
Abstract
We report a novel method for the isolation of adult human epithelial stem cells (hEpiSCs) from the epithelial component of the periodontal ligament—the human epithelial cell rests of Malassez (hERM). hEpiSC-rich integrin-α6+ve hERM cells derived by fluorometry can be clonally expanded, can grow organoids, and express the markers of pluripotency (OCT4, NANOG, SOX2), polycomb protein RING1B, and the hEpiSC supermarker LGR5. They maintain the growth profile of their originating hERM in vitro. Subcutaneous cotransplantation with mesenchymal stem cells from the dental pulp on poly-
Introduction
Embryonic stem cell study and lineage tracing of stem cell fate reveal the principles of cell differentiation. Induced pluripotent stem cells are generated from somatic cells by the forced expression of markers of stem cell pluripotency, Oct4, c-Myc, Sox2, and Klf4 or Nanog (Bayart et al. 2013). Reports of genetic and epigenetic aberrations occurring during the stem cell derivation and reprogramming have raised doubts about the application of embryonic stem cells and induced pluripotent stem cells for tissue regeneration (Huch et al. 2015). Evidently human stem cells identified in tumors (Reya et al. 2001) cannot be used for regeneration. Recently, genetically unmodified, noncancer-origin adult human epithelial stem cells (hEpiSCs; Artwert et al. 2012) were successfully cultured from skin, intestinal, pancreatic, and liver stem cells, based on their expression of cell surface proteins (Calloni et al. 2013) integrin-α6 (Nowak and Fuchs 2009) and LGR5 (Huch et al. 2015). Organ regeneration through hEpiSCs requires accessibility of donor tissues, isolation, propagation, and overcoming cell death caused by loss of cell-to-cell contact (Chiarugi and Giannoni 2008). We report here the successful clonal isolation, characterization, and maintenance of integrin-α6+ve cells and LGR5+ve cells of the periodontal ligament (PDL) in a xeno-free, feeder layer–free tissue culture system. These cells are a by-product of orthodontic removal of wisdom teeth and exist around all teeth, throughout adult life, therefore representing an easily accessible homologous donor hEpiSC source.
By definition, stem cells give rise to unaltered daughter cells while also being “progenitors” of somatic cells. As distinct epithelial organs share molecular developmental mechanisms, a common profile emerges for epithelial stem cells (EpiSCs): they are slow cycling and highly enucleated; they are remnants of a developmental process; and they share patterns of gene expression with the cells to which they are precursors. Evidence of stemness consists of the presence of proliferation markers (e.g., Ki-67) and markers of pluripotency and the ability to form organoids (Calloni et al. 2013; Hynds and Giangreco 2013).
The epithelial cell rests of Malassez (ERM) structures are nested within the PDL. In oblique section, they appear like a fishnet, while in transverse section, they liken nests of highly enucleated cells (Rincon et al. 2006). They also become mitotically activated in response to injury, inflammation, or in vitro explantation and can give rise to squamous metaplastic cells, cystic lesions, and ameloblastoid odontogenic tumors (Cerri and Katchburian 2005). They are therefore suspect as the remnant dental hEpiSC niche (Nam et al. 2011). During enamel formation, integrin-α6 is expressed in preameloblasts (Salmivirta et al. 1996), the dental epithelia, and precursors of the ERM. Integrin binds Laminin5-α3 and drives odontogenesis. We therefore focused our study to the investigation of the integrin-α6+ve cells of the PDL as we searched for hEpiSCs. After our first reports on integrin-α6+ve cells (Athanassiou-Papaefthymiou et al. 2010), several others (Xiong et al. 2012; Xiong et al. 2013) reported the ERM origin, pluripotency, and plasticity of ovine integrin-α6+ve cells and confirmed that integrin-α6+ve cells are the ERM cells within the PDL. ERM cells have thus been counted among the ever-increasing variety of the integrin-α6+ve stem cell–rich epithelial compartments that include murine mammary EpiSC and the hair follicle, the organ that most closely resembles the tooth in its development.
EpiSCs in the developing mouse maintain quiescence, survive, proliferate, and differentiate under the control of the Wnt, Hedgehog, and Notch pathways (Reya et al. 2001). Lgr5 has been shown to be a marker of adult murine (Sato et al. 2009) and human (Huch et al. 2015) intestinal EpiSC, with culture conditions established that can overcome anoikis. Lgr5+ve clones isolated by trituration and cultured in matrigel organize into organoids forming complete villar structures. Subsequent discovery of LGR5 among integrin-α6+ve stem cells of the hair follicle and other work established LGR5 as an EpiSC supermarker (Jaks et al. 2008). Finally, Lgr5 marked the EpiSCs of the mouse incisor (Suomalainen and Thesleff 2010) and the ERM (Handrigan et al. 2010; Oka et al. 2012), meriting further investigation into LGR5+ve cells of the human ERM (hERM).
We report here that integrin-α6+ve cells can grow organoids and express the markers of pluripotency, OCT4, NANOG, and SOX2, as well as the stem cell maintenance polycomb protein RING1B and the EpiSC supermarker LGR5. LGR5+ve cells are a subpopulation of the integrin-α6+ve cells, and we were able to isolate and maintain both types of cells in xeno-free culture conditions. Based on expression of markers of pluripotency or genes important for secretory epithelial and dental epithelial function, the genetic profile of LGR5+ve cells gives a first insight into an hEpiSC population of dental interest. Subcutaneous cotransplantation with mesenchymal human dental pulp stem cells (hDPSC) on poly-
Materials and Methods
Extensive experimental details are part of the Appendix.
Cell culture of ERM from PDL explants: Epithelial cell outgrowths from the PDL tissue were grown in the KnockOut DMEM system (Invitrogen, Carlsbad, CA, USA) supplemented with keratinocyte growth factor 2 (fibroblast growth factor 10).
Fluorescence-activated cell sorting: Primary PDL cultures were cell sorted with a PE-anti-Integrin-α6 antibody (eBiosciences, San Diego, CA, USA) and an anti-human LGR5 (Sigma-Aldrich, St. Louis, MO, USA) as described in the Appendix. Cells were harvested by fluorescence-activated cell sorting (FACS) in survival medium in tubes or plated clonally on MaxGel-ECM (Sigma-Aldrich) plates with jagged peptide.
Immunofluorescent immunocytochemistry: Immunofluore-scent cytochemistry was performed at the Microscopy and Image Analysis Laboratory at the University of Michigan.
PCR arrays on LGR5+ve cells: LGR5+ve cell RNA was prepared (PCR RNAqueus Kit, Ambion; Thermo Fisher Scientific, Waltham, MA, USA) according to manufacturer’s instructions. Superscript III (Invitrogen)–generated cDNAs were verified for beta-actin, GAPDH, and CD44 expression (Platinum PCR; Invitrogen) and SABiosciences WNT and Stem Cell (Qiagen, Valencia, CA, USA) human polymerase chain reaction (PCR) microarrays were used with FastStart SYBR Green mix (Roche, Pleasanton, CA, USA) on a 7900H ABI machine (Thermo Fisher Scientific) for real-time PCR.
In vivo experiments: Experiments were performed in accordance with the code of ethics for animal research at the University of Michigan. DPSCs (103) were coseeded with integrin-α6+ve cells (103) on PLLA nanoscaffolds and implanted subcutaneously in the back of nude mice (Jackson Laboratory, Bar Harbor, ME, USA), 1 in each limb quadrant. Mice were sacrificed at 6- and 10-wk time points and analyzed.
Results
EpiSC Properties among the Cells of Human PDL
We verified the presence of hEpiSC-like cells in the PDL with preliminary experiments utilizing a previously characterized population (Xu et al. 2009) depleted of both mesenchymal and endothelial stem cells (STRO1-ve/CD146-ve cells) and thus lacking the corresponding markers STRO1 and CD146 (Fig. 1a). Apart from the ERM, the only other epithelial cells in the PDL are endothelial cells that line capillaries, the CD146+ve cells. CD146-ve cells that are epithelia, can only be ERM. These cells were cultured in selective LHC-9 medium (Shinmura et al. 2008; Shimonishi et al. 2010), and the resulting population expressed the epithelial cell marker E-cadherin, coexpressing amelogenin, an ameloblast marker, in osteogenic induction media (Fig. 1b, c). LHC-9 selected cells could also grow organoids (Fig. 1a).
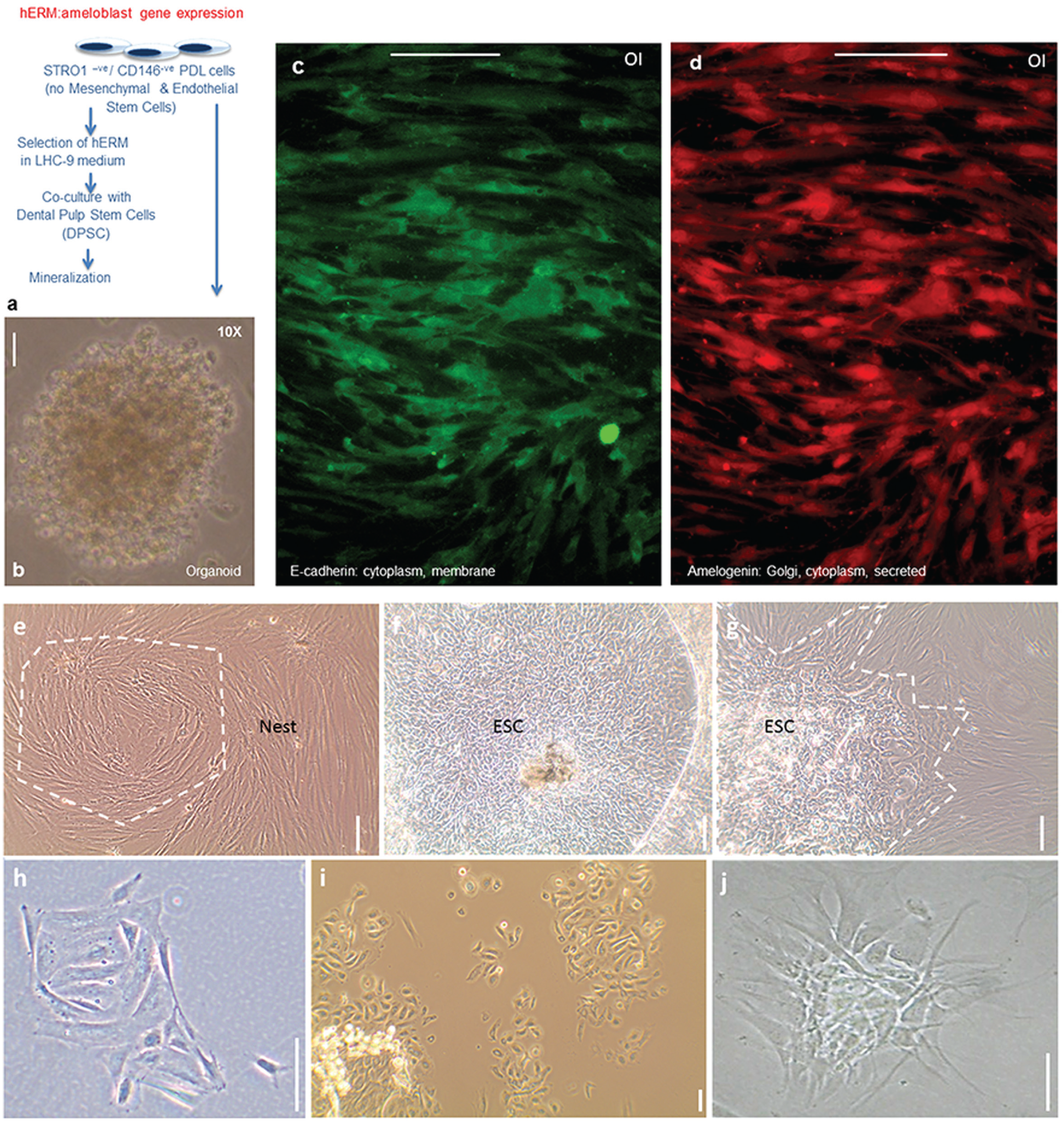
Isolation and initial characterization of human epithelial cell rests of Malassez derived from periodontal ligament (PDL). (
Subsequent studies involved isolating the epithelial cell component from the PDL of healthy third molars from human subjects. To establish a xeno-free culture favoring epithelia, without introducing components from an extratumoral environment, instead of Matrigel, we provided human extracellular matrix and components of human origin interacting with dental epithelia in vivo: collagen IV, laminin (epithelial-binding component), fibronectin (essential for stem cell migration). Passage led to vigorous growth of high cell numbers in primary cultures (Appendix Fig.). Within PDL cells of morphological heterogeneity, islands of rapidly growing epithelial cells, even embryonic stem cell–like cells, could be seen (Fig. 1d–f). Ring cloned colonies from the epithelial cell islands gave cells with epithelial morphology, forming nest-like, net-like structures.
Epithelial cells from the PDL are particularly sensitive to handling during FACS, but we optimized cell survival during the washes, incubation with antibody, and isolation under proprietary conditions. FACS experiments showed that the integrin-α6+ve cells were a subpopulation of keratin-expressing cells (Alam et al. 2011), and LGR-5+ve cells were shown to be a subpopulation of integrin-α6+ve cells (Nam et al. 2011). Here we show that integrin-α6+ve clones from the hERM can be maintained live in 2-dimensional culture resembling their physiologic fishnet appearance with nests of cells extending to touch and connect as if in a nest (Fig. 2a–c). The cells can be passaged multiple times over prolonged periods and can infiltrate PLLA nanoscaffolds. In their physiologic state, ERM cells maintain cell numbers via equal rates of cell proliferation and apoptosis. The slow growth of the clonally selected cells in the absence of mesenchyme is consistent with their in vivo profile. Conditioned media from hDPSC culture alone was not sufficient to produce enhanced proliferation, either because mesenchymal and epithelial cell contact is necessary or because an unidentified, nonsoluble extracellular matrix component produced by the mesenchyme is critical for the proliferation or differentiation of the epithelial cells. Integrin-α6+ve hERM also grow organoids when seeded in Matrigel at clonal density (Fig. 2f).
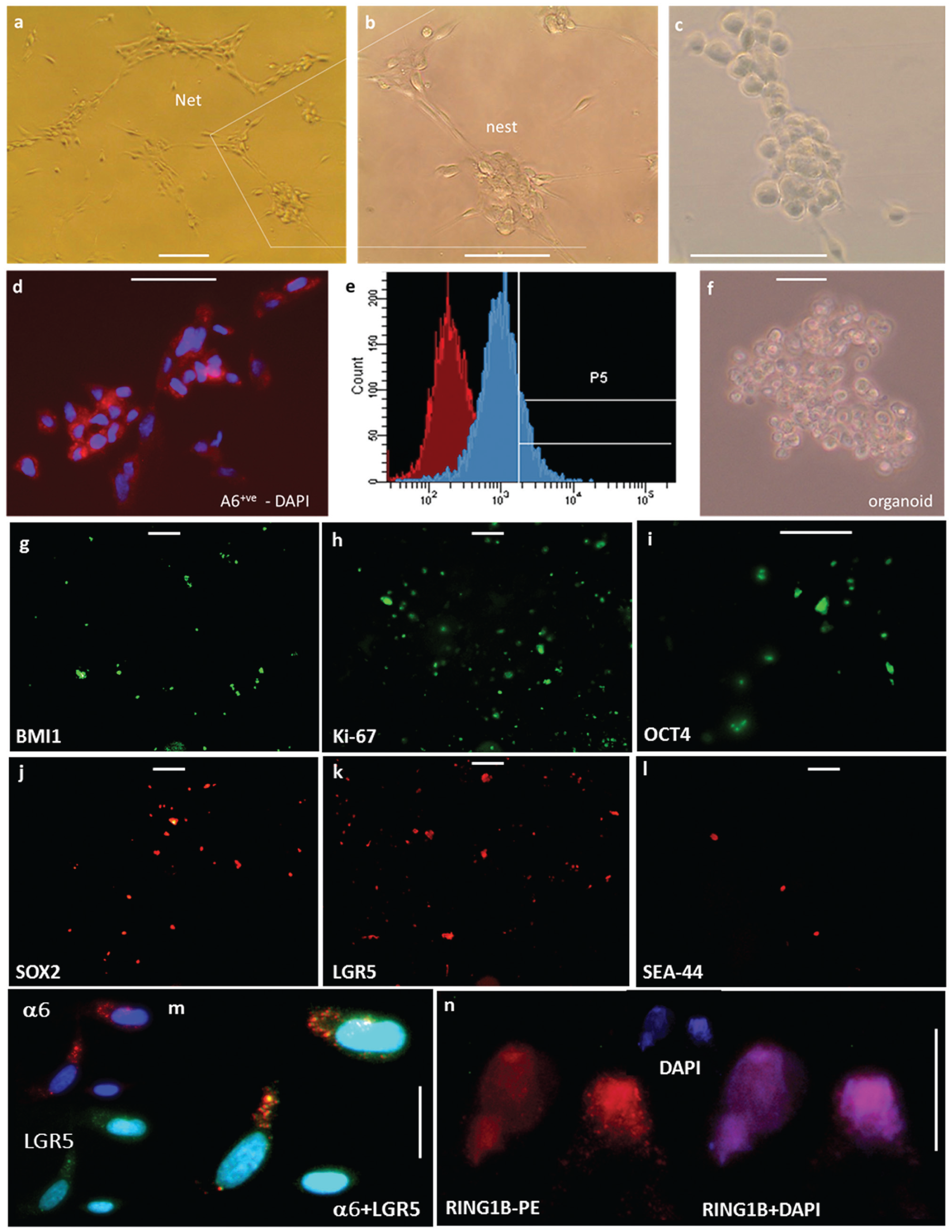
Isolation, clonal expansion, in vitro stem cell properties and in vivo mineralization potential of integrin-α6+ve epithelial cell rests of Malassez. (
Integrin-α6+ve and LGR5+ve Cells Show Profiles of hEpiSC
Our integrin-α6+ve cells (Fig. 2d, e) grow organoids in 3-dimensional culture (Fig. 2f) and express stem cell markers SOX2, NANOG, OCT4, and LGR5 and the polycomb protein RING1B, which is required for stem cell maintenance (Fig. 2g–n). They thus show hallmarks of pluripotency. As the integrin-α6+ve cell population has been widely established as stem cell rich, we consider the integrin-α6+ve hERM, an hEpiSC-rich PDL population.
We further examined the expression of stem cell markers in LGR5+ve hERM cells (Fig. 3). LGR5 was detected by immunofluorescence in our primary culture of the PDL and was coexpressed with Ki-67, a marker of cycling cells (Fig. 3d–g). We succeeded in live sorting of LGR5+ve cells among the PDL (Fig. 3m) and in identifying factors critical for cell survival. Live LGR5+ve cells account for 0.8% and up to 2% of early passage cells (passages 0 to 3). LGR5 is being expressed in integrin-α6+ve cells (Fig. 2l). Other stem cell markers, such as epCAM, NOTCH, and CD34, are also expressed among our PDL tissue culture. By immunocytochemistry of cultures of live-sorted LGR5+ve cells from hERM, we found them positive for Ki-67, for markers of pluripotency OCT4 and NANOG, and for p63, which is at the heart of epithelial cell and tooth development (Laurikkala et al. 2006). As expected by their expression of integrin-α6, our LGR5+ve cells also express cytokeratin 14, another marker of Mallassez origin.
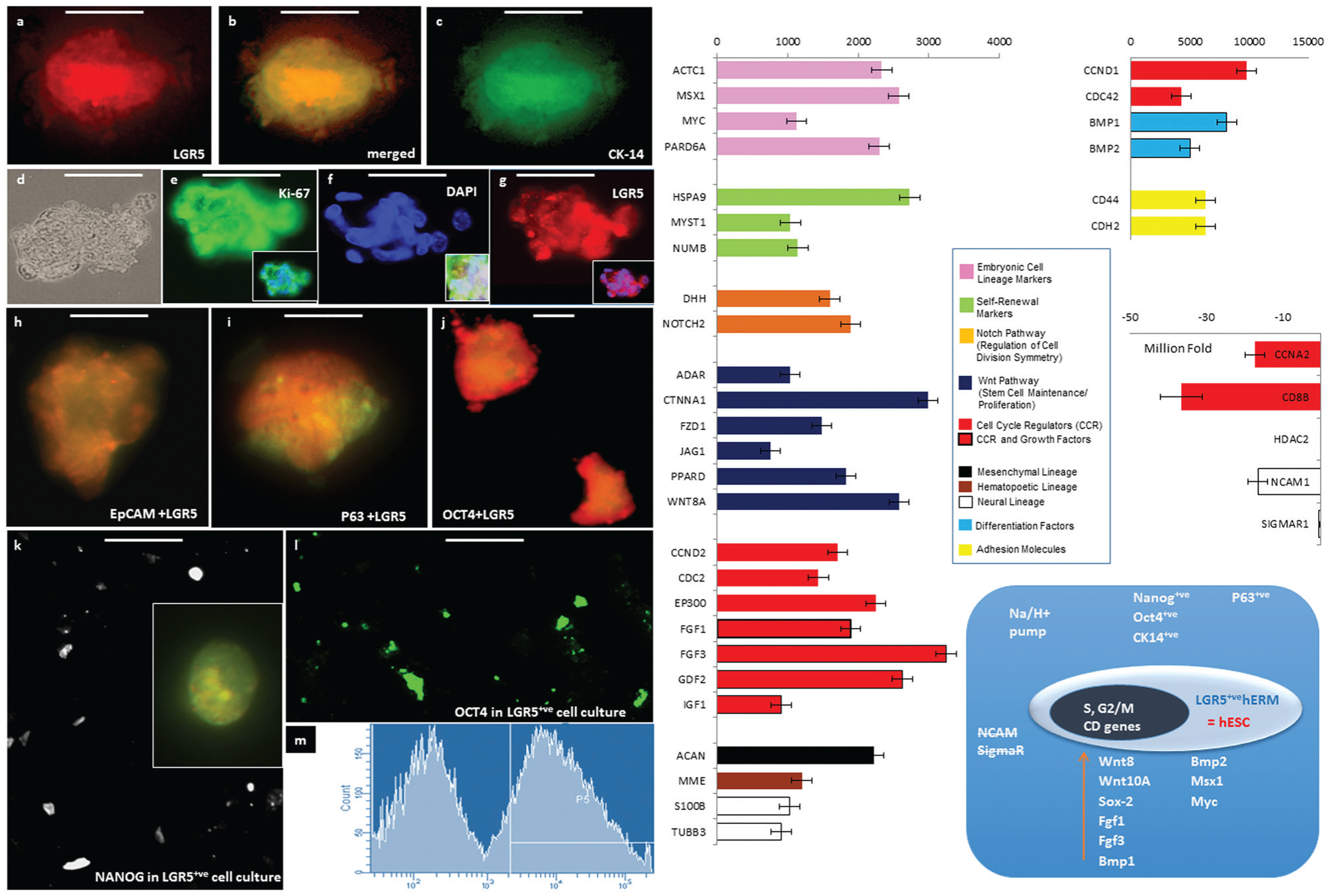
Characterization of LGR5+ve adult human epithelial stem cells (hEpiSCs). (
The RNA of LGR5+ve cells and matched donor–origin control PDL cells was then used to test for the expression of 165 genes using the SABiosciences WNT and Stem Cell PCR arrays. The differential display of genes associated with the stem cell phenotype in our LGR5+ve cells (Fig. 4), as opposed to the originating cells, showed enrichment in a variety of genes, including genes associated with stem cell maintenance and expressed in embryonic stem cells, as well as cell cycle regulators and growth factors. Elevated WNT family member expression was general as expected by the role of Wnt in tooth development (Suomalainen et al. 2010) with highest for WNT11 and FRIZZLED1 as well as WNT8A and WNT10A, which characterize ectodermal origin during embryonic development (Niehrs 2012) and are essential regulators of mesenchymal morphogenesis. WNT10A deletion leads to tooth agenesis (Yang et al. 2015). This result is consistent with our finding that full mineral formation in vivo occurs only when the hEpisSC-rich cells are coimplanted with hDPSC. In addition to the expression of OCT4 and NANOG, which was verified with immunocytochemistry, the arrays showed enrichment of MYC and SOX2. Not only is SOX2 an inducer of pluripotency, but it also marks epithelial competence to generate teeth in mammals (Juuri et al. 2013). NCAM, a molecule associated with mesenchymal stem cell lineage, was completely suppressed. We were surprised to see the muscle differentiation gene MYOD also enriched, and it is worth exploring whether integrin-α6+ve hEpiSCs can give rise to myoepithelial cells. Genes implicated in dental epithelium development were increased, such as PITX2, an exclusive marker of dental epithelium FGF1 and FGF3 and BMP1 and BMP2. Most pronounced was the increase of the CFTR gene, a Na/H+ pump that is important for ameloblast mineralization but also for the function of mucosal epithelia. In addition to OCT4 and NANOG, the arrays showed enrichment of MYC, SOX2, and T (brachyury). Notably, brachyury also acts in concert with Wnt8. While NCAM, SIGMAR, and GUSB were completely suppressed, MYOD was surprisingly enriched, but this was consistent with our later observation of myoepithelial cells of human origin in studies of integrin-α6+ve dental hEpiSCs implanted in nude mice. Also surprising was the increase in COL1A1, but like WNT8 expression, it is consistent with active transcription from chromosome 17 alterations that are associated with cancer development in many epithelial tissues. Expression of collagen genes found on other chromosomes was not enriched. In in vitro cultures, epithelial cells secrete their own extracellular matrix and assemble a laminin-rich basal matrix to establish the basal cue that determines polarity (Manninen 2015). Several genes implicated into cell cycle control showed significant changes in their expression levels: 1) CCND1, which is required for the G1-to-S transition, was remarkably reduced; 2) CDC2, which is also required for a coherent S phase and for the G2-to-M transition, was increased; and 3) CDC42, which transduces signals from integrins, GPCRs, and growth factors to p21 family kinases, was highly enriched. PARD6 was also elevated—a semi-cd42 gene controlling asymmetric cell division and polarity and playing a role in epithelial-to-mesenchymal transitions.
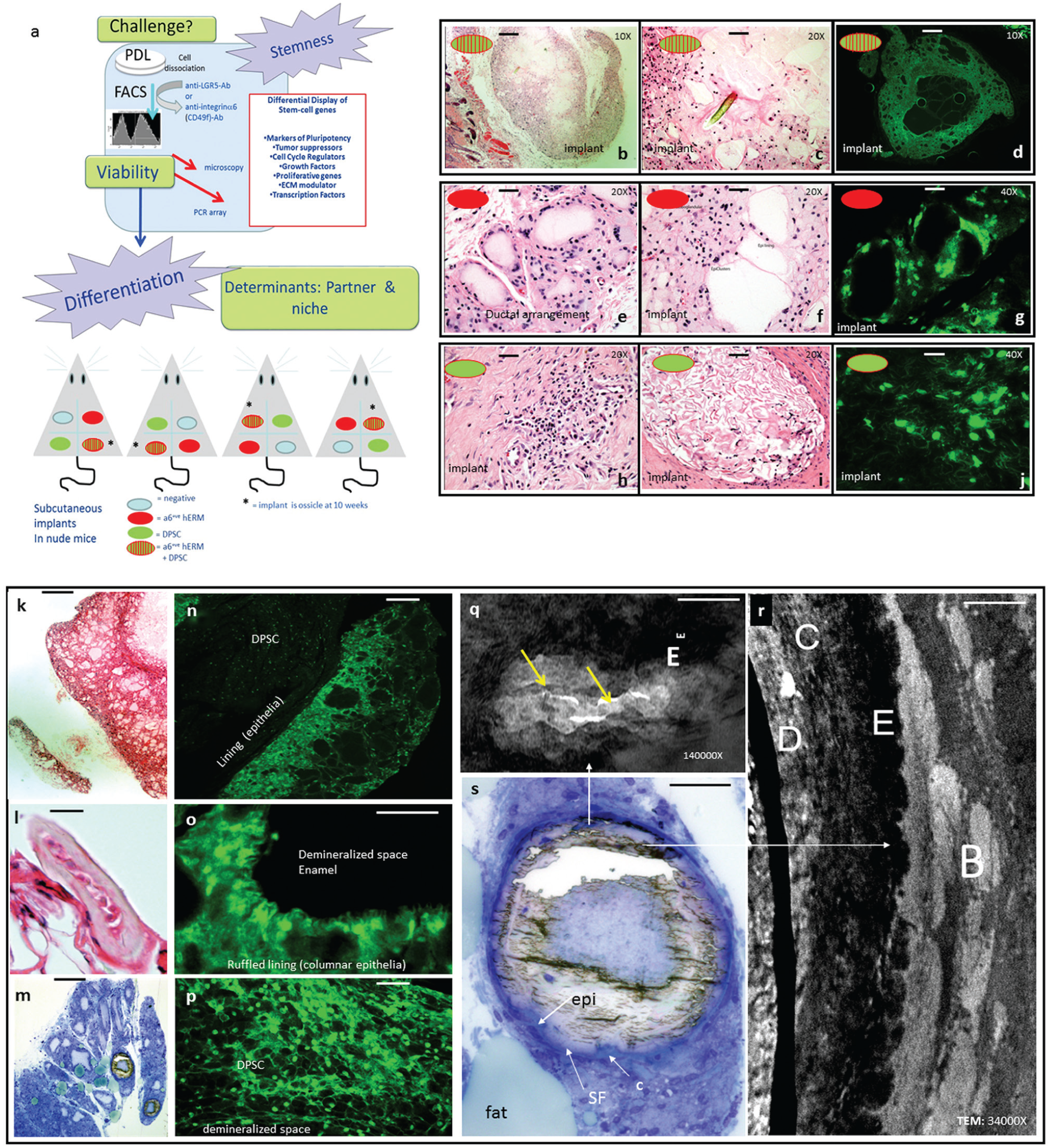
Cotransaplantation of integrin-α6+ve cells and human epithelial stem cells (hEpiSCs) leads to formation of fully mineralized ossicles. (
In Vivo Potency of hERM as hEpiSC: Epithelioid Organelle Formation or Mineralization—A Matter of Cross Signaling
Integrin-α6+ve hEpiSCs (103) were coseeded with an equal number of hDPSCs on PLLA nanoscaffolds, which were implanted into nude mice subcutaneously, 1 in each limb quadrant. Four scaffolds were implanted per mouse: the sample and 3 controls. Control implants in each mouse included an acellular implant, an integrin-α6+ve cell only (Fig. 4e–g), and an hDPSC-only control of 2 time points at 6 and 10 weeks with the 4 scaffolds rotated clockwise on each lumbar quadrant to control for positional effects (Fig. 4a; cotransplants shown on Fig. 4b–d, k–r). Histologic analysis of explants from sacrificed mice at the 6-wk time point (Fig. 4b–j) showed acinar, secretory epithelial-like structures but no mineralized tissue in the hERM-only control (Fig. 4f, g). Staining with anti-HuNu antibody identified the nuclei of human origin cells (Fig. 4d, g, j, n–p). At the 10-wk time point, 4 of 4 coimplants and 0 of 12 controls developed remarkable well-formed ossicles, surrounded by dark blue cartilage when stained with Toluidine blue for histologic analysis (Fig. 4m, s). Histologic analysis of the same explants demineralized for histology (Fig. 4k–s) and stained with anti-HuNu antibody showed columnar human epithelial cells lining the demineralized space (Fig. 4o), where hDPSCs were also found (Fig. 4p). Transmission electron microscopy (TEM) showed mineral depositions characteristic of dentin, cementum, enamel, and bone (Fig. 4r, s) as well as profiles of secretory epithelia secreting mineralized fibers, secretory vesicles, mineralization of areas rich in collagen fibers, acellular spicules within mineralizing tissue, as well as trabecular structures of bone (TEM is the subject of another report). The formation of dentin was expected by the inclusion of hDPSCs. Ameloblasts and cementoblasts respectively derive from the enamel organ (committed to epithelial lineage) and the dental papilla (considered mesenchymal). The formation of these different types of mineralized tissue in heterotopic ossicles is seen here for the first time. Because we consider integrin-α6+ve cells multipotent but committed to an epithelial lineage, we propose that signaling from the integrin-α6+ve cells was required for this route of differentiation. The cementoblast potential of the ERM has also been hypothesized (Nam et al. 2011). Further experimentation is needed to understand the epithelial and mesenchymal crosstalk in tooth regeneration as well as the origin of the fibroblasts around the explants.
Discussion
The PDL is a readily accessible, promising source for human therapy. With the additional incentive that the dental hEpiSC niche remains largely unexplored, the search for hEpiSCs in the ERM becomes particularly exciting.
Teeth are highly mineralized organs resulting from sequential and reciprocal interactions between the oral epithelium and the underlying cranial neural crest–derived mesenchyme (Mitsiadis et al. 2003; Wang et al. 2007), important at each stage of tooth development. The program of hair follicle morphogenesis is similar to that of tooth morphogenesis; the onset of both morphogenetic processes is heralded by the formation of the ectodermal placode. The molecular mechanisms regulating the development of distinct epithelial organs are shared to a great extent (Pispa and Thesleff 2003).
Several studies suggest that the biochemical pathways that lead to differentiation of DPSCs into functional odontoblasts are similar to differentiation pathways of BMSCs into osteoblasts (Huang et al. 2009). The odontogenic potential of hDPSCs has been assessed by a number of organ, explant, and cell culture methods, noting the ability of such cultures to mineralize, at least in part (Gronthos et al. 2000; Sonoyama et al. 2006; Rosa et al. 2012). However, DPSCs alone form sparse and dense calcified nodules and fail to develop lipid-laden adipocytes. In our studies, the combination of hDPSCs with hEpiSCs leads to a fully formed osseous explant with the ultrastructure of cementum, dentin, enamel, and bone, surrounded by cartilage-producing fibroblasts (Fig. 4m, r, s; Appendix, TEM studies). Abundant lipid-laden adipocytic clusters are also uniquely seen. Such results were not seen in our control implants with only hERM or only hDPSCs and have not been reported elsewhere, possibly indicating that a single type of “stem” or progenitor cell is insufficient for tissue regeneration. In contrast, a combination of cells would produce a fully functional organ. Grem1+ve cells, for example, were shown to be required to provide a functioning lamina for intestinal organelles produced by the Lgr5+ve cells (Worthley et al. 2015). We propose that mesenchymal-epithelial signaling is necessary for the full formation of mineralized tissues (also suggested by Zhang et al. 2010). Our results raise the possibility that to exploit the genetic potential of human stem cells to regenerate organized tissues, architectural engineering of the supportive extracellular microenvironment may need to be combined with organized cell printing.
This article describes an examination of the hEpiSC properties of hERM integrin-α6+ve and LGR5+ve cells obtained from human subjects (appropriate use statement in Materials and Methods in Appendix).
We have maintained integrin-α6+ve and LGR5+ve cells in xeno-free and feeder-free conditions, and we believe that our methods and findings are of general interest in work with hEpiSCs or progenitor cells of human origin and have potential in nondental tissue application. Integrin-α6+ve clones show ERM-like growth patterns in 2-dimensional culture and grow organoids in matrigel. They express markers of pluripotency and grow organoids (Fig. 2). LGR5+ve cells, a subpopulation of integrin-α6+ve cells, expressed OCT4, NANOG, MYC, and SOX2, the 4 genes that have been shown sufficient for the creation of induced pluripotent stem cells and stem cell pluripotency. An exclusive marker of dental epithelium in the orofacial cavity, the transcription factor PITX2 showed the highest enrichment, confirming the ERM origin of LGR5+ve cells. Additionally, the CFTR gene and S100B genes, necessary for the differentiation of dental epithelia during enamel tissue formation (Josephsen et al. 2010) and ameloblast differentiation, were highly elevated. LGR5+ve cells also showed suppression of genes associated with a neurogenic phenotype and enrichment of genes that maintain an undifferentiated phenotype and control the cell cycle. Differentiation protocols to assess their tissue regeneration capabilities in vitro as well as in vivo are being evaluated.
Our laboratory focuses on the biology of enamel tissue formation. Our long-term objective is to integrate the use of hEpiSCs within the current efforts for tooth organ regeneration. The potential for tissue regeneration achieved with this report constitutes an advance toward the successful tissue engineering of complex mineralized tissue with the cooperation of mesenchymal-epithelial progenitor cell populations that mediate tooth development. The dental enamel structure cannot be replaced nor repaired, because the cells that synthesize the enamel matrix are lost during tooth eruption. The establishment of a novel therapy to engineer enamel would provide an important therapeutic option for reversal of enamel loss.
Author Contributions
M. Athanassiou-Papaefthymiou, contributed to conception, design, and data analysis, drafted and critically revised the manuscript; P. Papagerakis, contributed to conception, design, and data analysis, critically revised the manuscript; S. Papagerakis, contributed to conception and design, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
Acknowledgements
We are grateful to University of Michigan Core Facilities—specifically to Dotty Sorenson at the Microscopy Imaging Laboratory; Mike, Aaron, and Danielle for flow cytometry; and Ronald Craig at the Department of Pathology for technical advice and help. We thank Drs. X. Ma and P. Ma for poly-
Periodontal ligament cells were explanted from the lower third of human third molars in accordance with guidelines for anonymized discarded material by the University of Michigan Institutional Review Board. All animal work complied with the University Committee on Use and Care of Animals code of ethics for animal research at the University of Michigan.
This research was partially supported by funding from the University of Michigan Cancer Center Support Grant (5P30 CA46592) to S. Papagerakis and start-up funds from the Department of Orthodontics and Pediatric Dentistry for P. Papagerakis.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Additional information and figures on data and methods can be obtained by communication with the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.