
Abstract
Fluoride-releasing restorative dental materials can be beneficial to remineralize dentin and help prevent secondary caries. However, the effects of fluoride release from dental materials on the activity of dental pulp stem cells are not known. Here we investigate whether different fluoride release kinetics from dental resins supplemented with modified hydrotalcite (RK-F10) or fluoride-glass filler (RK-FG10) could influence the behavior of a human dental pulp stem cell subpopulation (STRO-1+ cells) known for its ability to differentiate toward an odontoblast-like phenotype. The 2 resins, characterized by similar physicochemical properties and fluoride content, exhibited different long-term fluoride release kinetics. Our data demonstrate that long-term exposure of STRO-1+ cells to a continuous release of a low amount of fluoride by RK-F10 increases their migratory response to transforming growth factor β1 (TGF-β1) and stromal cell–derived factor 1 (SDF-1), both important promoters of pulp stem cell recruitment. Moreover, the expression patterns of dentin sialoprotein (dspp), dentin matrix protein 1 (dmp1), osteocalcin (ocn), and matrix extracellular phosphoglycoprotein (mepe) indicate a complete odontoblast-like cell differentiation only when STRO-1+ cells were cultured on RK-F10. On the contrary, RK-FG10, characterized by an initial fluoride release burst and reduced lifetime of the delivery, did not elicit any significant effect on both STRO-1+ cell migration and differentiation. Taken together, our results highlight the importance of taking into account fluoride release kinetics in addition to fluoride concentration when designing new fluoride-restorative materials.
Keywords
Introduction
The common feature of dental caries treatment is the selective removal of caries-infected tissue, while leaving intact the caries-affected tissue. The “caries-affected” dentin is characterized by demineralization of the intertubular dentin, deposition of crystals in the tubules, minimal destruction of the collagen matrix, and no bacterial penetration. On the contrary, the “caries-infected” dentin shows distortion of the dentin tubule microstructure, irreversible denaturing of the collagen fibers, and marked bacterial invasion. It is the latter damaged tissue that should be removed during caries excavation since the caries-affected dentin can be physiologically remineralized due to the presence of viable odontoblast cells in the inner layer and of the collagen network still capable of binding calcium and fluoride ions (Ogushi and Fusayama 1975; Kuboki et al. 1977; Fusayama 1979). Minimally invasive dentistry has evolved not only from the increased understanding of the caries process but also from the development of adhesive and biomimetic restorative materials capable of promoting the remineralization of dental lesions (Tyas et al. 2000; Mm et al. 2014). Several studies reported a synergism in the remineralizing potential when restorative material is combined with fluoride (Khoroushi and Keshani 2013).
Nowadays, fluoride-restorative materials (F-RMs) such as glass-ionomers, resin-modified glass-ionomer cements, polyacid-modified composites or compomers, composites, and amalgams are available on the market. Moreover, commercialized F-RMs exhibit a nonconstant rate of fluoride release depending mainly on the material composition and fluoride content (Wiegand et al. 2007). Therefore, one of the as yet unsolved problems relating to the formulation of F-RM is the modulation of the release and bioavailability of the fluoridating agent. In a recent study, we reported the formulation, preparation, and characterization of modified hydrotalcite (LDH-F)–based materials able both to modulate the release of fluoride in a controlled manner for a prolonged time and to induce human dental pulp cell (hDPSC) proliferation and extracellular matrix mineralization (Tammaro et al. 2014).
The hDPSCs, isolated from adult pulp tissue, have demonstrated in vitro a self-renewal capability of mineralizing and differentiating toward odontoblasts (Shiba et al. 1998; Yokose et al. 2000). Although the isolation of hDPSCs can provide a potential stem cell source for studies in regenerative dentistry, their behavior is quite variable (Gronthos et al. 2002; Miura et al. 2003). STRO-1 is a cell surface protein expressed by bone marrow stromal cells used to identify a population of stem cells able to differentiate into dental hard tissue–forming cells (Yang et al. 2007; Yang et al. 2009; Wang et al. 2010). These sorted stem cells (STRO-1+) in high purity might provide a better cell source for therapeutic purposes than heterogeneous unsorted cells.
The aim of this study was to investigate how the fluoride release kinetics from dental resins supplemented with LDH-F (RK-F10) or fluoride-glass filler (RK-FG10) could influence the ability of STRO-1+ cells to migrate and differentiate toward an odontoblast-like phenotype. The 2 resins, characterized by similar physicochemical properties and fluoride content, exhibited different fluoride release kinetics (slow for RK-F10 and rapid for RK-FG10). Here we demonstrated that STRO-1+ cell migration and differentiation into odontoblast-like cells were enhanced by the slower fluoride-releasing material (RK-F10) compared with RK-FG10, which showed a more rapid fluoride release, thus making LDH-F a promising filler for evaluation in clinical trials of minimally invasive dentistry.
Materials and Methods
Preparation and Characterization of Fluoride-releasing Resins
A fluoride-intercalated layered double hydroxide (LDH-F) was obtained as previously described (Tammaro et al. 2014). Commercial light-activated restorative material (RK), provided by Kerr s.r.l. (Scafati, Italy), consists of bisphenol-A glycidyl methacrylate (Bis-GMA), tri-ethylene glycol dimethacrylate (TEGDMA), camphorquinone (CQ), ethoxylated bisphenol A dimethacrylate (EBPADMA), and glass fillers. RK-F10 and RK-FG10 were prepared by adding an amount of either LDH-F or fluoride-glass filler (FG; Kerr s.r.l.) to obtain a final fluoride concentration of about 0.9% by weight in RK.
Specimen disks 14 mm in diameter and 1 mm thick were fabricated using steel molds. The composites obtained were cured by photo-polymerization using a visible light-curing unit with an irradiation time of 60 s. During the experiment, the light intensity was maintained at 550 mW/cm2. RK composite resin was used as a control. RK-F10 and RK-FG10 were characterized by dynamic-mechanical analysis using a dynamic mechanical thermo-analyzer (DMA 2980; TA Instruments, Milan, Italy). The samples were tested by applying a variable flexural deformation in dual cantilever mode. The displacement amplitude was set at 0.1%, whereas the measurements were performed at the frequency of 1 Hz. The temperature range was −50 °C to 150 °C, and the scanning rate was 3 °C/min.
Fluoride Release Study
Weighed disks of RK-F10 and RK-FG10 were placed at 37 °C under magnetic stirring in artificial saliva medium (SAGF, 15 mL) (Wandera et al. 1996). SAGF was prepared from calculated amounts of chemicals supplied by Sigma-Aldrich (Milan, Italy), according to the procedure described in the literature (Schiff et al. 2002; Kocijan et al. 2011).
After the time intervals (every hour for 8 h, then every day for 10 d, and then every week for 3 wk), the free fluoride ion concentration (ppm) was determined using an ion chromatograph (DX 100, Dionex; Thermo Scientific, Milan, Italy) with suppressed conductivity as previously described (McCabe et al. 2002). The analysis was done in triplicate and the values averaged.
Cell Isolation and Magnetic-activated Cell Sorting
The hDPSCs were enzymatically isolated from impacted third molars from 10 adults (18–22 years of age) as previously described (Gronthos et al. 2000; Tammaro et al. 2014). The teeth were obtained in compliance with Italian legislation (including informed consent and institutional review board approval of the protocol number 7413). The hDPSCs derived from all 10 donors were cultured twice until sufficient cells were obtained, and proliferation, clonogenic potential, and stem cell markers were analyzed. Cells were cultured with α–minimum essential medium (α-MEM) supplemented with 15% fetal bovine serum (FBS), 2 mM L-glutamine, 100 mM L-ascorbic acid-2-phosphate, 100 U/mL penicillin-G, 100 mg/mL streptomycin, and 0.25 mg/mL fungizone (HyClone, Milan, Italy) and maintained in 5% CO2 at 37 °C. To obtain STRO-1+ stem cells, hDPSCs were directly sorted from pulp cell cultures at passage 3 with mouse anti–human STRO-1 IgM (Life Technologies, Milan, Italy) with immune magnetic beads according to the manufacturer’s protocol (Dynabeads; Life Technologies). After cell sorting, each of the following experiments was performed in triplicate on pooled STRO-1–sorted cells (STRO-1+ cells).
Cytotoxicity Assay
The effect of selected materials on STRO-1+ cell viability was determined using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay according to the manufacturer’s instructions (Sigma-Aldrich). STRO-1+ cells were plated at 1 × 104 cells/well on RK, RK-F10, and RK-FG10 materials in 24-well flat-bottomed plates in culture medium. After 1, 3, and 7 d, MTT solution was added to each well and incubated for 4 h at 37 °C and 5% CO2. Absorbance was measured at 570 nm using a microplate reader (Cytation 3; AHSI, Milan, Italy). Cells cultured on tissue culture polystyrene were used as the control. The experiment was repeated 3 times and the mean value calculated.
Cell Migration of STRO-1+ Cells by Transwell Chemotaxis Assay
The cell migration assay was performed using a modified Boyden Chamber method (Albini et al. 1987). STRO-1+ cells were cultured for 7 d on RK, RK-F10, and RK-FG10 materials. The cells were then trypsinized and plated on the upper side of the Boyden Chamber insert filter precoated for 2 h at 37 °C with α-MEM containing 1% FBS and 0.25% (w/v) bovine serum albumin (BSA) to facilitate cells’ initial attachment. The medium in the lower chambers was replaced with α-MEM containing transforming growth factor β1 (TGF-β1, 150 ng/mL) or stromal cell–derived factor 1α (SDF-1, 150 ng/mL), and the migration of STRO-1+ cells was monitored. After 24 h, the cells that had migrated to the lower side of the filter were fixed with 4% paraformaldehyde for 30 min, stained with crystal violet for 20 min, and counted under a microscope in 7 predetermined fields. All experiments were independently repeated 3 times.
Odontogenic-related Gene Expression of STRO-1+ Cells by Real-time Polymerase Chain Reaction
Total RNA was extracted from STRO-1+ cells seeded in the presence of resins for 28 d, using TRIzol reagent (Invitrogen, Milan, Italy) according to the manufacturer’s instructions. Total RNA (0.2 µg) was first treated at 37 °C for 30 min with DNase (Promega, Milan, Italy) and then subjected to reverse transcription (RT) with 0.4 µg random hexamers and 20 U AMV reverse transcriptase (Promega) in a 25-µL reaction mixture at 42 °C for 1 h. The resulting mixture was amplified by real-time polymerase chain reaction (PCR) using specific primers for osteocalcin (ocn), matrix extracellular phosphoglycoprotein (mepe), dentin sialophosphoprotein (dspp), dentin matrix protein 1 (dmp1), and glyceraldehyde-3-phosphate dehydrogenase (gapdh) as listed in Table 1.
Sequence of Primers Used in Real-Time Polymerase Chain Reaction.
Real-time PCR assays were run on an Opticon-4 machine (Bio-Rad, Milan, Italy). The reactions were performed according to the manufacturer’s instructions using SYBR Green PCR Master mix (Invitrogen). The PCR conditions were as follows: AmpliTaq Gold DNA Polymerase (Life Technologies) activation for 10 min at 95 °C and 40 cycles at 95 °C (denaturation) for 15 s and 60 °C (annealing/extension) for 1 min. All reactions were run in triplicate and were normalized to the housekeeping gene, gapdh. Relative differences in the PCR results were calculated using the comparative cycle threshold (CT) method. The variations in gene expression are given as arbitrary units.
Statistical Analysis
All quantitative data are presented as the mean ± SD. Each experiment was performed at least 3 times. Student’s t test was used for the fluoride release. Statistical analyses for the cytotoxicity test, cell migration assay, and quantitative real-time PCR were performed by 1-way analysis of variance (ANOVA) with Bonferroni’s post hoc test.
Results
Materials Properties
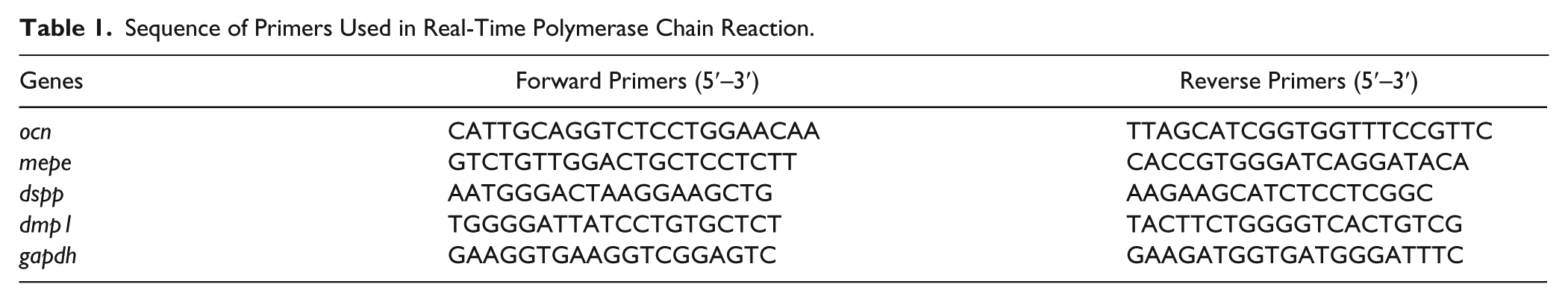
As shown in Figure 1A, the mechanical properties obtained at a wide range of temperatures (0 °C, 37 °C, 50 °C) demonstrated that the values of the elastic modulus of the resin containing the fluoride inorganic solid (RK-F10) showed a greater increase than the resins RK and RK-FG10. In particular, the storage moduli of the RK-F10 were consistently higher than the pristine resin at 37 °C. The observed reinforcement increased with the increase in the filler concentration, as reported by Tammaro et al. (2014). As expected, the deformation at breaking of the RK-F10 was slightly lesser than the pristine resin and RK-FG10 (data not shown). However, the toughness of the 3 materials was similar. Cumulative fluoride release was evaluated for 28 d at 37 °C in artificial saliva medium (SAGF), as shown in Figure 1B. The analyses were done every hour for the first 8 h, then daily for the first week and weekly until the end of the experiment. A time-dependent increase in the fluoride content was observed in SAGF for RK-F10 with respect to RK-FG10 at all time points tested (P < 0.05). After 7 d of incubation, RK-FG10 released 2.723 ± 0.163 ppm of fluoride, an amount reached as early as 24 h. On the contrary, RK-F10 released 0.750 ± 0.053 ppm after 7 d with a daily mean increase of about 0.1 ppm, reaching a concentration of 1.667 ± 0.116 ppm at the end of the experiment. Nonsignificant differences of fluoride release were observed in cell culture medium (data not shown).
Material properties. (
Effects of Fluoride on STRO-1+ Cell Cytotoxicity and Migration
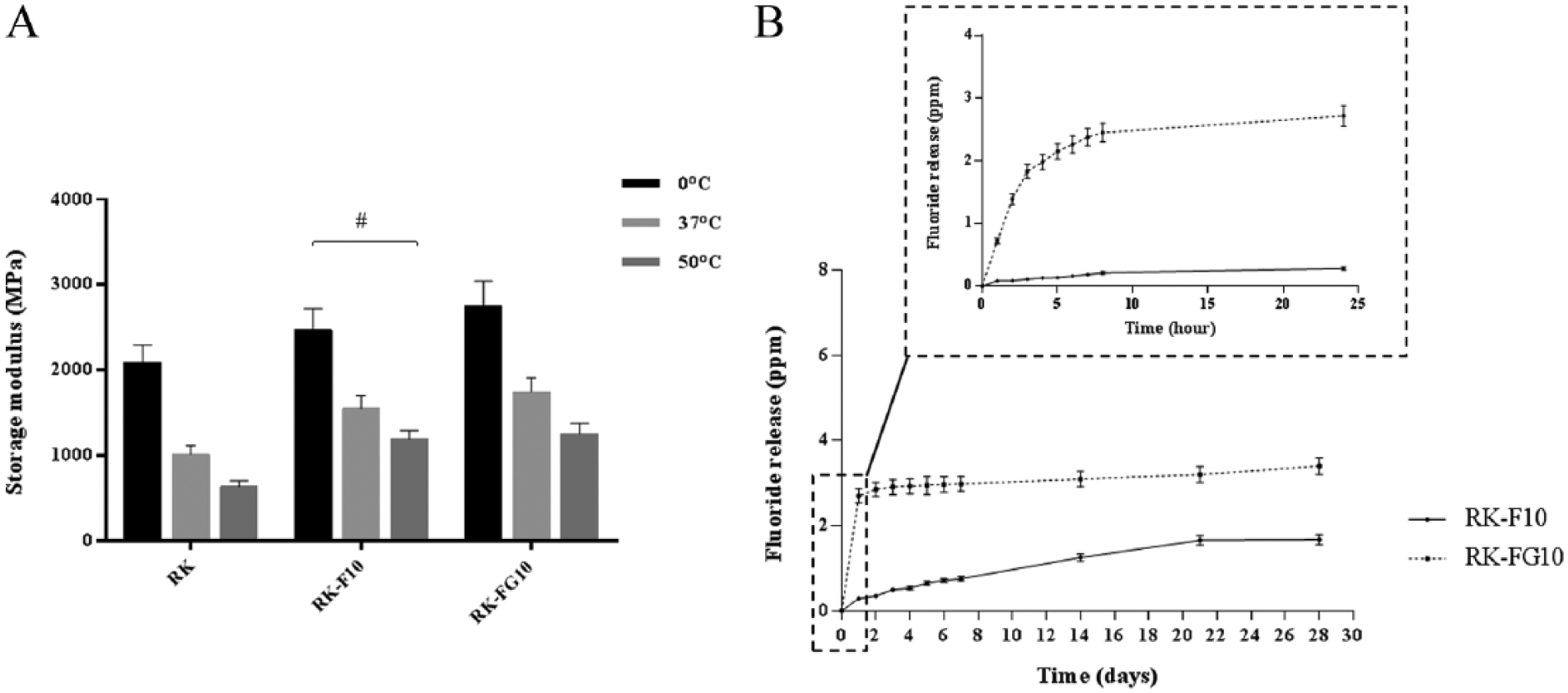
Significant variability in growth patterns, colony-forming unit-fibroblasts (CFU-F) efficiency, and stem cell immunophenotypic profiles have been recorded in hDPSC cultures established from different healthy donors of similar age and stage of third molar root development (Bakopoulou et al. 2011). Because of this, cells with pronounced expression of stem cell properties, including high growth potential and high clonogenicity (CFU-F efficiency >35%), were sorted by magnetic-activated cell sorting (MACS) with a STRO-1 antibody. The amount of sorted stem cells (STRO-1+ cells) ranged from 9% ± 2.5% to 14% ± 3.7%. Immediately after sorting, the STRO-1+ cells were pooled, cultured according to Yu et al. (2010), and used for further experiments. To examine the effects of RK-F10 and RK-FG10 on the migration of STRO-1+ cells, we first determined the viability of the cells using the MTT assay. Cells cultured on tissue culture polystyrene (CTL) and fluoride-free resin (RK) were used as controls. As shown in Figure 2, the cell viability and proliferation were virtually the same for all materials.
Cytotoxic effects of fluoride-releasing materials on STRO-1+ cells after 1, 3, and 7 d of culture. Cells cultured on tissue culture polystyrene were used as control. The bars represent means ± standard deviation (n = 3).
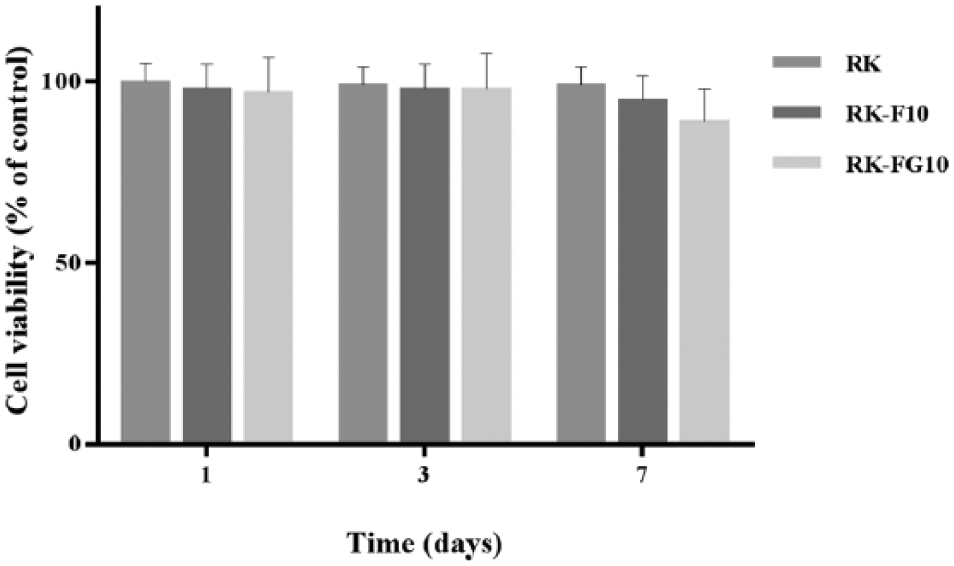
The migratory response of the STRO-1+ cells cultured for 7 d on RK, RK-F10, and RK-FG10 materials (STRO-1+ cells/RK, STRO-1+ cells/RK-F10, and STRO-1+ cells/RK-FG10, respectively) was examined using a modified Boyden chamber in the presence of different chemoattractants (TGF-β1 or SDF-1). STRO-1+ cells/RK-F10 showed a significant ability (P < 0.05 vs. CTL, RK, and RK-FG10) to respond to cytokines, while a modest cell migration in the presence of both chemoattractants was exhibited by STRO-1+ cells/RK-FG10 (Fig. 3).
Chemotactic migration of STRO-1+ cells in the presence of transforming growth factor β1 (TGF-β1) and stromal cell–derived factor 1 (SDF-1). Cell migration assays were evaluated using a modified Boyden chamber. Cells were cultured on RK, RK-F10, and RK-FG10 materials for 7 d before the migration assay was performed. *P < 0.05 versus CTL, RK, and RK-FG10 (n = 3).
Effects of Fluoride-releasing Materials on the Differentiation of STRO-1+ Cells
The expression of a set of genes related to odontoblast differentiation (ocn, dspp, dmp1, mepe) was examined using real-time PCR on STRO-1+ cells over a period of 28 d of culture on RK-F10 and RK-FG10. Cells cultured on polystyrene (CTL) and on fluoride-free resin (RK) were used as controls. Quantitative PCR showed that mepe messenger RNA (mRNA) was significantly higher in STRO-1+ cells/RK-F10 (P < 0.001 vs. CTL, RK, and RK-FG10) as early as 3 d of culture and was downregulated for the rest of the culture period (Fig. 4). The expression of later marker genes (ocn, dspp, dmp1) of odontoblast differentiation was upregulated throughout the experiment in STRO-1+ cells/RK-F10 compared with CTL, RK, and RK-FG10. Although a significant ocn and dmp1 gene expression was observed in STRO-1+ cells/RK-FG10 (P < 0.01 vs. CTL and RK), this resin was not able to modulate the expression of all chosen odontoblast markers throughout the experiment. In fact, ocn, dspp, dmp1, and mepe mRNA levels remained almost constant over the culture period in STRO-1+ cells/RK-FG10.
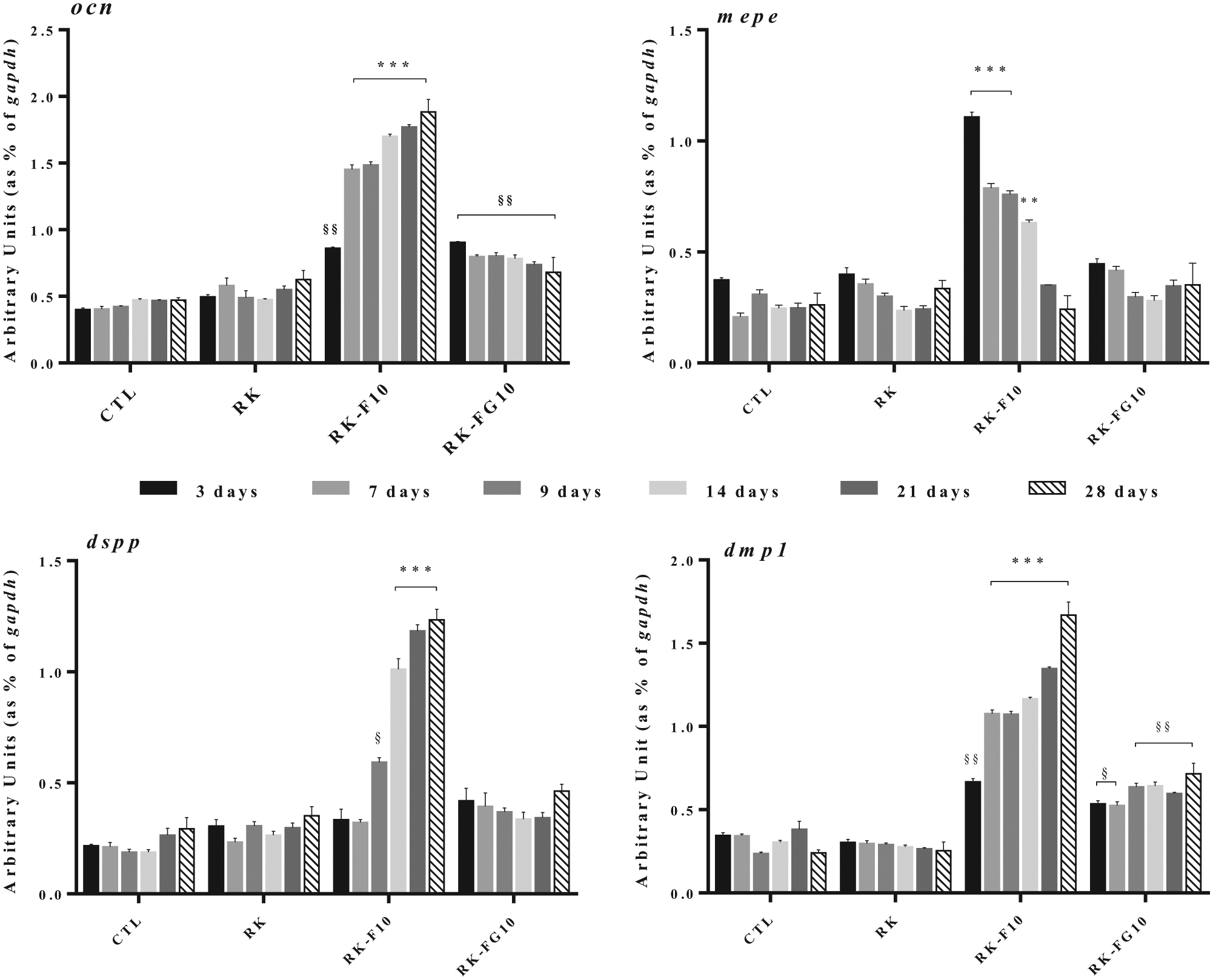
Quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis of ocn, mepe, dspp, and dmp1 in STRO-1+ cells cultured in the presence of RK-F10 or RK-FG10 for 3, 7, 9, 14, 21, and 28 d. Cells cultured on tissue culture polystyrene (CTL) or on RK were used as controls. The target gene expression was normalized to the housekeeping gene gapdh. Relative differences in PCR results were calculated using the comparative cycle threshold (CT) method. The bars represent means ± standard deviation (n = 3). **P < 0.01 versus CTL, RK, and RK-FG10; ***P < 0.001 versus CTL, RK, and RK-FG10; §P < 0.05 versus CTL, and RK; §§P < 0.01 versus CTL and RK.
Discussion
In this study, we demonstrated that the differentiation and chemotaxis capacity of hDPSCs were modulated by long-term controlled delivery of micromolar amounts of fluoride using a modified-hydrotalcite restorative dental resin (RK-F10). The use of fluoride as a supplementary material to improve the dental materials already existing and widely used seems very promising and has led to the commercialization of F-RMs as remineralizing agents in the treatment of “caries-affected” dentin. Since there is no general consensus on the local threshold dose of fluoride or on the duration of exposure to fluoride necessary to elicit a restorative effect, the commercially available F-RMs have dramatically different fluoride-releasing profiles. Recently, several studies showed a definite correlation between fluoride release from different F-RMs and increased mortality of DPSCs. A recent study (Kanjevac et al. 2012) evaluated the potential cytotoxic effects of commercial biomaterials on hDPSCs and showed that F-RMs that released fluoride in high quantities were more toxic to hDPSCs than materials releasing low levels of fluoride. However, there is currently no direct proof that long-term release kinetics of nontoxic amounts of fluoride from F-RMs can affect the migration, proliferation, and differentiation of DPSCs.
The involvement of fluoride at low concentrations (0.05–0.4 ppm) (Mura-Galelli et al. 1992) seems to be necessary for enamel mineralization, but it is of paramount interest also to know whether the same concentrations can modulate DPSC functions. Indeed, DPSCs play a fundamental role in the repair mechanism against dental damage by migrating toward the injury site and, once in place, differentiating into odontoblast-like cells and forming reparative dentin.
In a previous work, we demonstrated that, unlike conventional dental resins, F-RMs modified with hydrotalcite present slow and constant fluoride release kinetics, eliciting beneficial effects on DPSC proliferation and mineralization (Tammaro et al. 2014). In the present study, we analyzed whether the fluoride release kinetics from different F-RMs with the same amount of fluoride can also affect the migration and differentiation of STRO-1+ cells, a DPSC subpopulation able to differentiate into dental hard tissue–forming cells.
It is well known that fluoride can affect sperm chemotaxis and the human osteoblastic cell migratory response to chemotactic factors in a dose-dependent manner (Ohno et al. 2013; Lu et al. 2014). However, no study has investigated the role played by fluoride on DPSC ability to migrate in response to different stimuli or the influence of the fluoride level on cell migration. In regards to cell migration, we demonstrated that long-term exposure of STRO-1+ cells to a continuous release of a low amount of fluoride from RK-F10 resin increased their migratory response to TGF-β1 and SDF-1, both important promoters of pulp stem cell recruitment. On the contrary, the initial burst of fluoride from RK-FG10 resin (RK filled with fluoride-glass filler) led to a higher initial fluoride delivery, which, however, reduced the effective lifetime of the delivery and the biological effects of fluoride on cell chemotaxis. It is possible that the effects of a low concentration of fluoride on modulating cell migration might be related, at least in part, to its well-known ability to activate G protein, initiating signaling cascades that regulate the migration of different cell types (Aoba and Fejerskov 2002; Cotton and Claing 2009). Another important finding in this study is that only STRO-1+ cells cultured on RK-F10 resin had gene expression changes, suggesting they were able to differentiate into functional odontoblast-like cells. Functional differentiation of odontoblasts, the second step in dentinogenesis, requires unique sets of genes to be turned on and off in a growth- and differentiation-specific manner (Linde and Goldberg 1993). To our knowledge, few studies have investigated the effect of long-term fluoride exposure on DPSC differentiation. As reported by Thaweboon et al. (2003), fluoride at low concentrations can stimulate proliferation and differentiation of DPSCs, whereas an inhibitory effect can be observed at higher concentrations. In addition, Wurtz and colleagues (2008) demonstrated that the gene expression profile of the odontoblast-like cell line MO6-G3 changed in response to a low fluoride concentration, without inducing cell stress or apoptosis.
The present study showed that in STRO-1+ cells cultured on RK-F10, the expression patterns of dentin sialoprotein (dspp), dentin matrix protein 1 (dmp1), osteocalcin (ocn), and matrix extracellular phosphoglycoprotein (mepe) indicate complete odontoblast-like cell differentiation. In fact, mepe, an early odontoblast differentiation marker, was significantly upregulated at 3 d of culture on RK-F10 compared with STRO-1+ cells cultured on polystyrene, RK, or RK-FG10. This was followed by a dramatic downregulation over the next 25 d, commensurate with advancing odontoblast differentiation. Although the exact role of mepe as a regulator of mineralization and the mechanisms underlying this function are still controversial, its expression is observed only in immature odontoblasts and becomes downregulated upon odontoblast differentiation (MacDougall et al. 2002; Liu et al. 2005). In addition, ocn, dmp1, and dspp, late markers of odontoblast differentiation responsible for the formation of tissue-specific dentin matrix (Begue-Kirn et al. 1998; Feng et al. 1998; Papagerakis et al. 2002; Qin et al. 2002), were all upregulated in STRO-1+ cells/RK-F10 compared with the controls and STRO-1+ cells/RK-FG10.
In conclusion, the results reported herein highlight the importance of taking into account the fluoride release kinetics in addition to the fluoride concentration when designing new fluoride-restorative materials. In particular, it is important to take into account that the kinetics of fluoride release from dental resins can govern the pulp’s response to restorative procedures and that the pulp reacts to the elements to which it may be exposed through the dental tubules not only during but also after dental restoration involving adhesive technologies (Ferracane et al. 2010; Khoroushi and Keshani 2013).
Author Contributions
A. Calarco, contributed to conception, design and data acquisition, drafted and critically revised the manuscript; A. Di Salle, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; L. Tammaro, O. Petillo, F. Riccitiello, V. Vittoria, contributed to data acquisition, critically revised the manuscript; I. De Luca, S. Mucerino, contributed to data analysis and interpretation, drafted the manuscript; G. Peluso, contributed to conception and design, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
Acknowledgements
We thank Elizabeth McGarry for critical reading of the manuscript and Mario Guarracino and Domenico Maisto (Institute of High Performance Computing and Networking [ICAR]–National Research Council [CNR], Naples, Italy) for their help in statistical analysis.
This work was supported by Progetto PON—“Ricerca e Competitività 2007–2013”—PON01_01802: “Sviluppo di molecole capaci di modulare vie metaboliche intracellulari redox-sensibili per la prevenzione e la cura di patologie infettive, tumorali, neurodegenerative e loro delivery mediante piattaforme nano tecnologiche” and PON01_02512: “Ricerca e sviluppo di bioregolatori attivi sui meccanismi epigenetici dei processi infiammatori nelle malattie croniche e degenerative.”
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.