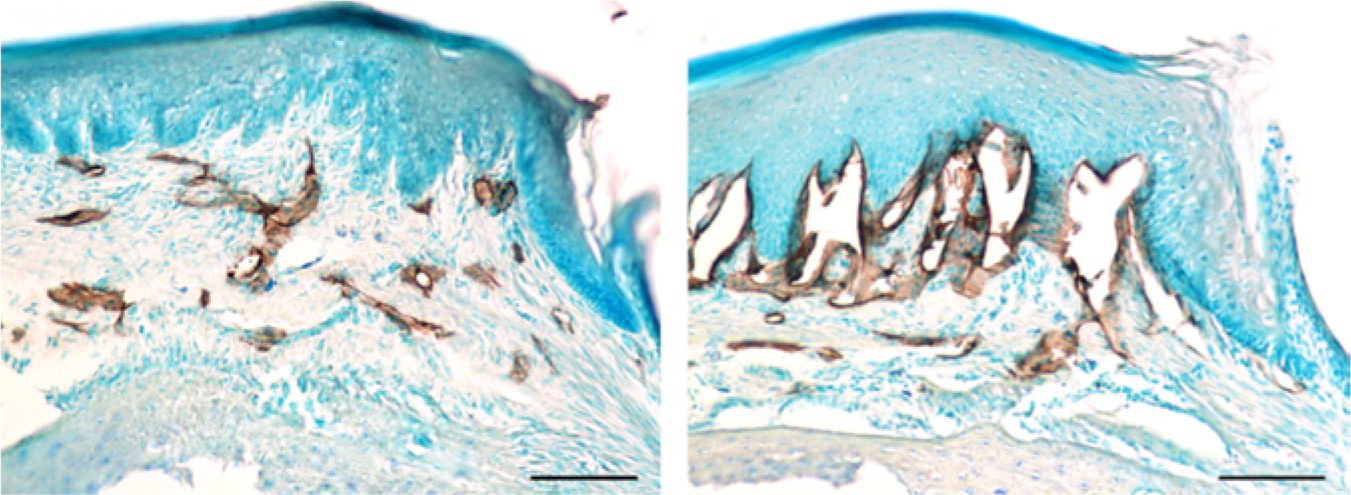
Lymphatic vessels in the palatal gingiva and mucosa immunolabeled with anti-LYVE-1 (brown), in a normal WT mouse
Importance of Lymphatics in Periodontal Disease
We recently showed a clearly protective role of gingival lymphatic vessels in periodontal disease development (Mkonyi et al., 2012b). Using the K14-VEGFR-3-Ig mouse model discussed above, we showed that these mice without lymphatic vessels in gingiva developed more severe periodontal disease with bone resorption than their WT littermates after challenge with the known periodontal pathogen P. gingivalis. Histological sections revealed a strong local inflammatory response, including microabscesses in the gingiva. Immunohistochemistry demonstrated high numbers of F4/80+ macrophages in bone resorption lacunae (Fig. 5A, K14-infected), and a higher number of F4/80+ cells and antigen-presenting MHC class II+ cells toward the alveolar bone in the transgenic mice (Figs. 5B, 5C). Neutrophils were surrounding microabscesses, and an enhanced inflammatory process in transgenic mice was further verified by cytokine analyses. Pro-inflammatory cytokines such as IFN-γ and IL-1β were significantly more numerous in the mutant mice. The latter stimulate RANKL and M-CSF, which together cause mature osteoclast activation. The impaired lymphatic drainage from gingiva of bacteria and bacterial products probably promotes a more severe local inflammatory response in the transgenic mice. The impaired drainage and thus reduced ability for antigen-presenting cells to travel in lymphatic vessels toward the lymph nodes is likely the explanation for the hampered humoral immune response measured as specific IgG to P. gingivalis (Mkonyi et al., 2012b). Depressed transport of solutes and dendritic cells from the skin to draining lymph nodes and lower antibody production have also been demonstrated in the skin in the same mouse model (Thomas et al., 2012).
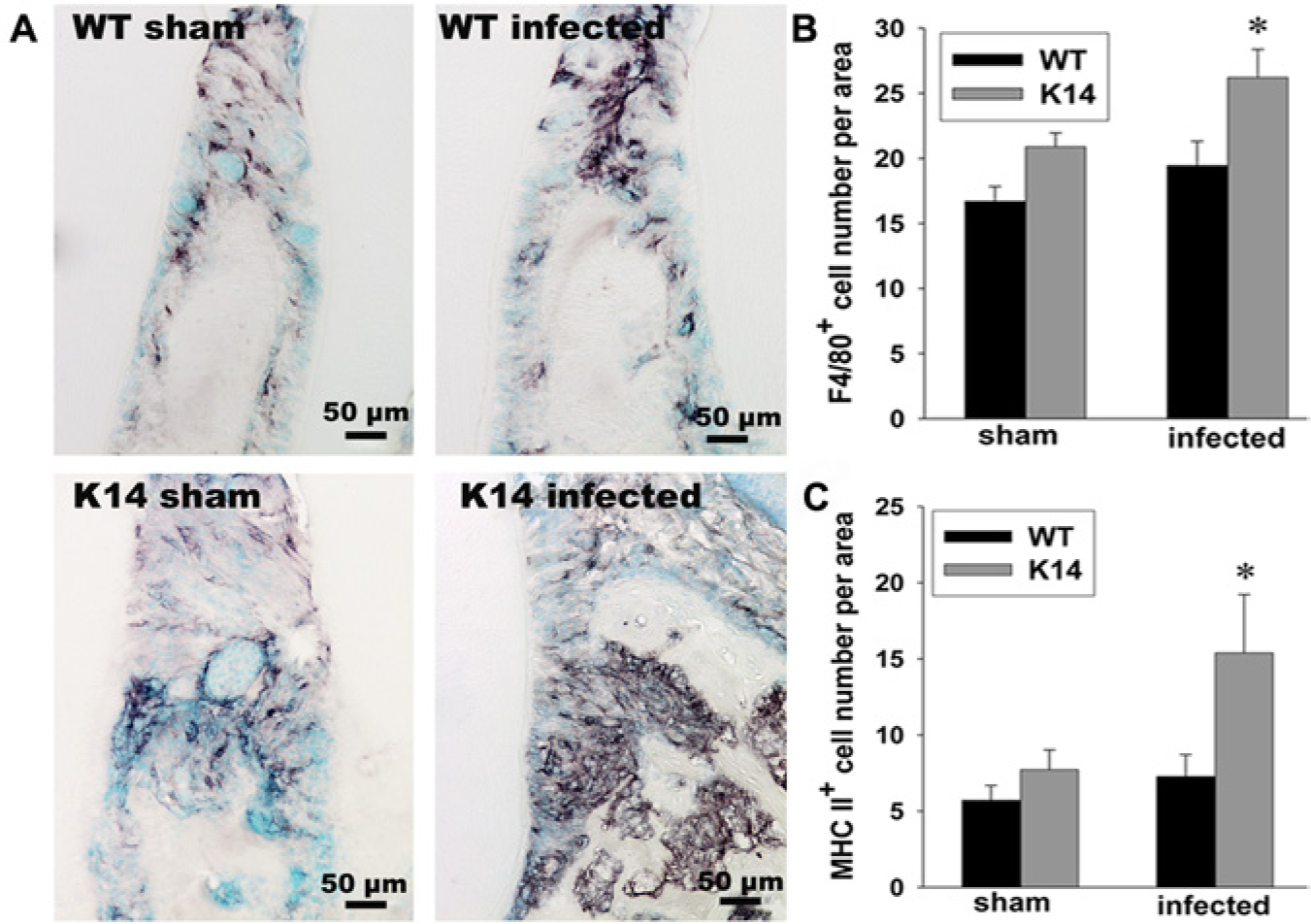
In a transgenic strain (K14-VEGF receptor 3-Ig) of mice lacking lymphatic vessels in gingiva, a severe infiltration of F4/80+ macrophages was found close to areas of bone resorption after P. gingivalis challenge
Clinical Implications
It is still unknown if lymphangiogenesis contributes to protecting patients with inflamed gingiva against bone loss. The lymphatic response in general is known to be long-lasting, even after an infection is cleared (Baluk et al., 2005), and a remaining expanded vessel area may contribute to an enhanced adaptive immune response if a new infection period occurs. Few studies have addressed the functional role of lymphatics in chronic infections, but lessons learned from other studies show that correction of defective lymphatic function may benefit the treatment of other inflammatory diseases in mucosal tissues (Baluk et al., 2005). Whether or not an enhanced immune response is favorable for periodontal disease development remains to be investigated. Little is known about lymphatic function in patients with periodontitis.
In a study investigating normal and inflamed gingiva from patients with periodontitis, the protein expression of CCL21 was detected in healthy gingiva only (Kuroshima et al., 2004). CCL21 is the primary determinant for dendritic cell entry into peripheral lymphatic vessels through engagement of their receptor CCR7. Lymphatic endothelial cells express the CCL21 on the abluminal surface (Vassileva et al., 1999; Tal et al., 2011), and a single CCL21 isoform exists in humans (Randolph et al., 2005). The authors speculated that increased proteolytic enzymatic activity in inflamed gingiva may cleave CCL21 into inactive fragments and contribute to impaired antigen presentation. Enzymatic activity is also responsible for degradation of the connective tissue due to production of metalloproteinases and collagenases in periodontally diseased tissue (Reynolds, 1996; Nomura et al., 1998). It is likely that tissue degradation is followed by impaired lymphatic function, due to the fact that lymphatic vessels are attached to the interstitium via anchoring filaments and open up in edema when the interstitial fluid volume and tissue pressure increase.
Concluding Remarks
It is tempting to speculate that impaired lymphatic function may exaggerate the progress of periodontal disease in patients, and this hypothesis remains to be tested. Investigations of the lymphatic endothelial cell responses to the local environment in gingiva during inflammation may also give new evidence on the pathogenesis of periodontal disease. Translational research in this relatively new field will be important for a more complete understanding of periodontal responses to inflammatory insults.
Footnotes
Acknowledgements
We thank Hanne Haslene-Hox for help with figure preparation.
The author(s) received no financial support and declare no potential conflicts of interest with respect to the authorship and/or publication of this article.