
Abstract
The tight junction–based paracellular pathway plays an important role in saliva secretion. Zonula occludens (ZO) proteins are submembranous proteins of tight junction complex; however, their function in salivary epithelium is poorly understood. Here, we found that activation of transient receptor potential vanilloid subtype 1 (TRPV1) by capsaicin increased rat saliva secretion both in vivo and ex vivo. Meanwhile, TRPV1 activation enlarged the width of tight junctions between neighboring acinar cells, increased the paracellular flux of 4-kDa fluorescein isothiocyanate (FITC)-dextran in submandibular gland (SMG) tissues, and decreased transepithelial electric resistance (TER) in SMG-C6 cells. ZO-1, -2, and -3 were distributed principally to the apical lateral region of acinar cells in SMG tissues and continuously encircled the peripheries of SMG-C6 cells in the untreated condition. TRPV1 activation obviously diminished ZO-1 and -2 staining, but not ZO-3 or β-catenin, at the cell-cell contacts ex vivo and in vitro. Moreover, in untreated SMG-C6 cells, ZO-1 and -2 single or double knockdown by small interfering RNA (siRNA) increased the paracellular flux of 4-kDa FITC-dextran. In capsaicin-treated cells, ZO-1 and -2 single or double knockdown abolished, whereas their re-expression restored, the capsaicin-induced increase in paracellular permeability. Furthermore, TRPV1 activation increased RhoA activity, and inhibition of either RhoA or Rho kinase (ROCK) abolished the capsaicin-induced TER decrease as well as ZO-1 and -2 redistribution. These results indicate that ZO-1 and -2 play crucial roles in both basal salivary epithelial barrier function and TRPV1-modulated paracellular transport. RhoA-ROCK signaling pathway is responsible for TRPV1-modulated paracellular permeability as well as ZO-1 and -2 redistribution.
Keywords
Introduction
Tight junctions are localized at the most apical region of lateral membranes between neighboring cells, serving as a rate-limiting structure in regulating paracellular transport of fluid and solutes as well as a key constructor in maintaining the apico-basal epithelial polarity (Baker 2010). Tight junctions are integrated by interactions of transmembrane proteins, such as claudins and occludin, with submembranous proteins, like zonula occludens (ZO) proteins, including ZO-1, -2, and -3 (Shen et al. 2011). ZO proteins link tight junction proteins to the underlying actomyosin cytoskeleton and are speculated to be basic organizers and key modulators of tight junction structure and function (Bauer et al. 2010). In salivary glands, the tight junction–based paracellular pathway plays an important role in saliva secretion (Zhang et al. 2013). To date, ZO-1 has been found in mouse, rat, rabbit, and human salivary glands (Hashizume et al. 2004; Peppi and Ghabriel 2004; Maria et al. 2008; Cong et al. 2012). ZO-1 expression and distribution are altered in labial salivary glands from patients with Sjögren’s syndrome as well as in hyposecretory rabbit submandibular gland (SMG) (Ewert et al. 2010; Cong et al. 2012), suggesting essential roles of ZO proteins in salivary epithelium. However, how ZO proteins regulate saliva secretion and the underlying mechanism are still poorly understood.
Transient receptor potential vanilloid subtype 1 (TRPV1) is a ligand-gated nonselective cation channel that can be activated by heat, acid, and capsaicin (Caterina et al. 1997). TRPV1 was originally found in the nervous system, following discoveries in various nonneuronal tissues, including bladder epithelial cells and keratinocytes (Birder et al. 2001; Inoue et al. 2002). We have reported that TRPV1 is expressed in rat, rabbit, and human SMG, and TRPV1 activation by capsaicin increases saliva secretion partially via enhanced paracellular permeability (Zhang et al. 2006; Ding et al. 2010; Cong et al. 2013). However, the exact role of ZO proteins in TRPV1-mediated salivation and paracellular transport remains unknown.
Therefore, the present study was designed to explore the potential roles of ZO proteins in salivary epithelium and TRPV1-modulated paracellular permeability by using rat SMG and a rat SMG acinar cell line, SMG-C6.
Materials and Methods
Reagents and Antibodies
Zostrix cream with 0.075% capsaicin was obtained from Medicis Pharmaceutical (Scottsdale, AZ, USA). Capsaicin, capsazepine (CPZ), and 4-kDa fluorescein isothiocyanate (FITC)-dextran were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). C3 transferase, Rhotekin-RBD beads, and antibody to RhoA were from Cytoskeleton Inc. (Denver, CO, USA). Y-27632 was obtained from Tocris Bioscience (Bristol, UK). Antibodies to ZO-1, -2, and -3 and occludin were from Life Technologies (Carlsbad, CA, USA). The antibody to β-catenin was from Abmart (Shanghai, China). The antibody to β-actin was from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Measurement of Saliva Secretion
Capsaicin cream (0.075%, 0.2 g) was spread over the skin (2 × 2 cm2) covering the SMG in healthy adult male Sprague-Dawley rats (250 to 300 g) at 9:00 to 11:00 am. Total saliva was collected every 5 min during 20 min before and after capsaicin cream application by putting a dry cotton swab into the mouth of the rat. The cotton swab was preweighed (Weight1) and reweighed after collection (Weight2). Saliva flow rate (mg/min) was calculated as (Weight2 − Weight1)/5 min. All experimental procedures were approved by the Ethics Committee of Animal Research, Peking University Health Science Centre, and complied with the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996). All animal research is reported in accordance with the ARRIVE guidelines (Kilkenny et al. 2010).
Perfusion of Isolated Rat SMG
The SMG was isolated and perfused through a polyethylene cannula placed in the external carotid artery under isoflurane anesthesia (Turner et al. 1996). The main excretory duct was cannulated for saliva collection. Krebs-Ringer-HEPES buffer (KRH, 37 °C, bubbled with 95% O2 and 5% CO2) was perfused to the glands for 30 min, and then different concentrations of capsaicin with or without CPZ were administered into the glands for 10 min. Saliva flow was measured as described by Schirmer’s test (Fox et al. 1985).
Transmission Electron Microscopy
The SMG specimens were fixed and examined with a transmission electron microscope (Hitachi H-7000; Hitachi, Tokyo, Japan). The distances between neighboring tight junctions were measured with ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Cell Culture and RT-PCR
SMG-C6 cells were cultured in Dulbecco’s modified Eagle medium: Nutrient Mixture F-12 (DMEM/F-12) (1:1) as described previously (Quissell et al. 1997). Total RNA was extracted by Trizol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The complementary DNA (cDNA) was prepared from 4 µg of total RNA with Moloney Murine Leukemia Virus (M-MLV) reverse-transcriptase (Promega, Madison, WI, USA). The primer sequences are shown in the Appendix Table. The products were separated by electrophoresis on a 1.5% agarose gel, and DNA bands were visualized by staining with ethidium bromide.
Western Blot Analysis
Proteins extracted from SMG tissues and cells were separated on 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride membranes. The membranes were blocked with 5% nonfat milk, incubated with primary antibodies at 4 °C overnight, and probed with horseradish-peroxidase-conjugated secondary antibodies. The immunoreactive bands were visualized with enhanced chemiluminescence reagent (Pierce, Rockford, IL, USA).
Paracellular Tracer Flux Assay
The rat SMG was removed, cut into small pieces, and placed in a chamber (Costar, Corning, NY, USA) with KRH solution. Next, 4-kDa FITC-dextran (1 mg/mL) was added to KRH solution for 10 min, and then capsaicin was added to the solution. The pictures of acini were caught every 5 s under a confocal microscope (Leica TCS SP5; Leica, Wetzlar, Germany) as described previously (Segawa 1994). For the paracellular tracer flux assay in SMG-C6 cells, 4-kDa FITC-dextran was added to the medium in the basal compartment. After 3 h of incubation, a 200-µL aliquot of the medium was collected from the apical compartment, and the tracer flux was measured as the fluorescence intensity of FITC-dextran using a fluorometer (BioTek, Winooski, VT, USA) (Umeda et al. 2006).
Immunofluorescence Staining
The fixed SMG sections (7 µm thick) and cells were blocked in 1% bovine serum albumin for 30 min and then incubated with primary antibodies at 4 °C overnight, followed by incubation with secondary antibodies at 37 °C for 2 h. Nuclei were labeled with 4′,6-diamidino-2-phenylindole (DAPI). Fluorescence images were captured using a confocal microscope. The fluorescent intensities were averaged by use of ImageJ software.
Preparation of Triton X-100 Insoluble and Soluble Fractions
Cells were lysed in ice-cold buffer (0.5% Triton X-100, 100 mmol/L NaCl, 10 mmol/L Tris-HCl, pH 7.4) at 4 °C for 20 min. Cell lysates were centrifuged at 13,000 × g for 30 min. The soluble supernatants were removed as the Triton X-100 soluble fractions. The remaining insoluble residues were resuspended as the Triton X-100 insoluble fractions (Kawedia et al. 2008).
Knockdown or Re-expression of ZO-1 and -2
SMG-C6 cells were cultured to 80% confluence and transfected with small interfering RNA (siRNA) using MegeTran 1.0 (Origene, Rockville, MD, USA). The potent rat siRNAs of ZO-1 5′-GAAGCUAUAUGAACGGUCUdTdT-3′ and ZO-2 5′-GCAAUAUAUGGCCCUAACAdTdT-3′, as well as the nonspecific control, were synthesized by Sigma-Aldrich Co. The cDNA clones of ZO-1 and -2 were from Addgene (Cambridge, MA, USA). Plasmid transfection was carried out and the cells were collected at least 24 h after transfection (Fanning et al. 2002; Oka et al. 2010).
Measurement of Transepithelial Electric Resistance
Confluent monolayers of SMG-C6 cells were grown in 24-well Transwell chambers (filter pore size: 0.4 µm; area: 0.33 cm2; Costar, NY, USA) for 6 to 7 d. Transepithelial electric resistance (TER) was measured at 37 °C with an epithelial voltohmmeter (EVOM; World Precision Instruments, Sarasota, FL, USA). The background value of the filter (90 Ω) was subtracted, and all measurements were performed in triplicate.
RhoA Pull-Down Assay
RhoA pull-down assay was performed with Rhotekin-RBD beads. Tissues and cells were rapidly lysed in pull-down lysis buffer (50 mmol/L Tris, pH 7.5; 500 mmol/L NaCl; 10 mmol/L MgCl2; and 2% IGEPAL, Sigma-Aldrich Co.) and centrifuged at 10,000 × g at 4 °C for 3 min. Equal amounts of the supernatant were incubated with Rhotekin-RBD beads at 4 °C with rotation for 1 h. The beads were washed and performed by Western blot analysis.
Statistical Analysis
Data are presented as mean ± SD. Statistical analyses were performed by 1-way or 2-way analysis of variance (ANOVA) and followed by Bonferroni’s test using GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA, USA). P < 0.05 was considered statistically significant.
Results
TRPV1 Activation Increases Saliva Secretion and Paracellular Permeability of Rat SMG
To detect the effect of capsaicin on saliva secretion, capsaicin cream was spread over the skin covering the SMG in rats. Capsaicin promoted saliva secretion during 20 min (Fig. 1A). The total amount of saliva within 20 min was significantly increased by application of capsaicin (Fig. 1B).
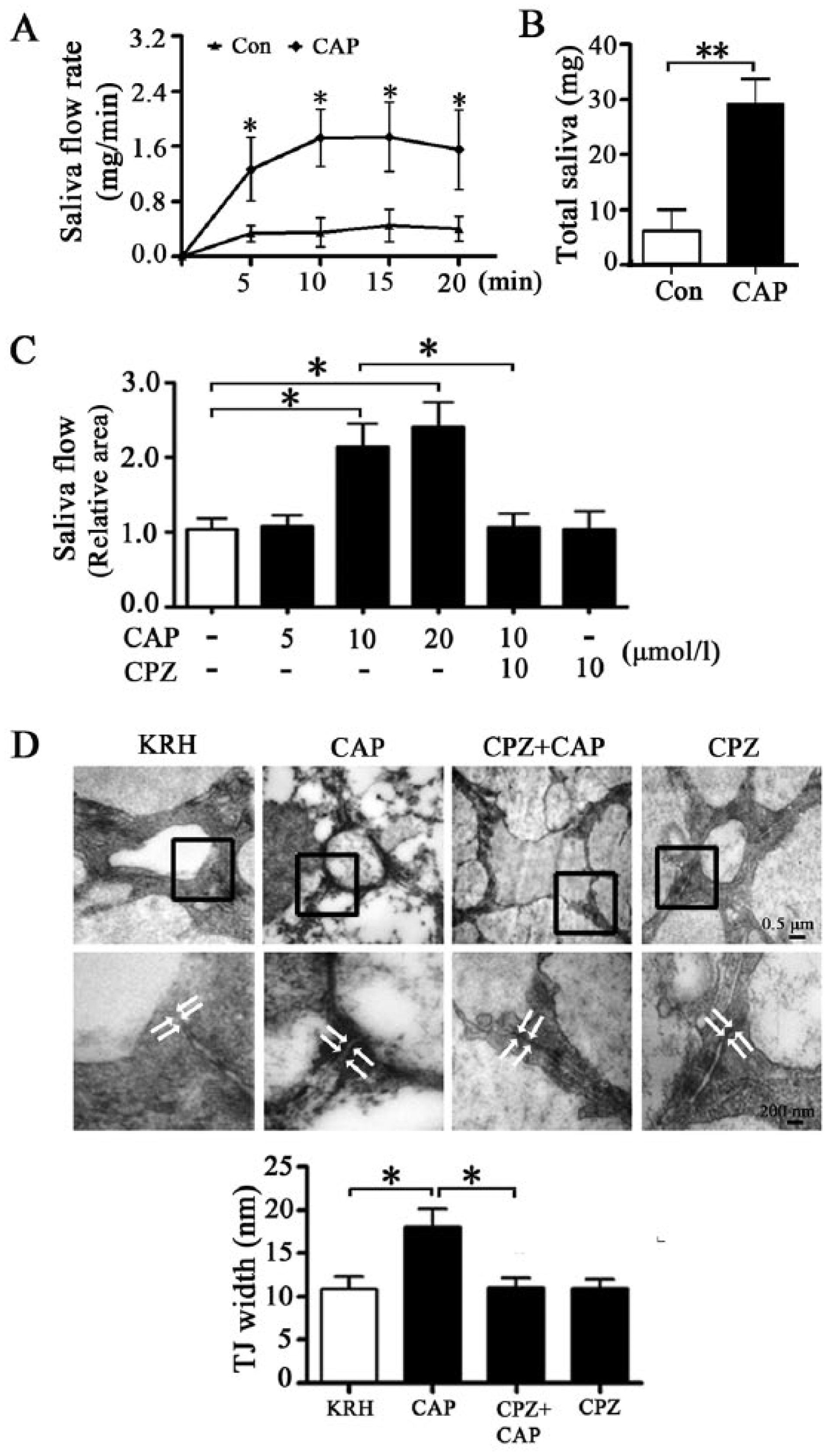
Activation of transient receptor potential vanilloid subtype 1 (TRPV1) increases saliva secretion and tight junction (TJ) width in rat submandibular gland (SMG). Capsaicin (CAP) cream (0.075%, 0.2 g) was spread over the skin (2 × 2 cm2) covering the SMG. Total saliva was collected every 5 min during 20 min before and after CAP cream application by putting a dry cotton swab into the mouth of rats. The cotton swab was preweighed (Weight1) and reweighed after collection (Weight2). (
To explore the direct effect of capsaicin on the function and structure of SMG, we performed ex vivo perfusion into the isolated rat SMG. Capsaicin (10 and 20 µmol/L) significantly increased saliva secretion (Fig. 1C). Preperfusion with CPZ (10 µmol/L), a TRPV1 antagonist, abolished the capsaicin-increased saliva secretion. Tight junctions were located in the apical region of lateral membranes between neighboring acinar cells and formed a slightly dilated distance under transmitted electron microscope. The average width of tight junctions, used as an indicator of tight junction opening, was 10.87 ± 1.48 nm in KRH-perfused glands. Capsaicin enlarged the width of tight junctions, whereas CPZ preperfusion suppressed capsaicin-induced increase (Fig. 1D). The fluorescent intensity of 4-kDa FITC-dextran (1 mg/mL), a tracer frequently used for characterization of paracellular pathway, was increased in the acinar lumen after capsaicin (10 µmol/L) incubation. CPZ inhibited this response (Appendix Fig. 1). These results suggest that TRPV1-mediated salivation might involve an opening of the tight junction–based paracellular route.
TRPV1 Activation Induces the Redistribution of ZO-1 and -2
Expression of ZO-1, -2, and -3 messenger RNA (mRNA) and protein were detected in both rat SMG tissues and SMG-C6 cells (Appendix Fig. 2A, B). Immunofluorescence images revealed that ZO-1, -2, and -3 were localized principally to the apical lateral region in SMG tissues (Appendix Fig. 2C). In SMG-C6 cells, ZO-1, -2, and -3 characteristically formed the continuous staining encircling the cell peripheries and also dispersed in the cytoplasm.
In the capsaicin-perfused SMGs, ZO-1 and -2 stainings appeared discontinuous and were significantly decreased by 48.59% ± 6.86% and 53.43% ± 8.31%, respectively, in the apical lateral region (Fig. 2A), whereas ZO-3 was unaffected. The capsaicin-induced changes of ZO-1 and -2 were inhibited by CPZ pretreatment. Meanwhile, occludin staining was significantly reduced by 50.35% ± 4.04% at the cell-cell contacts after capsaicin perfusion, whereas the distribution of either claudin-3 or β-catenin was unaffected (Fig. 2B). Western blot analysis showed that the total amounts of ZO-1, -2, and -3, occludin, and β-catenin were not altered by capsaicin perfusion (Appendix Fig. 3).
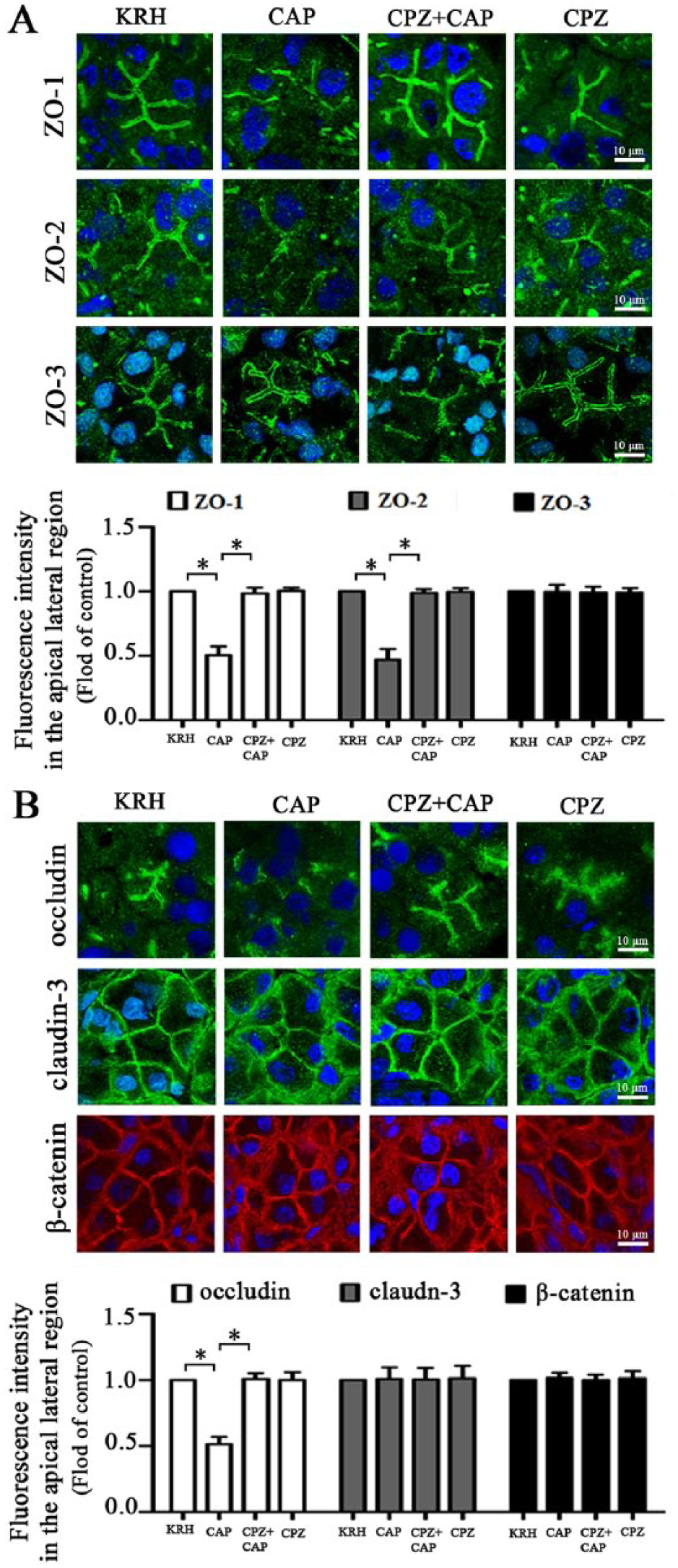
Activation of transient receptor potential vanilloid subtype 1 (TRPV1) induces redistribution of zonula occludens (ZO)-1 and -2, but not ZO-3, in rat submandibular gland (SMG). (
In SMG-C6 cells, the double staining for ZO-1 and occludin from both horizontal (x-y) and vertical (x-z) planes revealed that ZO-1 and occludin were colocalized at the cell borders in untreated cells. Capsaicin (10 µmol/L) caused ZO-1 to partially disappear from cell-cell contacts at 5, 10, and 30 min, and it relocated to the cell borders at 60 min, in parallel with the alteration of occludin (Fig. 3A). Similar changes were observed for ZO-2 (Fig. 3B). The distribution of ZO-3 was unaffected by capsaicin (Fig. 3C, D). Furthermore, the double staining confirmed that TRPV1 activation selectively induced redistribution of ZO-1 and -2 but not β-catenin (Appendix Fig. 4). Western blot analysis showed that the total protein levels of ZO-1, -2, and -3 and β-catenin were not changed by capsaicin treatment within 60 min (Appendix Fig. 5A). However, capsaicin induced ZO-1 and -2 decreases in the Triton X-100 insoluble fractions, concomitant with increases in the soluble fractions at 5, 10, and 30 min, and then recovered to basal levels at 60 min (Appendix Fig. 5B, C), suggesting a rapid and reversible translocation of ZO-1 and -2 from the tight junction complex into the cytoplasm. The levels of ZO-3 and β-catenin in either insoluble or soluble fractions were unaffected.
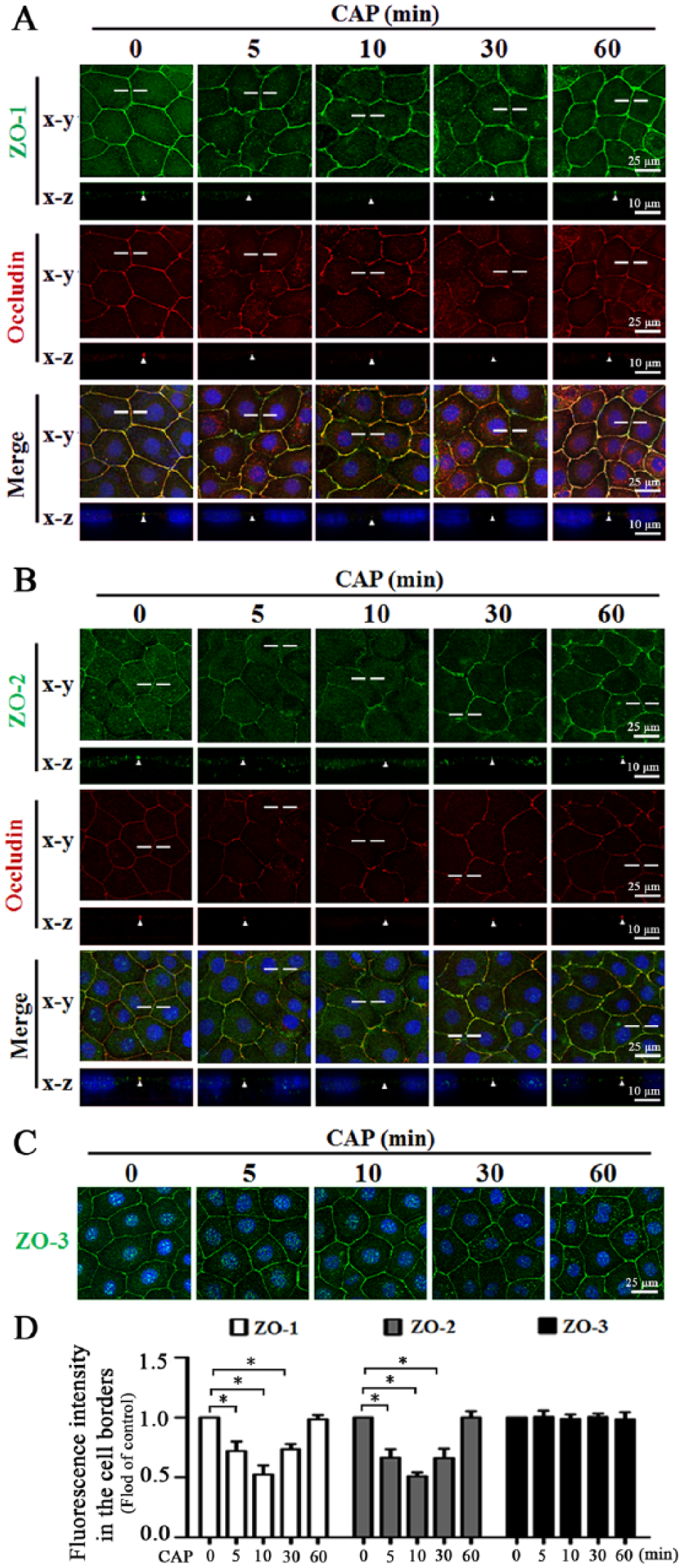
Capsaicin (CAP) induces redistribution of zonula occludens (ZO)-1 and -2, but not ZO-3, in SMG-C6 cells. (
Knockdown of ZO-1 and -2 Alters Paracellular Permeability in SMG-C6 Cells
Next, we examined roles of ZO-1 and -2 in determining the paracellular permeability of SMG-C6. In ZO-1 and -2 single or double knockdown cells, the levels of ZO-3, occludin, and β-catenin were not affected (Fig. 4A–C). There were no significant differences in the basal TER values among ZO-1, ZO-2, ZO-1/-2 knockdown, and scrambled cells (Fig. 4D–F). Capsaicin caused a prominent drop in TER values from 5 to 60 min in scrambled cells, whereas the responses were significantly inhibited in the ZO-1 and -2 single or double knockdown cells. Moreover, the decreased TER reappeared in these corresponding reexpressed (rescue) cells (Fig. 4G–I). Paracellular flux assay revealed that the flux of 4-kDa FITC-dextran was significantly increased in ZO-1, ZO-2, and ZO-1/-2 knockdown cells compared with the scrambled cells, whereas the flux was restored to basal levels in the rescue cells (Fig. 4J–L). Capsaicin significantly increased the tracer flux in the scrambled cells but had no further effect in the knockdown cells. The responses reappeared in the rescue cells.
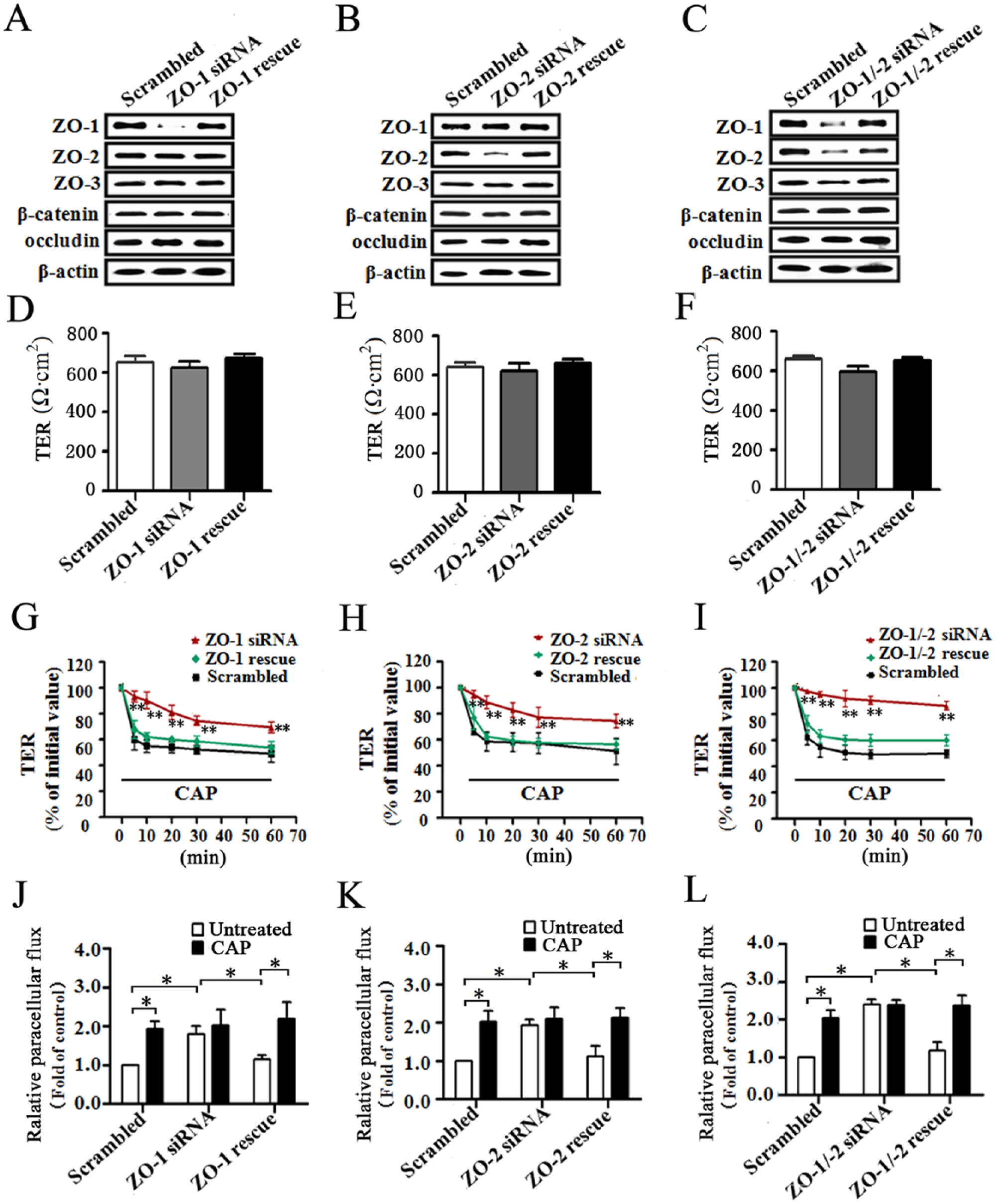
Zonula occludens (ZO)-1 and -2 are required for the capsaicin (CAP)-induced increase in paracellular permeability. (
RhoA-ROCK Signaling Pathway Is Involved in TRPV1-Modulated Paracellular Permeability and Redistribution of ZO-1 and -2
We next explored the possible intracellular signaling pathway. Capsaicin significantly increased RhoA activity in both perfused SMG and cultured SMG-C6 cells (Fig. 5A, B). Preincubation with CPZ blocked the capsaicin-induced increase in RhoA activity ex vivo and in vitro (Fig. 5A, C). Furthermore, pretreatment with either RhoA inhibitor C3 transferase or Rho kinase (ROCK) inhibitor Y27632 suppressed the capsaicin-induced TER decrease (Fig. 5D, E) as well as the translocation of ZO-1 and -2 from Triton X-100 insoluble fractions into soluble fractions (Fig. 5F, G). Immunofluorescence staining further confirmed that the capsaicin-induced ZO-1 and -2 redistribution was suppressed by inhibition of RhoA-ROCK signaling pathway (Fig. 5H).
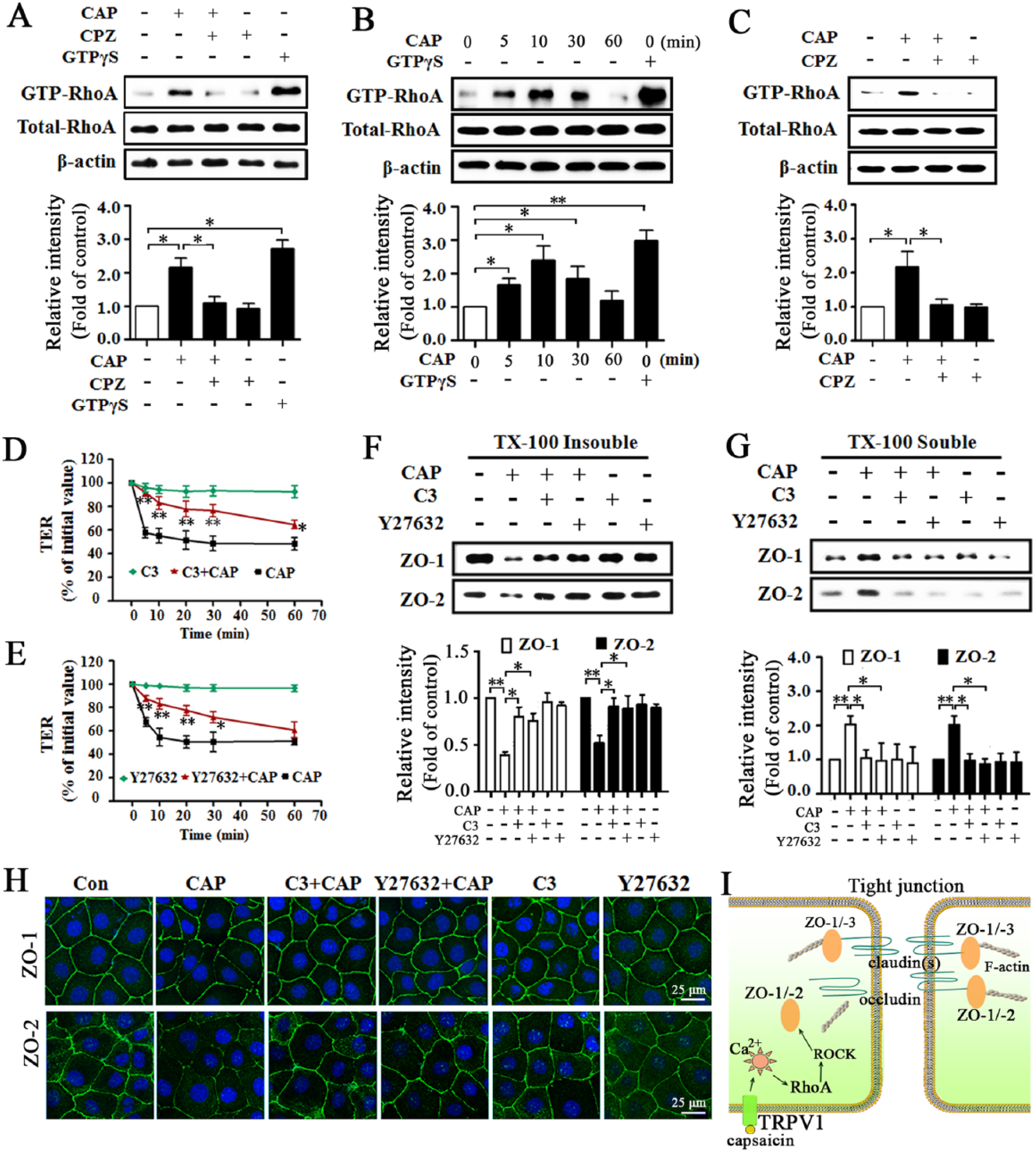
The signaling pathway for RhoA and Rho kinase (ROCK) is involved in transient receptor potential vanilloid subtype 1 (TRPV1)–modulated paracellular permeability as well as redistribution of zonula occludens (ZO)-1 and -2. (
Discussion
In the present study, we demonstrate that ZO-1 and -2 are essential components in tight junction barrier function in rat submandibular epithelial cells. The redistribution of ZO-1 and -2 from tight junctions into the cytoplasm plays a critical role in TRPV1-induced increases in paracellular permeability ex vivo and in vitro. Furthermore, the RhoA-ROCK signaling pathway is responsible for TRPV1-modulated ZO-1 and -2 redistribution. These results reveal an important role of ZO proteins in both basal and TRPV1-modulated tight junction properties and characterize a novel mechanism of TRPV1-modulated paracellular transport in SMG.
ZO proteins play vital roles in tight junction structure and function. Targeted deletion of either ZO-1 or -2 in mice leads to embryonic lethality, whereas ZO-3 deficiency does not show any abnormity (Adachi et al. 2006; Katsuno et al. 2008; Xu et al. 2008). Knockdown of either ZO-1 or -2, but not ZO-3, in cultured epithelial cells leads to delayed formation of tight junctions and an increase in paracellular flux, whereas exogenous expression of either of the two proteins alleviates the mutant phenotype (Adachi et al. 2006; Umeda et al. 2006; Van Itallie et al. 2009). Thus, ZO-1 and -2 have been considered as critical determinants of barrier structure and pivotal regulators of barrier function (Fanning and Anderson 2009). However, few data are available on the function and significance of ZO proteins in salivary epithelium. Here, we characterized that ZO-1, -2, and -3 were mainly localized to the apical lateral region and scattered in the cytoplasm in submandibular epithelial cells. As reported previously, TER reflects that ions pass through the claudin-based charge-selective pores (~4 Å in radius) with instantaneous (typically in seconds) high permeability, whereas paracellular flux assay indicates that large solutes (>4 Å in radius) pass through the so-called “leaky pathway” with lower permeability that is measured over much longer intervals (typically >2 h) (Srinivasan et al. 2015). Here, we showed that the TER values were not different, whereas the flux of 4-kDa FITC-dextran significantly increased in the ZO-1, ZO-2, or ZO-1/-2 knockdown cells compared with the scrambled control cells in untreated condition, and these changes were reversed by reexpression of ZO-1 and/or ZO-2. These results suggest that ZO-1 and -2 are essential elements to maintain the barrier function and that they might exert a specific effect on the permeability of large solutes without an obvious influence on the ionic permeability in normal salivary epithelium. A similar phenomenon has been observed in Eph4 and MDCK cells (Umeda et al. 2006; Fanning et al. 2012).
Next we explored the role of ZO proteins in TRPV1-modulated tight junction barrier function. Activation of TRPV1 markedly increased paracellular permeability of SMG as characterized by increased flux of 4-kDa FITC-dextran and decreased TER. Knockdown of ZO-1 and -2 alone or in combination partially abolished the capsaicin-induced increase in paracellular permeability, whereas their rescue restored this paracellular permeability. These results further indicate a critical requirement of ZO-1 and -2 in TRPV1-modulated tight junction barrier function. Furthermore, the increased paracellular permeability was accompanied by an increased saliva secretion, which suggests that the TRPV1-promoted saliva secretion is mediated, at least in part, by paracellular transport through ZO-1 and -2. As previously reported, application of capsaicin to the apex of the tongue or to the palatal mucosa induces salivation through the parasympathetic cholinergic pathway (Dunér-Engström et al. 1986). We found that capsaicin promotes saliva secretion in vivo and ex vivo in rat, rabbits, and healthy volunteers, indicating that TRPV1 activation can directly stimulate saliva secretion in SMG (Zhang et al. 2006; Cong et al. 2012; Ding et al. 2010).
Numerous studies have shown that changes in the subcellular distribution of ZO proteins in response to various stimuli are often in association with altered tight junction structure and barrier function (Nusrat et al. 2000). For example, acute lung injury induces ZO-1 redistribution accompanied by disruption of barrier integrity in pulmonary epithelium (Jacob and Gaver 2012). Renal ischemia-reperfusion injury causes ZO-1 relocalization followed by disrupted barrier function in rat kidney tissues (Lee et al. 2011). Accordingly, ZO-1 has been recognized as a marker for the integrity of tight junctions (Hurd et al. 2003; Xu et al. 2011). Here, we found that TRPV1 activation selectively induced ZO-1 and -2 redistribution, but not ZO-3 or β-catenin, in both SMG tissues and SMG-C6 cells, suggesting that ZO-1 and -2 are involved in TRPV1-modulated tight junction barrier function by altering their subcellular distribution.
We further explored the signaling pathway linking TRPV1 to ZO proteins. The RhoA-ROCK signaling pathway is considered to play important roles in regulating ZO proteins (González-Mariscal et al. 2008). In Eph4 cells, RhoA activator ARHGEF11 mediates ZO-1-dependent junction assembly and barrier formation (Itoh et al. 2012). In intestinal epithelium, RhoA activity is required for the increased paracellular permeability and relocation of ZO-1 in response to leptin stimulation (Le Dréan et al. 2014). Here, we provide additional evidence that the RhoA-ROCK signaling pathway is involved in modulating tight junction properties of salivary epithelium. TRPV1 activation increased RhoA activity ex vivo and in vitro. Inhibition of the RhoA-ROCK signaling pathway abolished the capsaicin-induced TER decrease as well as redistribution of ZO-1 and -2, indicating that TRPV1-modulated changes in tight junction barrier function and structure are mediated by the RhoA-ROCK signaling pathway.
In summary, we provide the first evidence that ZO-1 and -2 are crucial elements in maintaining tight junction barrier integrity and are key targets in TRPV1-modulated paracellular transport in rat submandibular epithelium. As well, we found that the RhoA-ROCK signaling pathway mediates the TRPV1-induced redistribution of ZO-1 and -2 as well as the increase in paracellular permeability (schematic illustration shown in Fig. 5I). These findings highlight the functional significance of ZO-1 and -2 in salivary epithelial cells and enrich our understanding of the mechanism involved in TRPV1-modulated saliva secretion.
Author Contributions
J. Li, contributed to data acquisition, analysis and interpretation, drafted the manuscript; X. Cong, contributed to data interpretation and analysis, critically revised the manuscript; Y. Zhang, contributed to data interpretation, critically revised the manuscript; R.L. Xiang, contributed to data analysis, critically revised the manuscript; M. Mei, N.Y. Yang, Y.C. Su, S. Choi, L.W. Zhang, contributed to data acquisition, critically revised the manuscript; K. Park, contributed to conception, critically revised the manuscript; L.L. Wu, G.Y. Yu, contributed to conception and design, critically revised the manuscript. All authors have given final approval and agree to be accountable for all aspects of the work.
Footnotes
Acknowledgements
We thank Dr. David O. Quissell from the School of Dentistry, University of Colorado Health Sciences Center, Denver, CO, USA, for the generous gift of rat SMG-C6 cell line and Prof. Guo-He Zhang from the National Center for Advancing Translational Sciences, NIH, Bethesda, MD, USA, for reviewing the manuscript.
This study was supported by the grants from the National Natural Science Foundation of China (Nos. 81271161, 81470756, 81311140269, and 81170974), Doctoral Fund of Ministry of Education of China (No. 20120001110045), and Beijing Natural Science Foundation (No. 7132201).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.