
Abstract
Degenerative changes of condylar subchondral bone occur frequently in temporomandibular disorders. Although psychologic stresses and occlusal abnormalities have been implicated in temporomandibular disorder, it is not known if these risks represent synergistic comorbid factors that are involved in condylar subchondral bone degradation that is regulated by the sympathetic nervous system. In the present study, chronic immobilization stress (CIS), chemical sympathectomy, and unilateral anterior crossbite (UAC) were sequentially applied in a murine model. Norepinephrine contents in the subjects’ serum and condylar subchondral bone were detected by ELISA; bone and cartilage remodeling parameters and related gene expression in the subchondral bone were examined. Subchondral bone loss and increased subchondral bone norepinephrine level were observed in the CIS and UAC groups. These groups exhibited decreased bone mineral density, volume fraction, and bone formation rate; decreased expressions of osterix, collagen I, and osteocalcin; but increased trabecular separation, osteoclast number and surface, and RANKL expression. Combined CIS + UAC produced more severe subchondral bone loss, higher bone norepinephrine level, and decreased chondrocyte density and cartilage thickness when compared to CIS or UAC alone. Sympathectomy simultaneously prevented subchondral bone loss and decreased bone norepinephrine level in all experimental subgroups when compared to the vehicle-treated counterparts. Norepinephrine also decreased mRNA expression of osterix, collagen I, and osteocalcin by mesenchymal stem cells at 7 and 14 d of stimulation and increased the expression of RANKL and RANKL/OPG ratio by mesenchymal stem cells at 2 h. In conclusion, CIS and UAC synergistically promote condylar subchondral bone loss and cartilage degradation; such processes are partially regulated by norepinephrine within subchondral bone.
Keywords
Introduction
Ten million Americans and 1% of the southern Chinese population suffer from temporomandibular disorders (TMDs) per annum (Slavkin 1996; Pow et al. 2001). Degenerative changes in the condylar subchondral bone occur frequently in symptomatic TMD patients (Cortés et al. 2011; Moncada et al. 2014). Psychological stresses (Slade et al. 2007) and occlusal abnormalities (Xie et al. 2013) have both been implicated in the development of TMD. However, it is difficult for clinical studies to analyze the different factors involved in condylar subchondral bone degeneration during TMD development and progression. To date, comorbidity of risk factors such as psychological stress and occlusal abnormalities in the etiology of condylar subchondral bone degeneration has not been established.
Mammalian bones are profusely innervated by sympathetic nerves (Hill and Elde 1991). Studies have shown that the sympathetic tone regulates bone remodeling (Elefteriou et al. 2014). Stressful stimuli such as depression induce bone loss through increased secretion of norepinephrine from the sympathetic nervous system (Yirmiya et al. 2006; Elefteriou et al. 2014). Blocking of sympathetic signals by chemical sympathectomy or a nonselective β-blocker such as propranolol prevented abnormal loading-induced hind limb bone loss in tail-suspension rodent models (Levasseur et al. 2003; Kondo et al. 2005). Blockage of sympathetic signal by superior cervical ganglionectomy, the β-2-adrenergic antagonist butoxamine, or knockout of β1/2-adrenergic receptors suppressed alveolar bone loss and osteoclast hyperactivities induced by occlusal hypofunction (Shimizu et al. 2011) or tooth movement (Kondo et al. 2013; Cao et al. 2014; Sato et al. 2014). Conversely, activation of sympathetic signals by isoproterenol, a β-adrenergic receptor agonist, accelerated alveolar bone loss induced by experimental tooth movement (Kondo et al. 2013).
Aberrant biomechanical loading resulting from unilateral anterior crossbite (UAC) induced subchondral bone loss and increased osteoclastic activity, with concomitant cartilage degradation in the mouse temporomandibular joint (TMJ; Liu et al. 2014; Lu et al. 2014). Occlusal disharmony increases extracellular norepinephrine levels in the hypothalamic paraventricular nucleus of rats (Yoshihara et al. 2001). Nevertheless, it is unknown whether UAC and psychological stress contribute synergistically to induce subchondral bone abnormalities during TMJ degradation and whether the sympathetic tone is involved in regulating these risk factors.
In the present study, subchondral bone remodeling of condyles was investigated in a murine model using UAC and/or chronic immobilization stress (CIS). The CIS model has been well established for generating endogenous psychological stressed in rodents (Campbell et al. 2012; Rahman et al. 2014) and inducing bone loss in their vertebral body and alveolar and distal femoral bones (Patterson-Buckendahl et al. 2001; Semenoff-Segundo et al. 2012; Elefteriou et al. 2014; Appendix Fig. 1). Intraperitoneal injection of 6-hydroxydopamine (6-HD) is a well-established chemical sympathectomy technique (Sherman and Chole 2000; Lucas et al. 2013). This technique was applied to UAC and/or CIS mice to identify the effect of generic blockage of sympathetic signals on condylar subchondral bone loss. 6-HD is a sympatholytic agent with specific toxic effects on terminal sympathetic nerve fibers. Injection with 6-HD inhibits chronic stress-induced bone resorption in the proximal tibia and distal femur of rodents (Hu et al. 2010; Appendix Fig. 1). To clarify the effects of norepinephrine on bone remodeling, mesenchymal stem cells (MSCs)—the progenitor cells of osteoblasts that play important roles in regulating osteoclast activities—were isolated from condylar subchondral bone and stimulated by norepinephrine. Expression of MSC genes related to osteoblast and osteoclast differentiation was investigated. The null hypotheses tested were as follows: (1) The combined effects of UAC and psychological stress are not different from those resulting from individual morbid factors alone in inducing condylar subchondral bone abnormalities. (2) The sympathetic signal is not contributory to the regulation of potential comorbid factors involved in condylar subchondral bone loss.
Materials and Methods
Groups and Sampling
In total, 112 C57BL/6J female mice (17 to 19 g) were obtained from the Animal Center of Fourth Military Medical University. Animal care and treatment were performed according to institutional guidelines approved by the university’s ethics committee. The mice were randomly divided into 7 groups (n = 16 per group; Fig. 1):
Control: sham
6-HD(–): CIS, UAC, CIS + UAC
6-HD(+): CIS, UAC, CIS + UAC
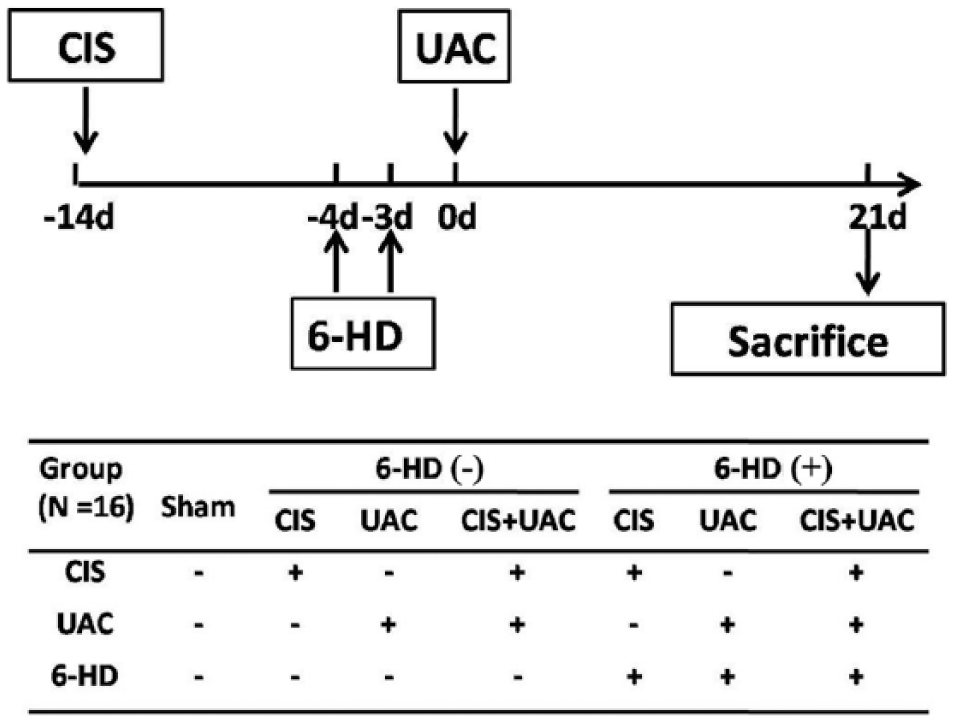
Outline of experimental design. 6-HD, 6-hydroxydopamine injection; CIS, chronic immobilization stress, UAC, unilateral anterior crossbite.
To induce CIS, each mouse was placed in a ventilated 50-mL laboratory conical tube for 4 h daily; sham operation was performed by removing the mouse immediately after placing it into the tube (Campbell et al. 2012). To establish UAC, metal tubes were bonded onto the mouse’s left maxillary and mandibular incisors to create unilateral anterior crossbite; sham operation was achieved without permanent bonding of the UAC prosthesis (Liu et al. 2014; Lu et al. 2014). For combined CIS and UAC, 2 wk of CIS pretreatment was performed before UAC treatment; sham operation was similarly performed for CIS and UAC. To induce acute peripheral sympathectomy, half the mice treated by CIS, UAC, or CIS + UAC were injected with 2 intraperitoneal doses of 6-HD (100 mg/kg on the first day and 250 mg/kg on the second day; Lucas et al. 2013). The other mice were injected with a placebo vehicle (phosphate buffered saline). The initial injection time of 6-HD or vehicle was 10 d after CIS or 4 d before UAC. Mice in CIS and CIS + UAC groups were sacrificed after 5 wk of CIS, while mice in the UAC group were sacrificed after 3 wk of UAC induction (Fig. 1).
Four days before sacrifice, calcein (5 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) was injected into 5 of the 16 mice in each group for examination of in vivo new bone formation. Because no differences in degraded changes were found between the left and right condyles in UAC mice (Liu et al. 2014; Lu et al. 2014), left condylar tissue blocks from each mouse were embedded in methylmethacrylate for measurement of calcein-labeled areas, and right condyles were used for micro–computer tomography (GE eXplore Locus SP, London, UK). Sera derived from 5 unlabeled mice in each group were collected for measurement of serum norepinephrine concentrations; subchondral bone norepinephrine levels were also determined using the condyles of these 5 mice. Left condylar tissue blocks were harvested from 6 additional unlabeled mice, fixed, decalcified, and embedded in paraffin for preparation of 5-μm-thick sagittal sections. The sections were stained with hematoxylin and eosin and tartrate-resistant acid phosphatase (TRAP). The right condylar subchondral bones were harvested for real-time polymerase chain reaction (PCR) analysis of genes related to osteoblast and osteoclast differentiation.
Norepinephrine Determination
To examine whether norepinephrine level in blood (general) and/or bone (local) was associated with subchondral remodeling induced by CIS and/or UAC, norepinephrine contents in the serum and condylar subchondral bone were measured using enzyme-linked immunosorbent assay (BA E-5200, Rocky Mountain Diagnostics, Colorado Springs, CO, USA), as previously described (Flierl et al. 2007; see Appendix).
Micro–computed Tomography
Three-dimensional trabecular microstructure of condylar subchondral bone was acquired and analyzed by micro–computed tomography (Jiao et al. 2014). Briefly, 2 cubes (each 0.3 × 0.3 × 0.3 mm) were selected from the midcenter and midposterior of condylar subchondral bone. Within these selected regions, bone mineral density (BMD), bone volume fraction (BV/TV), trabecular thickness, trabecular number, and trabecular separation (Tb.Sp) were determined using MicroView Advanced Bone Analysis 2.1.2 software (GE Healthcare, Pittsburgh, PA, USA).
Histomorphometry
Unstained sections with calcein labeling were observed using a fluorescence microscope (BX-60, Olympus, Japan). Two square frames (each 0.3 × 0.3 mm) under the osteochondral interface were selected from the midcenter and midposterior of the condyle. Within the selected squares, the percentage areas of the fluorescent regions, representing the bone formation rate, were measured (Jiao et al. 2010). TRAP staining (387-A, Sigma-Aldrich) was performed to examine osteoclast activity of the condylar subchondral bone. Osteoclast number (Oc.N) and the percentage of osteoclast surface area with respect to bone surface (Oc.S/BS) were measured within the selected frames (Jiao et al. 2011; Jiao et al. 2014). Hematoxylin and eosin–stained sections were used for the measurement of cellular density and thickness of condylar cartilage, as previously described (Liu et al. 2014; see Appendix).
Real-time PCR
Gene expression of osterix (OSX), type I collagen, osteocalcin (OCN), RANKL, and OPG were detected by real-time PCR (Jiao et al. 2013; see Appendix). Primers for target genes were listed in Appendix Table.
Norepinephrine Stimulation of MSCs
MSCs were isolated from the condylar subchondral bone of additional mice and cultured following an established protocol (Yamaza et al. 2011). Characterization of the harvested MSCs was performed by flow cytometry analysis of their cell-surface markers and by testing of their osteogenic, adipogenic, and chondrogenic differentiation (see Appendix). The MSCs seeded in 24-well plates were stimulated by adding 1μM NE (N5785, Sigma-Aldrich) to the osteogenic differentiation medium (7531, ScienCell, USA). mRNA levels of OSX, type I collagen, and OCN were analyzed by real-time PCR after 3, 7, or 14 d. In addition, seeded MSCs were stimulated by 1Μm norepinephrine for 2, 6, or 24 hr for determination of RANKL and OPG mRNA expressions. Each test was repeated 3 times, and the average values were used for statistical analyses.
Statistical Analyses
Data were analyzed separately by 1-way analysis of variance and Tukey-Kramer multiple comparison procedures, after affirming the normality and homoscedasticity assumptions of the data sets. When these assumptions were violated, the data sets were nonlinearly transformed prior to the application of parametric statistical procedures. P values <0.05 were considered to be significantly different.
Results
Body Weight
No significant change in body weight was observed in any of the groups during the course of the study (data not shown).
Norepinephrine Level in Serum and Condylar Subchondral Bone
Compared to those of the sham group (Fig. 2), bone norepinephrine levels—but not serum norepinephrine levels—significantly increased in the CIS, UAC, and CIS + UAC groups (P < 0.05). Bone norepinephrine levels in the CIS + UAC groups were significantly higher than those of the CIS and UAC groups (P < 0.05).
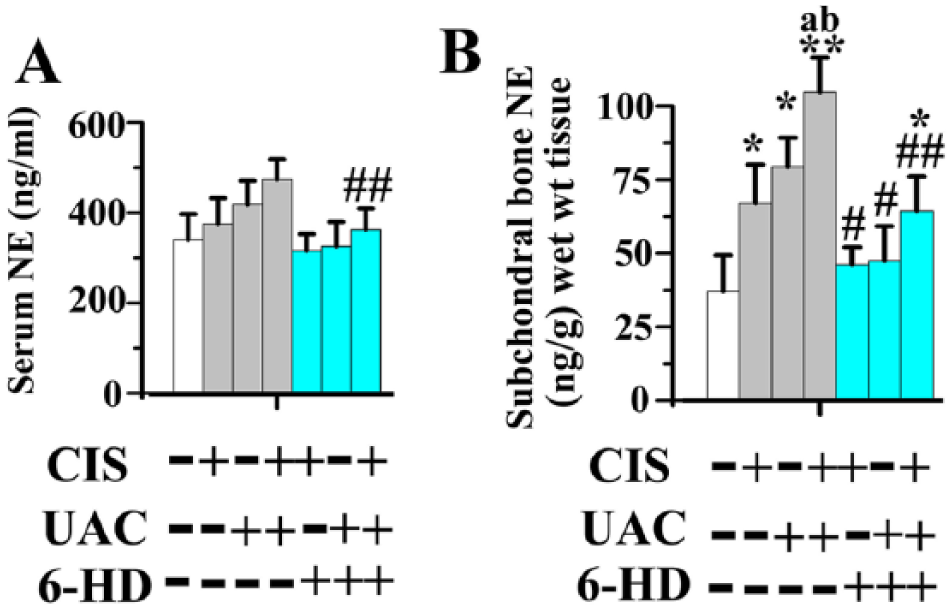
Norepinephrine (NE) levels in serum (
In mice injected with 6-HD, the bilateral ptosis observed within 6 h of initial injection was indicative of the success of sympathectomy (Appendix Fig. 2; Sherman and Chole 2000). Sympathectomy with 6-HD significantly decreased serum norepineprhine levels in the 6-HD(+) CIS + UAC group and bone norepinephrine levels in all groups when compared to those of the vehicle-treated counterparts (P < 0.05). Bone norepinephrine levels in the 6-HD(+) CIS and 6-HD(+) UAC groups were not significantly different from the sham group, while that in 6-HD(+) CIS + UAC group was significantly higher than that of the sham group (P < 0.05; Fig. 2).
Microstructure and Remodeling Activities of Condylar Subchondral Bone
Subchondral bone in the sham group was regularly aligned with bone marrow cavities evenly distributed across the entire joint (Fig. 3A). Newly formed subchondral bone labeled by calcein exhibited thick and well-connected trabeculae in which very few osteoclasts could be identified by TRAP staining (Fig. 4A).
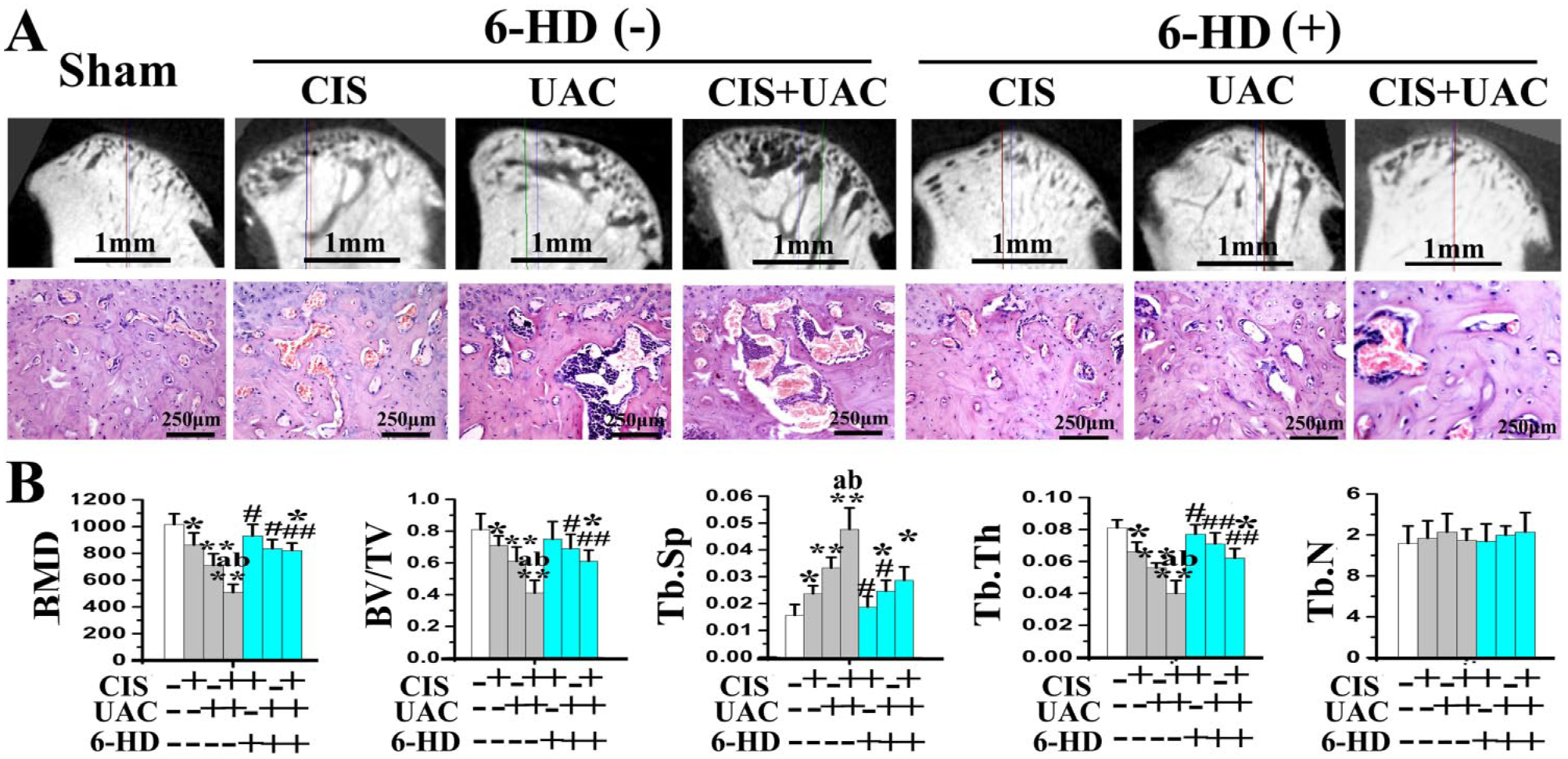
Micro–computed tomography and hematoxylin and eosin staining of mandibular condylar subchondral bone of mice treated by CIS and/or UAC, with 6-HD or placebo vehicle injection. (
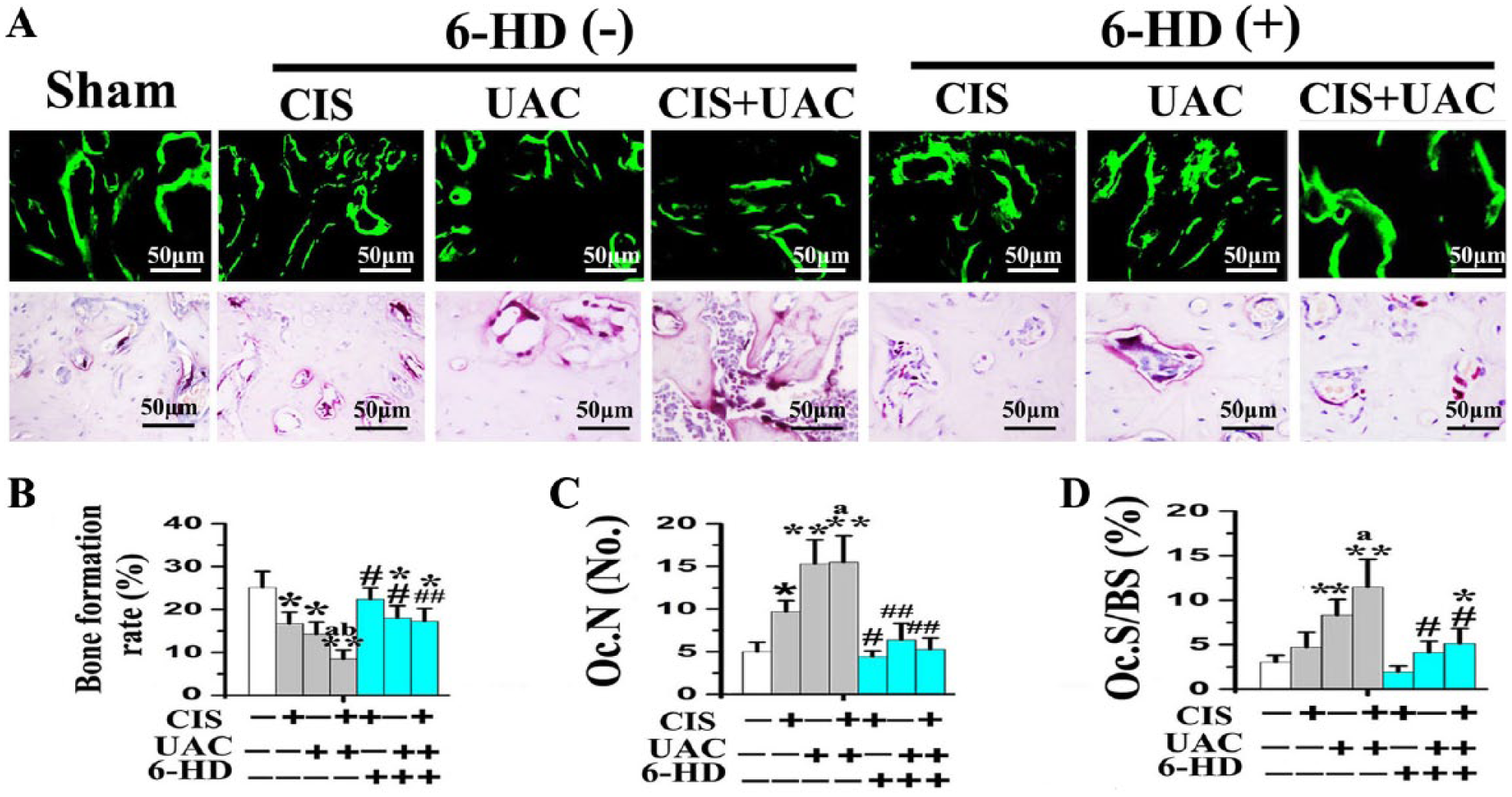
New bone formation and osteoclastic bone resorption in condylar subchondral bone of mice treated by CIS and/or UAC, with 6-HD or placebo vehicle injection. (
In the CIS and UAC groups, localized subchondral bone loss and enlarged bone marrow cavities were observed in the center and posterior condyles (Fig. 3A). Newly formed trabecular bone was sparser and thinner, as revealed by calcein labeling (Fig. 4A). In those groups, data analyses revealed decreased BMD, BV/TV (Fig. 3B), bone formation rate (Fig. 4B), and OSX, Col1, and OCN expression (Fig. 5A) but increased Tb.Sp (Fig. 3B), Oc.N, Oc.S/BS (Fig. 4B), and RANKL expression and RANKL/OPG ratio (Fig. 5A). In the CIS + UAC group, subchondral bone lesions were larger and more severe when compared to those in CIS or UAC groups (Fig. 3A). These morphologic observations were also supported by decreased BMD (Fig. 3B), bone formation rate (Fig. 4B), and OSX, Col1, and OCN expression (Fig. 5A) and increased Tb.Sp (Fig. 3B) and RANKL expression and RANKL/OPG ratio (Fig. 5A; all P < 0.05).
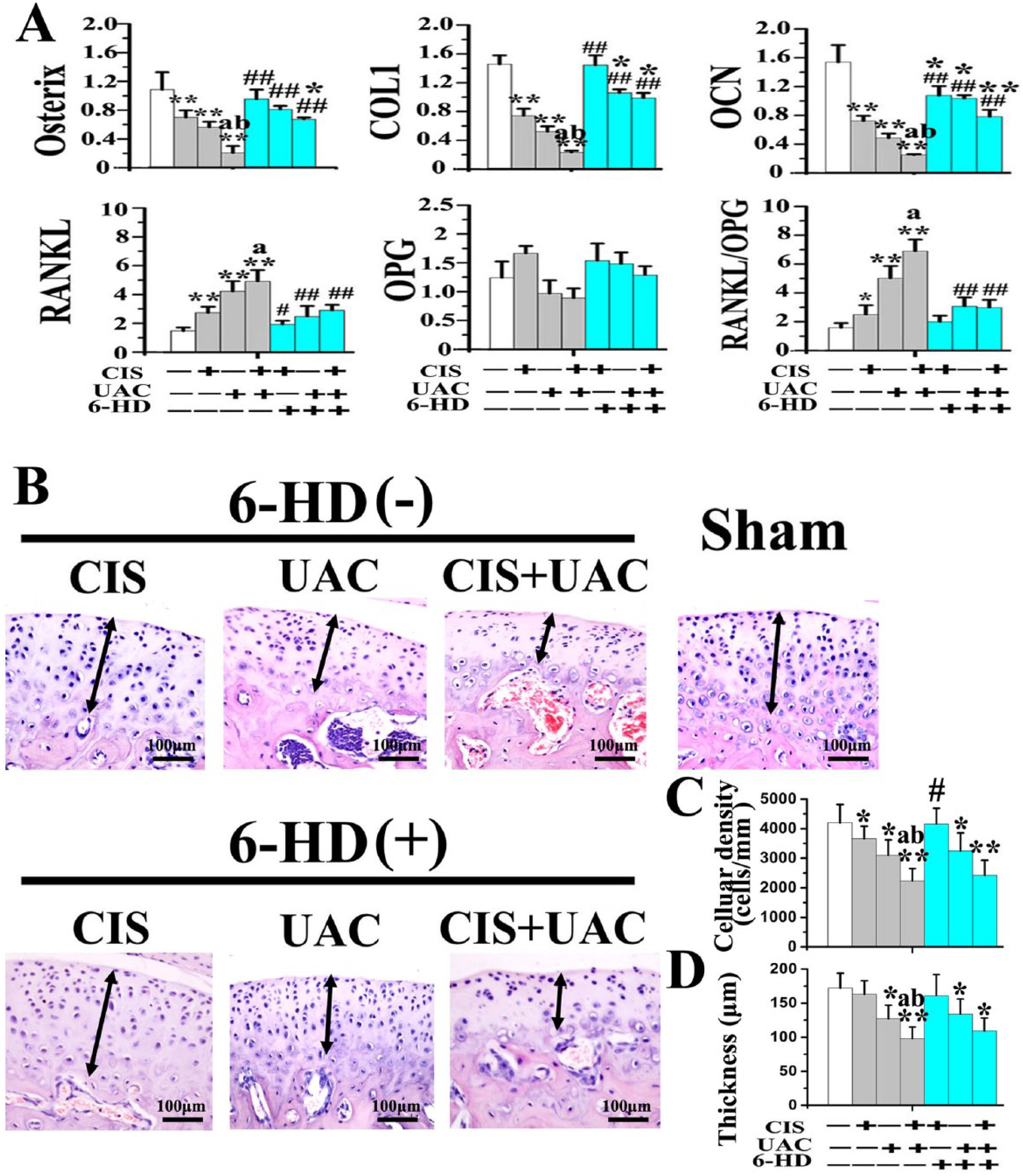
Expression of genes related to osteoblastic and osteoclastic differentiation in the condylar subchondral bone and the condylar cartilage changes of mice treated by CIS and/or UAC, with 6-HD or placebo vehicle injection. (
Sympathectomy with 6-HD reversed subchondral bone loss in the UAC, CIS, and UAC + CIS groups (Fig. 3A). This observation was supported by significant increases in BMD, BV/TV, trabecular thickness (Fig. 3B), bone formation rate (Fig. 4B), and OSX, Col1, and OCN expression (Fig. 5A), as well as decreased Tb.Sp (Fig. 3B), Oc.N, Oc.S/BS (Fig. 4B), RANKL expression, and RANKL/OPG ratio (Fig. 5A), when compared to the vehicle-treated counterparts (all P < 0.05). After 6-HD treatment, the microstructure and remodeling activities of condylar subchondral bone in the 6-HD(+) CIS and 6-HD(+) UAC groups were not significantly different from the sham group (Fig. 3, Fig. 4; P > 0.05).
Condylar Cartilage Changes
Decreased chondrocyte density in the CIS or UAC groups and decreased cartilage thickness in the UAC group were observed comparing to those in the sham controls (Fig. 5B–D; P < 0.05). In the CIS + UAC subgroups, cartilage degradation taking the form of cell-free areas was larger and more severe when compared to those in CIS or UAC groups. These morphologic observations were supported by decreased cellular density and cartilage thickness (Fig. 5B–D; P < 0.05).
Sympathectomy with 6-HD increased chondrocyte density in 6-HD(+) CIS group when compared to that in the vehicle-treated 6-HD(–) CIS counterpart (Fig. 5B–D; P < 0.05). However, cellular density and cartilage thickness in the 6-HD(+) UAC and 6-HD(+) CIS + UAC groups were not different from those in the vehicle-treated counterparts and were still significantly lower when compared with the sham group (Fig. 5B–D; P < 0.05).
Effects of Norepinephrine on Condyle-derived MSCs
Condyle-derived MSCs were positive for MSC-associated markers (SCA-1 and CD73) but failed to express hematopoietic markers (CD45 and CD34). These cells also possessed osteogeneic, adipogenic, and chondrogenic differentiation capacities (Appendix Fig. 3A). Compared to that of the vehicle-treated controls, norepinephrine decreased the mRNA expression of OSX, Col1, and OCN by MSCs after 7 and 14 d of stimulation (Appendix Fig. 3B; both P < 0.05) but had no significant effect at 3 d (P > 0.05). Expression of RANKL and RANKL/OPG ratio by MSCs increased maximally after 2 hr of stimulation (Appendix Fig. 3C; P < 0.05); these activities decreased progressively with prolonged stimulation times. There was no significant difference between the 24-h norepinephrine stimulation group and the time-matched control (Appendix Fig. 3C; P > 0.05).
Discussion
Remodeling of mandibular condylar subchondral bone occurs in the early stages of TMJ-osteoarthritis (TMJ-OA), which is one of the most serious subgroups of TMD (Zhang et al. 2013). Loss of subchondral bone further triggers degradation of the overlying cartilage during TMJ-OA progression (Karsdal et al. 2014; Jiao et al. 2014). These results suggest an etiologic role for subchondral bone in TMJ-OA pathology. In the present murine model, although the use of CIS or UAC alone induced condylar subchondral bone loss, 2 factors behaved synergistically in producing degenerative condylar changes, resulting in more extensive subchondral bone loss and cartilage degradation. Thus, the first null hypothesis has to be rejected—that the combined effects of UAC and psychological stress are not different from those resulting from the individual morbid factors alone in inducing condylar subchondral bone abnormalities.
Subchondral bone norepinephrine level increased significantly during bone loss induced by CIS or UAC. Additional increase in bone norepinephrine level occurred when UAC and CIS were applied in combination. By contrast, sympathectomy by 6-HD reduced bone norepinephrine level and reversed subchondral bone loss induced by CIS, UAC, and more significant, CIS + UAC. Collectively, these data suggest that the pathogenic effects of CIS or UAC were regulated by increased bone norepinephrine level within the subchondral bone regions, which in turn activated the sympathetic signal. Thus, the second null hypothesis has to be rejected—that the sympathetic signal is not contributory to the regulation of potential comorbid factors involved in condylar subchondral bone loss.
Although the relationship between TMD and dysphoria—including depression, anxiety, tension, and stress—has been suggested by several clinical reports (Slade et al. 2007; Fillingim et al. 2011; Calixtre et al. 2014), the pathogenic mechanism underlying such a relationship remains obscure. Epidemiologic studies have implicated that psychological stresses such as depression and anxiety are responsible for reduction in BMD of the lumbar spine and proximal femur and increased incidence of osteoporosis (Kahl et al. 2005; Erez et al. 2012). Experimental mice exposed to emotional stimuli, such as chronic depressing stress, exhibit apparent bone loss and impaired trabecular bone structures in their distal femoral metaphysis and vertebral body (Yirmiya et al. 2006; Elefteriou et al. 2014). Thus, CIS was employed in the present work to testify whether endogenous psychological stress could induce degradation of the TMJ. Indeed, results from the present work identified that CIS alone could precipitate subchondral bone loss and decreased chondrocyte density in mouse condyles and further aggravate malocclusion-induced subchondral bone loss and cartilage degradation. This finding paralleled the results obtained from ligature-induced periodontitis animal models indicating that immobilization stress aggravates alveolar bone resorption (Semenoff-Segundo et al. 2012). The present result indicates that subchondral bone lost was accompanied by an increase in osteoclast activity and more severe cartilage degradation because the loss of subchondral bone may have compromised the biomechanical environment of the overlying cartilage and adversely affected its remodeling (Jiao et al. 2014; Karsdal et al. 2014). Combined with the observation that chemical sympathectomy could reverse bone loss and reduce bone norepinephrine level induced by CIS and/or UAC, amplification of the sympathetic signal by increased bone norepinephrine level appears to be a major risk factor of abnormal subchondral bone remodeling in TMJ degradation. β-adrenergic activity plays a pivotal role in sympathetic signal-regulated bone remodeling (Elefteriou et al. 2014). Clinically, administration of propranolol—a nonselective β-adrenergic receptor antagonist—reduces musculoskeletal pain in TMD patients (Tchivileva et al. 2010). The potential effect of propranololon on reestablishing homeostasis of subchondral bone requires further validation. Because chondrocyte density and cartilage thickness in 6-HD(+) UAC and 6-HD(+) CIS + UAC groups were still lower than the vehicle-treated counterparts, other pathogenic mechanism may be involved in the development of cartilage degradation, apart from subchondral bone remodeling induced by norepinephrine.
Occlusal disharmony causes transient increase in norepinephrine level in the vicinity of the hypothalamic paraventricular nucleus of rats (Yoshihara et al. 2001). In the UAC group, bone norepinephrine level increased significantly during the course of subchondral bone loss. Combined with previous studies that showed that the sympathetic signal plays an important role in the bone mechanoadaptive response (Levasseur et al. 2003; Kondo et al. 2005; Shimizu et al. 2011; Kondo et al. 2013; Cao et al. 2014; Sato et al. 2014), the present results supported the speculation that increased release of norepinephrine within the bone marrow could regulate bone metabolism induced by mechanical loading (Serre et al. 1999; Wada et al. 2001).
Recent studies showed that MSCs isolated from the destructive bone microenvironment promote bone resorption by osteoclasts (Xu et al. 2012). The activities of these niche-specific MSCs are regulated by sympathetic signals within the bone marrow (Ehninger et al. 2011). The present in vitro data showed that norepinephrine increased the expression of pro-osteoclastic genes by MSCs immediately, but this effect diminished with time. Conversely, norepinephrine decreased expression of osteoblastic genes by MSCs after 7 d of stimulation (Appendix Fig. 3B–D). These in vitro data supported our in vivo data in that the significant increase in osteoclast activities occurred after 1 wk of CIS or UAC treatment, while the decrease in osteoblast activities was not apparent until 3 wk later. Collectively, these results indicate that increased release of norepinephrine within the TMJ subchondral bone may exert dual effects on bone remodeling by early promotion of osteoclastic activities and late suppression of osteoblastic activities. Admittedly, these in vitro data are preliminary; further studies have to be performed using a β-antagonist such as propanonol to more precisely elucidate the mechanism in which norepinephrine regulates subchondral bone loss in the TMJ.
In conclusion, the present study demonstrates that CIS or occlusion abnormalities induce significant bone loss and increase bone norepinephrine levels in condylar subchondral bone. The combination of these 2 morbidity factors results in more extensive subchondral bone loss and higher level of bone norepinephrine. Chemical sympathectomy by 6-HD decreases subchondral bone norepinephrine level and reverses bone loss in all experimental groups. These findings suggest that pathologic subchondral bone remodeling of the condyle is intimately controlled by the increased release of norepinephrine within bone.
Author Contributions
K. Jiao, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; L.N. Niu, contributed to design, data acquisition, and interpretation, critically revised the manuscript; X.Y. Xu, Y.D. Liu, X.J. Li, contributed to data acquisition and analysis, critically revised the manuscript; F.R. Tay, M.Q. Wang, contributed to conception, design, and data interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
This work was supported by the
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.