
Abstract
Porphyromonas endodontalis and its main virulence factor, lipopolysaccharide (LPS), are associated with the development of periapical diseases and alveolar bone loss. Calcium hydroxide is commonly used for endodontic therapy. However, the effects of calcium hydroxide on the virulence of P. endodontalis LPS and the mechanism of P. endodontalis LPS–induced bone destruction are not clear. Calcium hydroxide rescued the P. endodontalis LPS–suppressed viability of MC3T3-E1 cells and activity of nuclear factor-κB (NF-κB) in these cells, resulting in the reduced expression of interleukin-6 and tumor necrosis factor-α. In addition, calcium hydroxide inhibited P. endodontalis LPS–induced osteoclastogenesis by decreasing the activities of NF-κB, p38, and ERK1/2 and the expression of nuclear factor of activated T-cell cytoplasmic 1 in RAW264.7 cells. Calcium hydroxide also rescued the P. endodontalis LPS–induced osteoclastogenesis and bone destruction in mouse calvaria. Taken together, our present results indicate that calcium hydroxide suppressed bone destruction by attenuating the virulence of P. endodontalis LPS on bone cells.
Keywords
Introduction
Gram-negative anaerobic micro-organisms play crucial roles in endodontic-periodontal infections (Wittgow and Sabiston, 1975; Assed et al., 1996). Among them, Porphyromonas endodontalis is the most commonly isolated species from root canal and apical lesions (Tomazinho and Avila-Campos, 2007; Montagner et al., 2012). A major virulence factor of P. endodontalis is lipopolysaccharide (LPS), which plays critical roles in inducing the secretion of pro-inflammatory cytokines and bone destruction (Hong et al., 2004; Yang et al., 2009; Tang et al., 2011). Calcium hydroxide has long been used as a root canal disinfectant. Because of its low solubility, calcium hydroxide releases calcium and hydroxyl ions slowly, thus providing an alkaline environment in the periapical region (Siqueira and Lopes, 1999; Farhad and Mohammadi, 2005). Previous studies indicate that calcium hydroxide disinfects the infected root canal, dissolves necrotic tissue, prevents absorption of teeth, and promotes hard-tissue formation (Khan et al., 2008). However, there has been little information about the mechanisms of calcium hydroxide in P. endodontalis LPS–induced bone destruction.
Bone remodeling is controlled by the cooperation between osteoblasts and osteoclasts to maintain normal bone volume and calcium homeostasis (Kukita et al., 2012). In periapical diseases, the balance between osteoblasts and osteoclasts is disrupted, leading to bone destruction. LPS liberated from micro-organisms acts as a potent stimulator of bone destruction. LPS has been reported to inhibit osteoblast differentiation and bone mineralization (Roberts et al., 2008; Bonsignore et al., 2013). LPS also induces many types of inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukins (ILs), and prostaglandin E2 in osteoblasts (Matsushita et al., 1999; Gao et al., 2013). These factors are reported to participate in alveolar bone resorption and the inhibition of bone formation (Lam et al., 2000; Inada et al., 2006; Rogers et al., 2007). Conversely, LPS promotes survival of pre-osteoclasts and induces their fusion to form active multinucleated osteoclasts (Suda et al., 2002).
Nuclear factor κB (NF-κB) is activated and translocated to the nucleus in response to LPS in many types of cells. NF-κB is a crucial factor which regulates a variety of inflammatory mediators (Baldwin, 2001). Mitogen-activated protein kinases (MAPKs) also play important roles in the expression of nuclear factor of activated T-cell cytoplasmic 1 (NFATc1) during osteoclastogenesis (Takayanagi et al., 2002). It was reported that P. endodontalis LPS activates NF-κB in post-natal dental pulp stem cells and stimulates osteoclastogenesis (Chang et al., 2005; Tang et al., 2011).
The purpose of this study was to examine the effects of calcium hydroxide on P. endodontalis–induced cytokine expression in osteoblasts. We also investigated the effects of calcium hydroxide on P. endodontalis–induced bone destruction and osteoclastogenesis in vivo.
Materials & Methods
Bacterial Culture and LPS Extraction
P. endodontalis (ATCC35406) was obtained from the Central Laboratory of Capital Medical University (Beijing, China) and cultured anaerobically at 37°C. LPS was extracted by the hot phenol-water method as previously described (Koga et al., 1985; Roberts et al., 2008). The bioactivity of purified P. endodontalis LPS was measured with the Limulus Amoebocyte Lysate (LAL) endotoxin assay kit (GenScript, Piscataway, NJ, USA).
Preparation of Calcium Hydroxide Solution
A 200-mg quantity of calcium hydroxide (Wako, Osaka, Japan) powder was dissolved in 50 mL distilled water, shaken vigorously, and centrifuged at 1,000 × g for 10 min. The supernatant was filtered and retained as the saturated solution (100% volume fraction). Calcium hydroxide (15%) and 10 µg/mL of P. endodontalis LPS were mixed and incubated for 6 hr at 37°C. The mixture (CH-LPS) was stored at −80°C prior to use.
Cell Viability Assay
MC3T3-E1 cells obtained from Riken (Tsukuba, Japan) were cultured in α-MEM supplemented with 10% FBS at 37°C in a humidified atmosphere with 5% CO2. They were seeded into 96-well plates at a density of 0.2 × 104 cells/well. Attached cells were treated with calcium hydroxide, P. endodontalis LPS, or CH-LPS for 5 days. Cell viability was measured with the MTT assay kit (Sigma-Aldrich, St. Louis, MO, USA).
RNA Isolation and RT-PCR Analysis
Total cellular RNA was extracted with the Trizol reagent (Invitrogen, Carlsbad, CA, USA). The cDNA synthesis reaction was carried out with the Reverse Transcription Kit (Takara, Kyoto, Japan). The primer sequences were as follows: IL-6, forward 5′-GGTGACAACCACGGCCTTCCC-3′, reverse 5′-G CCACTCCTTCTGTGACTCCAGC-3′; TNF-α, forward 5′-AC GGCATGGATCTCAAAGAC-3′, reverse 5′-CGGACTCCGC AAAGTCTAAG-3′; and β-actin, forward 5′-CAGAGCAA GAGAGGCATCC-3′, reverse 5′-CTGGGGTGTTGAAGGT CTC-3′. PCR products were analyzed by separation on 1.5% agarose gels containing 400 ng/mL ethidium bromide.
Immunofluorescence
MC3T3-E1 cells were treated for 30 min with P. endodontalis LPS or CH-LPS. Cells were then fixed with 3.7% formaldehyde and incubated with an anti-NF-κB antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C overnight. After incubation with Alexa Fluor 488 goat IgG (Invitrogen) for 40 min and 10 µg/mL Hoechst 33342 for 20 min, the cells were mounted with fluorescent mounting medium.
Luciferase Assay
NF-κB luciferase reporter was obtained from Stratagene (La Jolla, CA, USA). The cells were transfected with NF-κB reporter vector with the aid of Lipofectamine LTXTM reagent (Invitrogen). GL3-basic vector (Promega, Madison, WI, USA) was used as the empty-vector control. The cells were then treated with P. endodontalis LPS or CH-LPS for 1 hr. The efficiency of transfection was standardized by co-transfection with pTK-Renilla (Promega). Total cell lysates were prepared with the Dual-Glo® Luciferase Assay System (Promega) and assessed for luciferase activity.
Western Blotting
Cells were scraped into lysate buffer. Proteins (15 µg) were probed with specific antibodies (1:1,000) followed by incubation with secondary antibodies (1:5,000). Interacting proteins were visualized with an ECL detection kit (Amersham Pharmacia Biotech, Uppsala, Sweden) according to the manufacturer’s protocol.
Tartrate-resistant Acid Phosphatase (TRAP) Staining
Cells and dissected mouse calvaria were fixed in acetone-citrate-formaldehyde for 15 min at ambient temperature. TRAP staining was performed with 0.01% naphthol AS-MX phosphate and 0.005% fast red violet LB salt in the presence of 50 mM sodium tartrate and 90 mM sodium acetate (pH 5.0) for 15 min at 37°C and rinsed twice with distilled water.
Micro-computed Tomography (µCT) and Histological Analyses
All animal experiments were conducted under the regulation and permission of the Animal Care and Use Committee of the University of Tokushima, Tokushima, Japan (toku-dobutsu 10051). Three-day-old Balb/c mice were injected daily in the calvarial region with 50 µL of P. endodontalis LPS (10 µg/mL), 15% calcium hydroxide, CH-LPS, or ultrapure water. After 12 days, mice were sacrificed, and calvaria were dissected. For computed tomography (CT)-based analysis of bone mineral density (BMD), the central region of the extracted calvaria was scanned at 0.2-mm intervals in an experimental animal µCT system (Aloka, Tokyo, Japan). BMD was calculated with LaTheta software (version 3.22) equipped with the CT system. Based on the data obtained from µCT scanning, 3-dimensional images of the calvaria were constructed with Avizo 6.1 software (Burlington, MA, USA). After being used for µCT analysis, dissected mice calvaria were embedded in paraffin and sectioned. Trichrome staining was performed with the Masson-Goldner staining kit (Merck, Darmstadt, Germany).
Statistical Analysis
All data are presented as mean ± standard error of the mean (SEM), and at least 3 independent experiments were performed for each assay. Statistical analyses were conducted by analysis of variance (ANOVA). Statistical significance is indicated with *p < .05 and **p < .01.
Results
Calcium Hydroxide Abrogated the P. endodontalis LPS–induced Reduction of Osteoblast Viability
MC3T3-E1 cells were treated with various concentrations of calcium hydroxide or P. endodontalis LPS for 5 days, and cell viability was measured by the MTT assay. Compared with the untreated cells, lower concentrations (less than 15%) of calcium hydroxide increased cell viability, whereas cell viability was decreased in the cells treated with higher concentrations (greater than 20%) of calcium hydroxide (Fig. 1A). P. endodontalis LPS decreased osteoblast viability in a dose-dependent manner (Fig. 1B). P. endodontalis LPS (10 µg/mL) was mixed and incubated for 6 hr at 37°C with 15% calcium hydroxide (CH-LPS). As shown in Fig. 1C, CH-LPS increased osteoblast viability, indicating that calcium hydroxide counteracted the negative effect of P. endodontalis LPS on osteoblast viability.
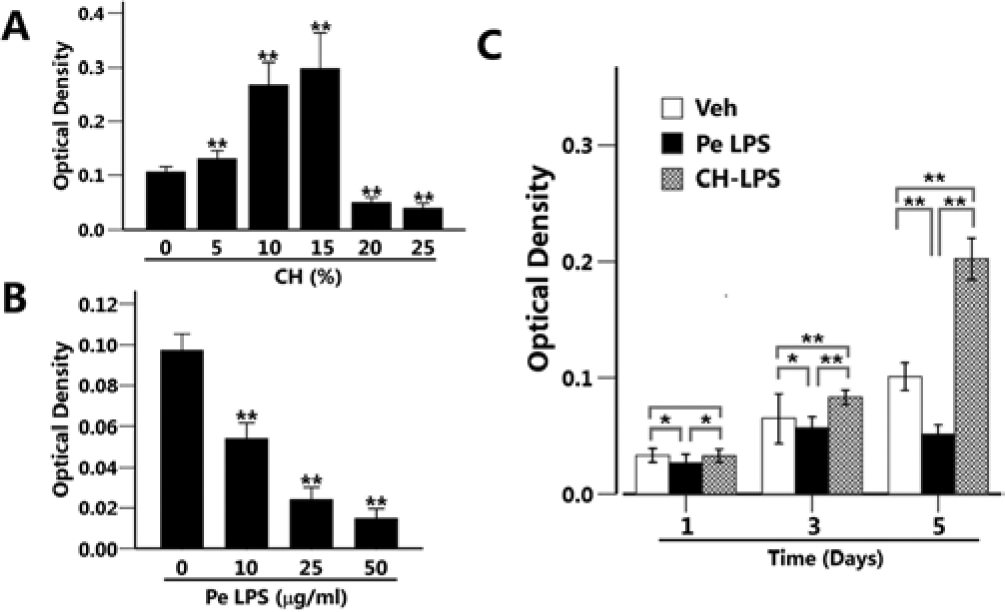
Calcium hydroxide abrogated the P. endodontalis lipopolysaccharide (LPS)–induced reduction of osteoblast viability.
Calcium Hydroxide Inhibited P. endodontalis LPS–induced IL-6 and TNF-α Expression and the Activation of NF-κB in Osteoblasts
Fig. 2A shows that P. endodontalis LPS induced expression of IL-6 and TNF-α in MC3T3-E1 cells. Calcium hydroxide alone did not affect the expression of these cytokines. P. endodontalis LPS was pre-incubated with calcium hydroxide for the indicated times and subsequently used for the treatment of MC3T3-E1 cells. Calcium hydroxide decreased the P. endodontalis LPS–induced expression of IL-6 and TNF-α (Fig. 2A). The expression of β-actin was used as an internal control and did not change in each group.
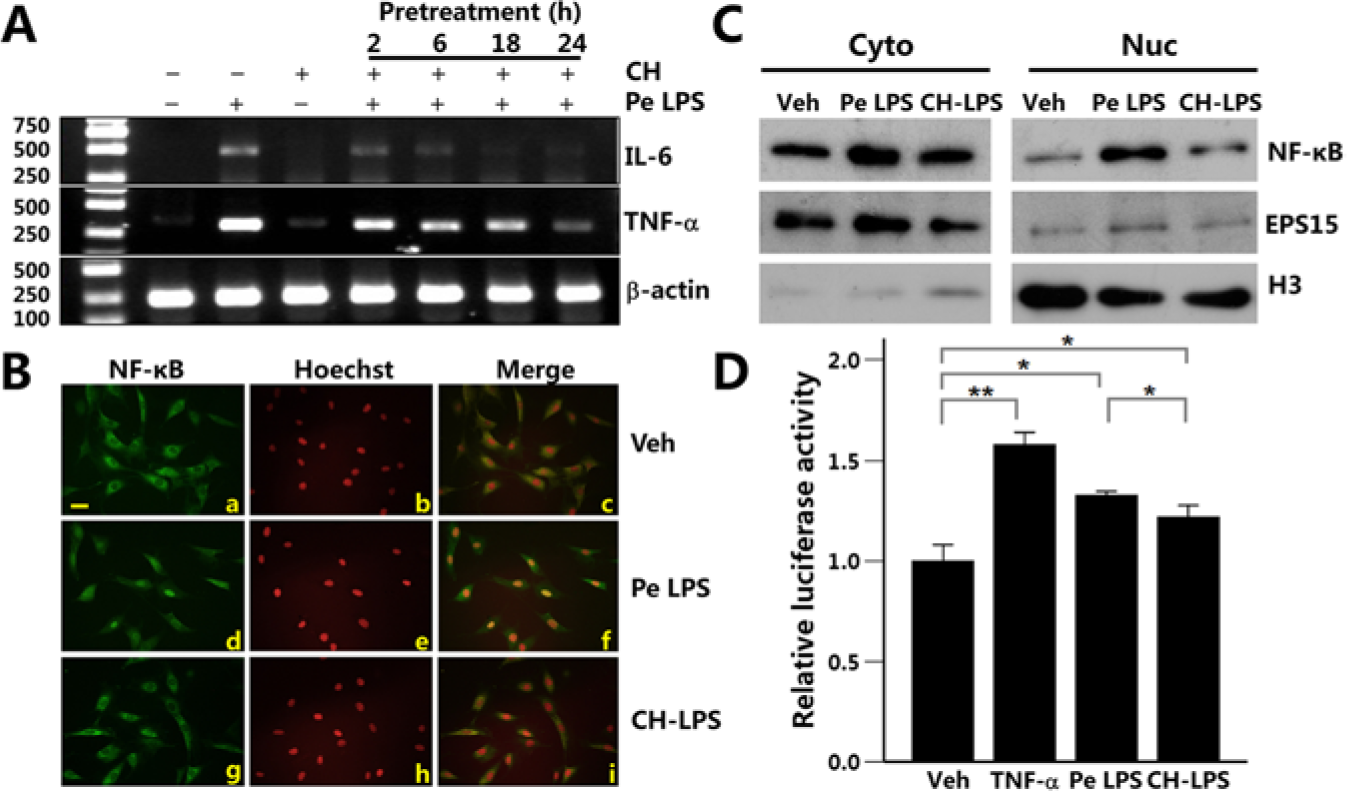
Calcium hydroxide inhibited P. endodontalis LPS–induced IL-6 and TNF-α expression and the NF-κB activation in osteoblasts.
To examine whether NF-κB activation participated in this course, we treated MC3T3-E1 cells with P. endodontalis LPS or CH-LPS for 30 min and analyzed the activation and nuclear translocation of NF-κB by immunostaining and Western blot. In the untreated cells, NF-κB localized in the cytoplasm (Fig. 2Ba). Nuclear translocation of NF-κB was observed in the cells treated with P. endodontalis LPS (Fig. 2Bd). In contrast, NF-κB localized in the cytoplasm in the cells treated with CH-LPS (Fig. 2Bg). The localization of NF-κB was further confirmed by cell fractionation. Increased expression of NF-κB was detected in the nuclear fraction of P. endodontalis LPS-treated cells but not in the untreated or CH-LPS–treated cells (Fig. 2C). The purity of the cytoplasmic and nuclear fractions was confirmed by the expression of Eps15 and histone H3, respectively. Results from the luciferase assay also indicated that P. endodontalis LPS increased NF-κB transcriptional activity. CH-LPS partially suppressed the P. endodontalis LPS–induced NF-κB transcriptional activity (Fig. 2D). TNF-α treatment was used as the positive control.
Calcium Hydroxide Suppressed P. endodontalis LPS–induced Osteoclastogenesis
P. endodontalis LPS dramatically increased the number of TRAP-positive multinuclear RAW264.7 cells. However, CH-LPS decreased P. endodontalis LPS–induced osteoclastogenesis (Figs. 3A, 3B). As shown in Fig. 3C, the maximum expression of NFATc1 was observed at 24 hr after stimulation with P. endodontalis LPS. However, CH-LPS suppressed P. endodontalis LPS–induced NFATc1 expression. P. endodontalis LPS increased the transcriptional activity of NF-κB in RAW264.7 cells, which was decreased by the CH-LPS treatment (Fig. 3D). Similarly, the levels of phosphorylated-p38 (p-p38) and phosphorylated-ERK (p-ERK) were increased in the cells treated with P. endodontalis LPS. Little expression of p-p38 and p-ERK was observed after the stimulation with CH-LPS (Fig. 3E).
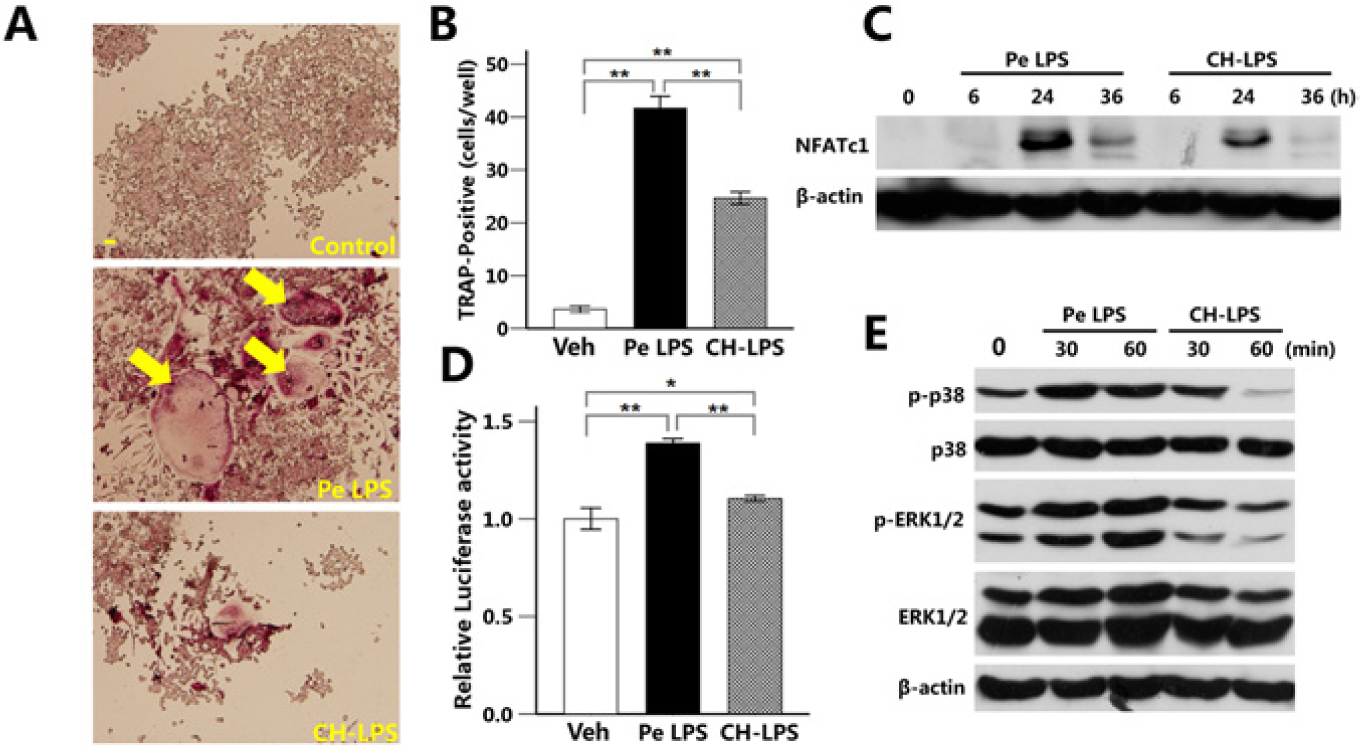
Calcium hydroxide suppressed P. endodontalis LPS–induced osteoclastogenesis.
Calcium Hydroxide Rescued P. endodontalis LPS–reduced Bone Formation
P. endodontalis LPS and CH-LPS were daily injected into mouse calvaria. TRAP staining showed that administration of P. endodontalis LPS increased osteoclastogenesis in mouse calvaria, while CH-LPS decreased the intensity of TRAP staining. Calcium hydroxide alone did not affect osteoclastogenesis (Fig. 4A). P. endodontalis LPS induced bone destruction, as demonstrated by the 3-dimensional µCT image. CH-LPS rescued the P. endodontalis LPS–suppressed bone formation (Fig. 4B). CH-LPS recovered P. endodontalis LPS–reduced bone mineral density (Fig. 4C). Calcium hydroxide alone increased bone mineral density (Fig. 4C). Fig. 4D shows that the thickness of calvaria was decreased by treatment with P. endodontalis LPS and that CH-LPS recovered the P. endodontalis LPS–reduced calvarial thickness.
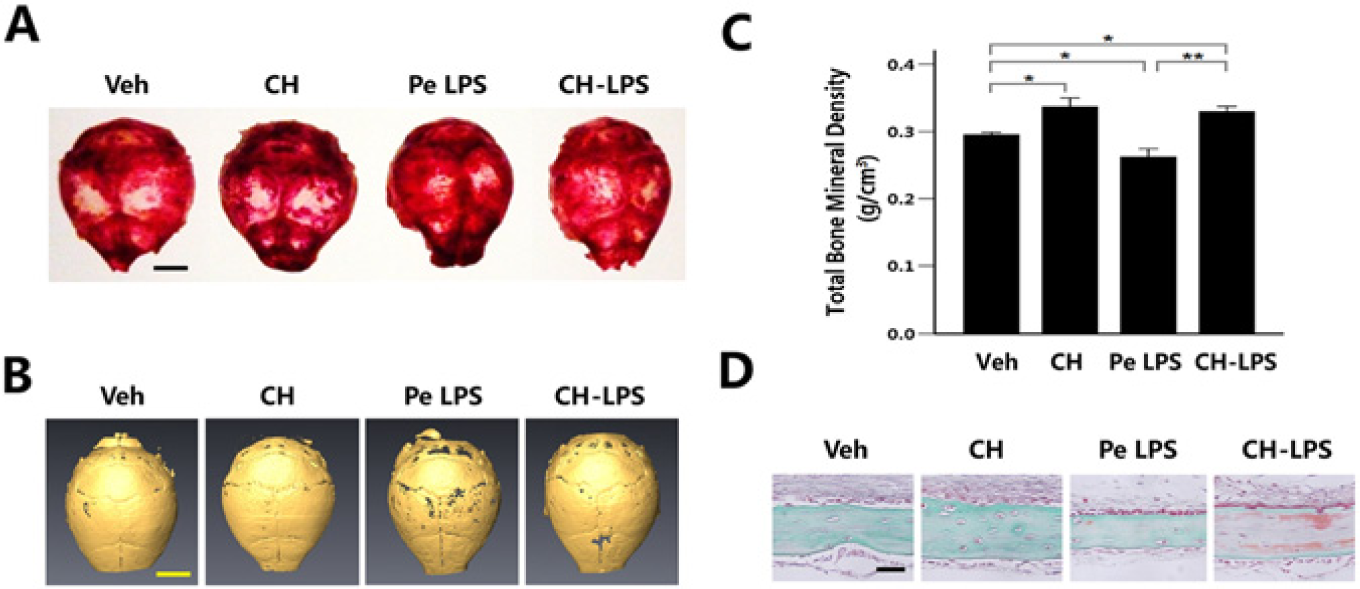
Calcium hydroxide inhibited the bone destruction caused by P. endodontalis LPS in mouse calvaria.
Discussion
In the present study, we demonstrate that calcium hydroxide (up to 15%) promoted cell viability in MC3T3-E1 cells. P. endodontalis LPS at 10 µg/mL decreased cell viability in MC3T3-E1 cells. Calcium hydroxide rescued the P. endodontalis LPS–suppressed cell viability in these cells. Calcium hydroxide is known to induce an alkaline environment and produce hydroxyl ions in the periapical region (Farhad and Mohammadi, 2005). LPS includes lipid A moiety, which is responsible for its activity, and is comprised of several ester bonds (Safavi and Nichols, 1993; Buck et al., 2001; Henderson et al., 2013). The ester bonds can be hydrolyzed by the hydroxyl ions produced from calcium hydroxide, causing the destruction of lipid A.
P. endodontalis LPS increased the expression of IL-6 and TNF-α in MC3T3-E1 cells. However, pre-incubation of P. endodontalis LPS with calcium hydroxide suppressed P. endodontalis LPS–induced cytokine expression. NF-κB is a transcriptional activator that regulates the expression of various genes controlling inflammatory responses. P. endodontalis LPS induced nuclear translocation and activation of NF-κB within 30 min after treatment, an effect that occurred prior to up-regulation of IL-6 and TNF-α. Pre-incubation with calcium hydroxide partly suppressed the nuclear translocation and activation of NF-κB induced by P. endodontalis LPS. These findings suggest that NF-κB is involved in the P. endodontalis LPS–induced IL-6 and TNF-α expression in osteoblasts. IL-6 and TNF-α are thought to contribute to bone resorption in inflammatory bone diseases. Several studies showed that TNF-α directly stimulates macrophages to differentiate into osteoclasts (Kobayashi et al., 2000; Quinn et al., 2000). IL-6 is also a powerful stimulator of osteoclastogenesis, and the blockade of the IL-6 receptor directly affects the osteoclast formation (Axmann et al., 2009). These findings and our present results suggest that calcium hydroxide regulates osteoclastogenesis by controlling cytokine production from osteoblasts.
Treatment of RAW264.7 cells with P. endodontalis LPS induced the formation of TRAP-positive multinuclear cells, which was accompanied by the activation of NF-κB, p38, and ERK and the expression of NFATc1. By contrast, pre-incubation with calcium hydroxide suppressed P. endodontalis LPS–induced osteoclastogenesis through inhibiting the activation of NF-κB, p38, and ERK and the expression of NFATc1. These results indicate that a modified structure of P. endodontalis LPS caused by calcium hydroxide abolishes its ability to activate the NF-κB, p38, and ERK signaling pathways, resulting in the reduced expression of NFATc1 and osteoclastogenesis. These findings are consistent with those in a previous report that the detoxification of Escherichia coli LPS by calcium hydroxide reduced osteoclast differentiation (Jiang et al., 2003).
We also examined the effects of calcium hydroxide on P. endodontalis LPS–induced bone destruction in vivo. To do so, we injected P. endodontalis LPS, calcium hydroxide, or CH-LPS into the mouse calvaria. CH-LPS decreased the intensity of TRAP staining of whole calvaria as compared with that of the P. endodontalis LPS–injected mice. Moreover, µCT analysis and histological examination revealed an increase in the BMD and thickness of calvaria in the CH-LPS–treated group as compared with that of the P. endodontalis LPS–treated group. Interestingly, BMD in the calcium hydroxide and CH-LPS group was slightly higher than that in the control group. It was reported that although hydroxyl ions did not affect bone mineralization, the calcium ions dissociated from calcium hydroxide were critical for inducing the mineralization of osteoblasts (Narita et al., 2010). Calcium hydroxide increased osteoblast viability in the present study. Therefore, we speculate that hydroxyl ions hydrolyze the lipid A of P. endodontalis LPS, which prevent osteoclastogenesis. The dissociated calcium ions should promote osteoblast viability and bone mineralization. Further studies are needed to clarify these speculations.
In the present study, we demonstrated that calcium hydroxide rescued the P. endodontalis LPS–inhibited viability of MC3T3-E1 cells. Calcium hydroxide also attenuated the P. endodontalis LPS–induced expression of IL-6 and TNF-α through the inhibition of NF-κB activity in these cells. In addition, calcium hydroxide inhibited P. endodontalis LPS–induced osteoclastogenesis in RAW264.7 cells. Calcium hydroxide also rescued the P. endodontalis LPS–induced bone destruction in mouse calvaria. Taken together, these findings demonstrate that calcium hydroxide suppressed bone destruction by attenuating the virulence of P. endodontalis LPS to osteoblasts and osteoclasts.
Footnotes
Acknowledgements
We thank Ms. E. Sasaki for her skillful technical assistance.
This study was supported by grants from the
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.