
Abstract
It is known from the paleontology studies of eutherian mammals that incisor numbers were reduced during evolution. The evolutionary lost incisors may remain as vestigial structures at embryonic stages. The recapitulation of the incisor patterns among mammalian species will potentially uncover the mechanisms underlying the phenotypic transition of incisors during evolution. Here, we showed that a minute tooth formed in the presumptive groove region of the gerbil upper incisor at the early developmental stages, during which multiple epithelial swellings and Shh transcription domains spatiotemporally appeared in the dental epithelium, suggests the existence of vestigial dental primordia. Interestingly, when we trimmed the surrounding mesenchyme from incisor tooth germs at or before the bud stage prior to ex vivo culture, the explants developed different incisor phenotypes ranging from triplicated incisors, duplicated incisors, to Lagomorpha-like incisors, corresponding to the incisor patterns in the eutherian mammals. These results imply that the phenotypic transition of incisors during evolution, as well as the achievement of ultimate incisors in adults, arose from differential integrations of primordia. However, when the incisor tooth germ was trimmed at the cap stage, a grooved incisor developed similar to the normal condition. Furthermore, the incisor tooth germ developed a small but smooth incisor after the additional removal of the minute tooth and a lateral rudiment. These results suggest that multiple dental primordia integrated before the cap stage, with the labial primordia contributing to the labial face of the functional incisor. The minute tooth that occupied the boundary of the 2 labial primordia might be implicated in the groove formation. This study sheds light on how rudiments incorporate into functional organs and aids the understanding of incisor evolution.
Introduction
Diversity in incisor patterns plays a crucial role in vertebrate evolution with modification of incisor number, shape, and size allowing an organism to adapt to new diets (Ohazama et al. 2010). The eutherian (placental) mammals contain 3 incisors in each quadrant while marsupial mammals have 4 or 5 incisors (Swindler 2002). During evolution, incisor numbers reduced in most mammal species, with 2 incisors remaining in primates and 1 incisor in rodents (Rose 2009; Charles et al. 2011). Two main proposed mechanisms were formulated to explain the evolutionary reduction of teeth number (Yuan et al. 2008; Cobourne and Sharpe 2010; Peterkova et al. 2014). Peterkova et al. (1993, 2006) proposed that the functional incisors originate from the integration of 3 primordia in the mandible and 5 to 6 primordia in the maxilla based on histological and 3-dimensional (3D) reconstruction analyses. Further molecular analysis detected 2 labial and superficial Shh expressed domains in the vestigial primordia, as well as 1 lingual and deeper Shh expressed domain in the functional incisor (Hovorakova et al. 2013). In contrast, some studies described that the incisor in mouse corresponds to the medial incisor of eutherian dental formula, while the other 2 primordia are lost (Strassburg et al. 1971; Charles et al. 2011). A concentration of apoptosis occurs at the anterior limit of the functional incisor primordium in the lower (Kieffer et al. 1999) and upper (Hovorakova et al. 2013) jaws, relating to the suppression of the rudimentary incisor primordia in both cases (Hovorakova et al. 2011; Hovorakova et al. 2013). Moreover, it has been proven that both integration and regression play roles in the morphogenesis of the first molar in mouse (Prochazka et al. 2010; Lagronova-Churava et al. 2013). Interestingly, both in vitro (Munne et al. 2009) and in vivo (Murashima-Suginami et al. 2007; Ohazama et al. 2008; Charles et al. 2011; Blackburn et al. 2015) studies of incisor development have indicated that the suppressed rudiments can be rescued and revitalized in mice. While both regression and integration of dental primordia have been mentioned in incisor development, the contributions of these structures to the functional incisor and their significance for the incisor evolution have yet to be determined.
Tooth development is regulated by the inductive crosstalk between dental epithelium and mesenchyme and passes through the epithelial thickening, bud, cap, and bell stages, named according to the stage-specific epithelial shapes. At the initiation stage, the transcription of Shh in the thickened epithelium represents odontogenesis (Sarkar et al. 2000; Hovorakova et al. 2013). Prior to the bud stage, the tooth inductive potential resides in the epithelium. Thereafter, the tooth inductive potential shifts to dental mesenchyme (Chen et al. 2000), accompanied by the shifts of Bmp4 expression (Vainio et al. 1993). The Bmp4 expression in the dental mesenchyme was responsible for the induction and maintenance of Shh expression in the dental epithelium (Zhang et al. 2000). Recent studies demonstrated that the dental mesenchyme has the inductive and restrictive potentials to control tooth number through the balance of bone morphogenetic protein (BMP) and Wnt signaling and their antagonists (Murashima-Suginami et al. 2007; Murashima-Suginami et al. 2008; Munne et al. 2009; Munne et al. 2010; Hu et al. 2012).
Analyzing the developmental mechanisms of the tooth in extant mammals beyond the mouse will extend our knowledge about tooth morphogenesis. Here, by analyzing the morphogenesis of the upper incisors in the Mongolian gerbil (subfamily Gerbillinae, genus Meriones), we demonstrated that multiple dental primordia integrate to form the functional incisor. The reindividualization of the gerbil incisor recapitulates the incisor patterns in different orders in mammals. Our results expand the integration model from molar to incisor, providing novel mechanistic insights into incisor development and evolution.
Materials and Methods
All experiments were performed according to the guidelines of the Intramural Animal Use and Care Committee (2013-0415), College of Dentistry, Yonsei University.
Animals
The Institute of Cancer Research (ICR) mice (Koatech Co, Pyeongtaek, Korea), the Mongolian gerbils (maintained at the Department of Oral Biology, Yonsei University), and nude mice (Orientbio, Seongnam, Korea) were used in this study. The day on which a vaginal plug was detected is designated as embryonic day 0.5 (E0.5). The estrus cycle of the female gerbils was assessed by using vaginal smears. The female gerbils were mated during either stage I (preestrus) or stage II (estrus) of the estrus cycle. The following day after mating was counted as E0.5. The mouse embryos were collected on E11.5, E13.5, and E14.5, whereas the gerbil embryos were collected on E18.5, E19.0, E19.5, E20.0, E20.5, E21.5, and E23.5 and postnatal day 0 (PN0). Six-week-old male nude mice were used for kidney transplantation.
Histology
Embryos were collected from pregnant females and fixed in 4% paraformaldehyde. Paraffin sections (4 µm thick) were made and stained with hematoxylin and eosin (H&E) for histological analyses.
Tooth Germ Culture and Kidney Capsule Transplantation
The incisor tooth germs were dissected out from the maxillae of the gerbils at E19.0 and E21.5. After removal of the surrounding dental mesenchyme, the incisor tooth germs were then transplanted into the subcapsular layer of the kidneys in the nude mouse for mineralization. All surgical procedures were performed under anesthesia with intraperitoneal injection of 1 mg/mL Zoletil and 0.04% Rompun (1 mL/kg). After 1 mo, the host mice were sacrificed to obtain the mineralized teeth, which were first photographed before extracting from the surrounding bone. The mineralized teeth were then imaged using micro–computed tomography (CT) or histological staining for further analysis.
Whole-Mount In Situ Hybridization
A digoxigenin (DIG)–labeled probe was transcribed from either Shh or Bmp4 complementary DNA (cDNA) inserted plasmid using a Maxiscript Kit (Ambion, Waltham, MA, USA). Briefly, the specimens were digested with proteinase K (Qiagen, Hilden, Germany) and prehybridized in a buffer containing 50% formamide (AMRESCO, Solon, OH, USA), 5× SSC (WelGENE, Daegu, Korea), 50 µg/mL tRNA (Roche, Mannheim, Germany), 50 µg/mL heparin (Sigma-Aldrich, St. Louis, MO, USA), 2% blocking reagent (Roche), 0.1% CHAPS (Roche), 0.1% Tween 20 (AMRESCO), and 1 mM EDTA (WelGENE) at 70°C for 2 h. Hybridization was performed with a DIG-labeled RNA probe overnight. After incubating with anti-DIG alkaline phosphatase conjugate (Roche) overnight, hybridization signal was visualized by using the BM Purple substrate (Roche).
Micro-CT and 3D Reconstruction
Micro-CT (Skyscan 1076; Skyscan, Antwerp, Belgium) was used to scan the mineralized teeth. The data were digitalized using a frame grabber and then were transmitted to a computer. For 3D reconstructions, images of the frontal serial scanning of the mineralized incisors were imported into the software for reconstruction. To show the 3D reconstructions of the dental epithelium and the domains of Shh expression, serial sections were aligned using Voloom v2.3.0 (München, Germany), and surface rendering was created semi-automatically by using the contour drawing model in Imaris 7.1 (Belfast, UK).
Results
Presumptive Groove Region in the Gerbil Incisor at the Early Developmental Stages
The upper incisors in gerbils have a longitudinal groove on the labial surface (Fig. 1B, C), whereas the incisors in mice have a smooth labial surface (Fig. 1G). Cross sections showed that the groove in the gerbil incisor was caused by an invagination of both enamel and dentin layers toward the dental pulp (Fig. 1D, E), which was not observed in the mice (Fig. 1H). The invagination that happened in the gerbil was obvious on the incisor tip (Fig. 1E, E′) and gradually became unapparent on the cervical end (Fig. 1F, F′). To understand the origin of the groove on the incisor, we compared the development of upper incisors in the gerbil and mouse from initiation stages to PN30. The basic stages of incisor morphogenesis in gerbils and mice are similar, but their timing is different (Fig. 1A). At the initiation stage, although both the gerbil and mouse incisor dental epithelia were thickened (Fig. 1I, L), there were 2 epithelial swellings along the labial-lingual axis in the gerbil on the sagittal view (Fig. 1I; red arrows). The 3D reconstructions of dental epithelium showed that the labial swelling was apparent in the midline of the tooth germ (Fig. 1P; red arrow). At the bud stage, a third epithelial swelling appeared on the distal side of the labial one (Fig. 1Q; red arrow). At the bud and cap stages, an evagination of the mesenchyme toward the dental lamina occurred at the boundary between the medial labial epithelial swelling and the lingual one to form a minute tooth in the gerbil (Fig. 1J, J′, K, K′, Q; black arrows), whereas the labial side epithelium of the mouse incisor was smoothly connected to the vestibule (Fig. 1M, N). A similar structure in mouse could not be found until E15.5 (Hovorakova et al. 2013). The minute tooth in gerbil was maintained at more advanced stages, even at PN0 (Fig. 1O). Three-dimensional reconstructions of the gerbil dental epithelium showed that the minute tooth was located on the midline of the labial surface of the tooth germ at E20.5 (Fig. 1Q), where the groove became visible at E23.5 (Fig. 1R; red arrow) and was apparent at PN30 (Fig. 1S; red arrow).
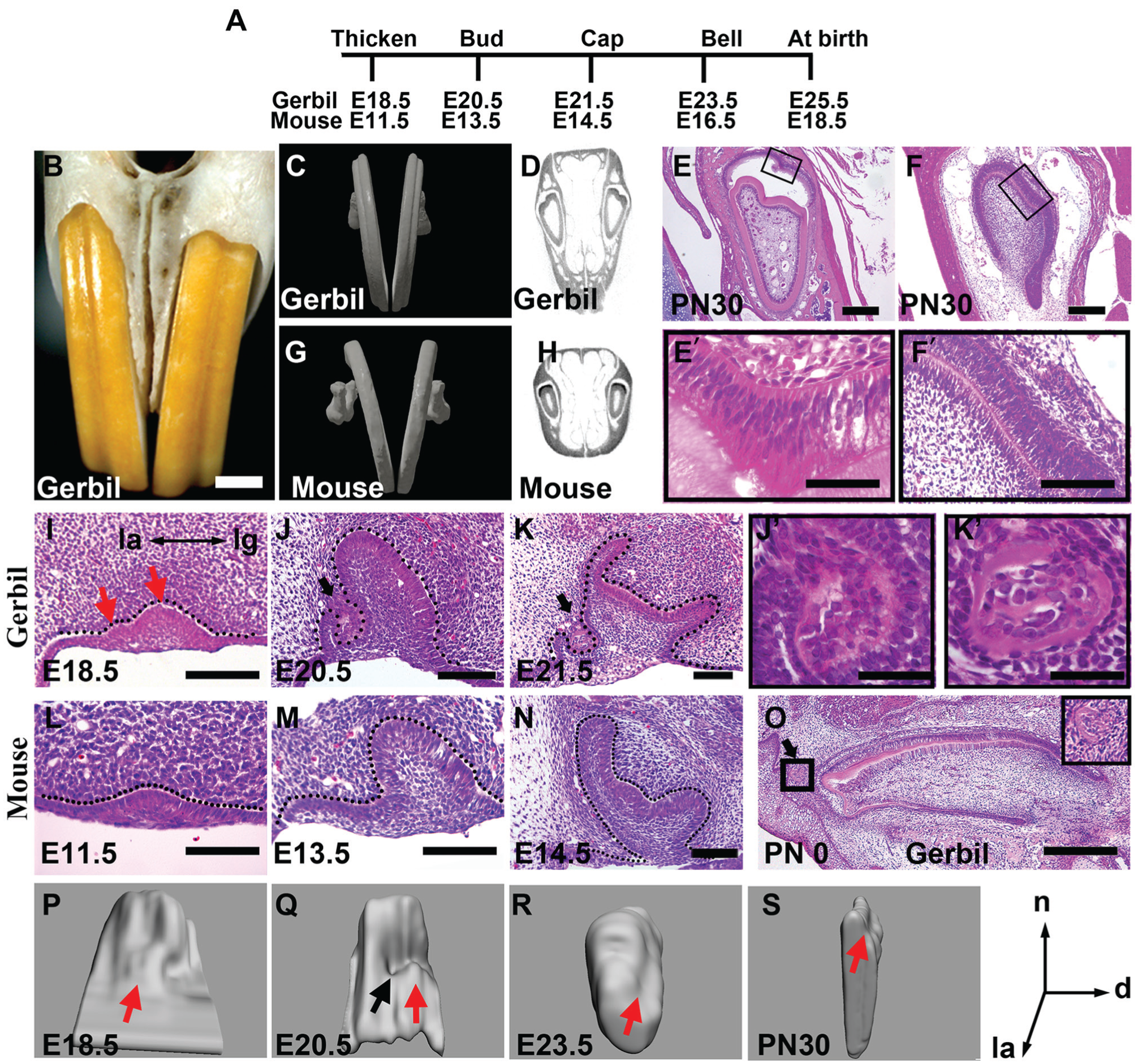
Morphological analysis of upper incisors in gerbil and mouse. (
Multiple Dental Primordia in the Gerbil Incisor at the Early Embryonic Stages
To understand the nature of the minute tooth in the developing incisor of the gerbil, we explored the minute tooth in detail. At E19.5, the newly invasive mesenchymal cells in the minute tooth were condensed compared with the other regions (Fig. 2A, A′). Subsequently, the invasive mesenchyme enriched with dentine-like components at E20.5, and the surrounding basal cells lost cellular polarity (Fig. 2B, B′). Interestingly, compared with the mesenchyme in other regions (Fig. 2H), the condensed mesenchyme in the minute tooth was accompanied by Bmp4 transcription (Fig. 2I). Both the mesenchymal condensation and Bmp4 transcription are evidence of tooth development (Zhang et al. 2005; Mammoto et al. 2011), suggesting the minute tooth is a rudimentary tooth.
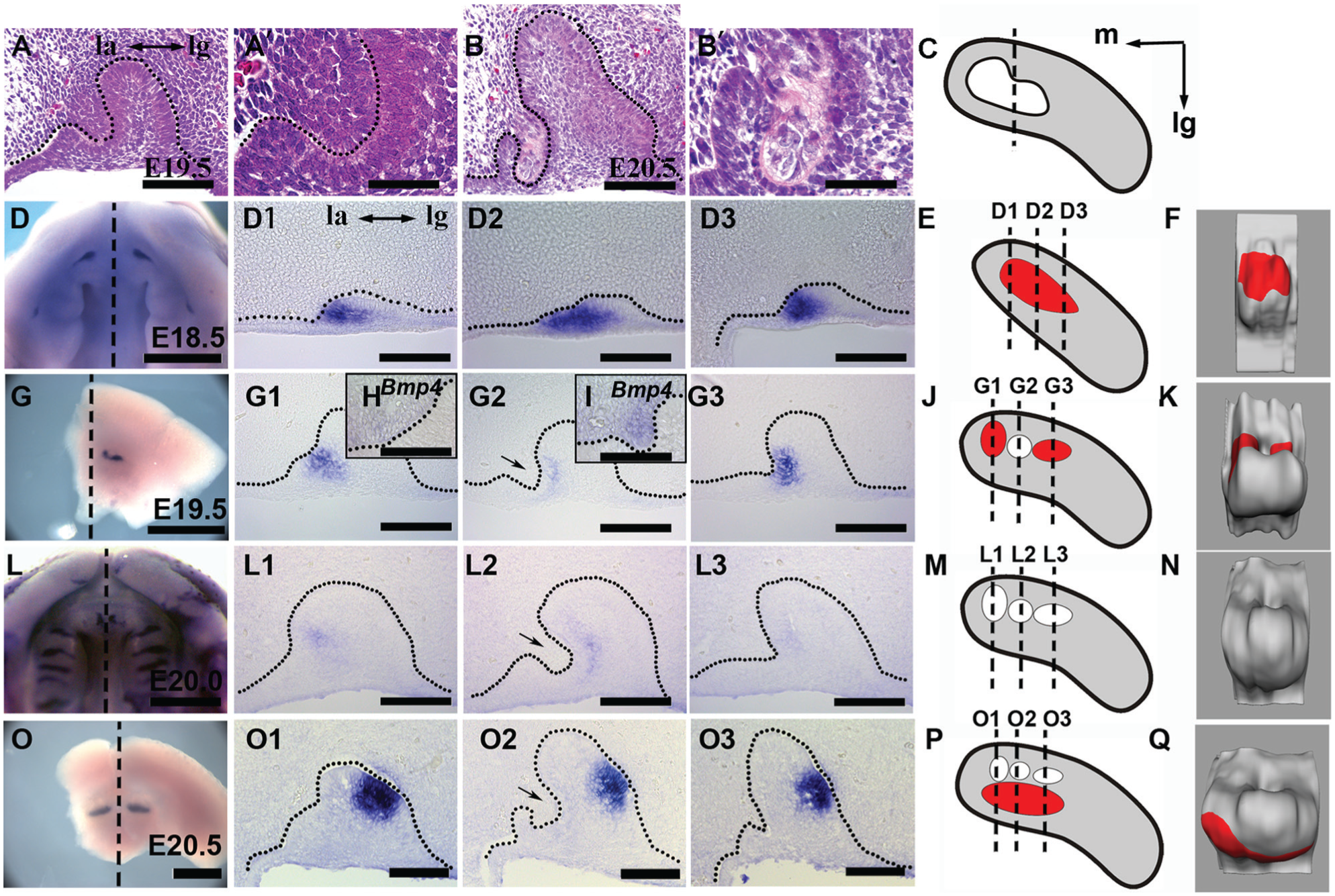
Multiple dental primordia in the gerbil incisor at the early embryonic stages. (
To further study the development of the rudimentary dental primordia in the gerbil upper incisor, we analyzed the expression patterns of Shh, an initial marker for early odontogenesis (Sarkar et al. 2000; Hovorakova et al. 2013). At E18.5, a single strip of Shh transcription domain appeared in the labial dental epithelium in each upper incisor quadrant (Fig. 2D–F), extending along the mesial-distal axis (Fig. 2D1–D3). At E19.5, Shh transcription (Fig. 2G, J, K) maintained in the mesial and distal primordia (Fig. 2G1, G3) but was downregulated in the minute tooth (Fig. 2G2). In more advanced embryos at E20.0, the Shh transcription in the labial dental epithelium became weaker and disappeared (Fig. 2L, L1–L3, M, N). Subsequently, a new large Shh transcription appeared at the lingual tip of the dental epithelium at E20.5 (Fig. 2O, O1–O3, P, Q). Based on all above, we demonstrated that there were 3 distinct Shh transcription domains in each upper incisor in the gerbil at the bud stage: 2 labial ones located medial-distally with the existence of a minute tooth between them (Fig. 2K), as well as a larger one located lingually (Fig. 2Q).
Reindividualization of the Dental Primordia from the Functional Incisor
Typically, extra incisors in the mice could only be gotten from the trimmed explants at or prior to the bud stage (Munne et al. 2009), at a time point before the labial expression of Shh disappeared (Hovorakova et al. 2013), suggesting that the extra incisors may result from the reviving of the rudimentary primordia. A reviving of rudimentary primordia in vitro may shed light on the incisor patterns during evolution. Therefore, we trimmed the mesenchyme of the upper incisor in gerbils before and after the bud stage. The removal of the surrounding mesenchyme of the incisor tooth germ in the gerbil at E19.0 (Fig. 3A) resulted in 3 different phenotypes of incisor: triplicated incisors, Lagomorpha-like incisors, and duplicated incisors. The transverse sections of the triplicated incisors clearly showed that 3 incisors were separated completely but shared the same bone socket (Fig. 3B, F; n = 2/12), indicating that they were developed from 1 incisor tooth germ. The triplicated incisors on the coronal view were situated in an inverted triangular manner, with 2 incisors in the labial side and 1 in the lingual side, similar to the configuration of the grooved incisor developed in vivo (Fig. 3E). The configurations of the triplicated incisors and the grooved incisor are reminiscent of the arrangements of 3 Shh-positive primordia at the embryonic stage. These results imply that the grooved incisor results from the integration of the 2 Shh-positive rudimentary primordia into the lingual functional primordium. The integration was confirmed by the observation of the Lagomorpha-like incisors, which has a tiny incisor located lingual to a grooved incisor, whose size is twice that of the tiny one (Fig. 3C, G, H; n = 2/12). This grooved incisor is likely the result of the incomplete integration of 2 labial Shh-positive primordia, whereas the tiny incisor may develop independently. The duplicated incisors are 2 incisors located abreast without remarkable differences on the transverse size (Fig. 3D, n = 8/12), implying that 1 dental primordium may have been suppressed during the incisor development. Impressively, each revitalized tooth consists of a clearly recognizable crown covered with enamel and a long root that is composed only of dentin (Fig. 3I, J).
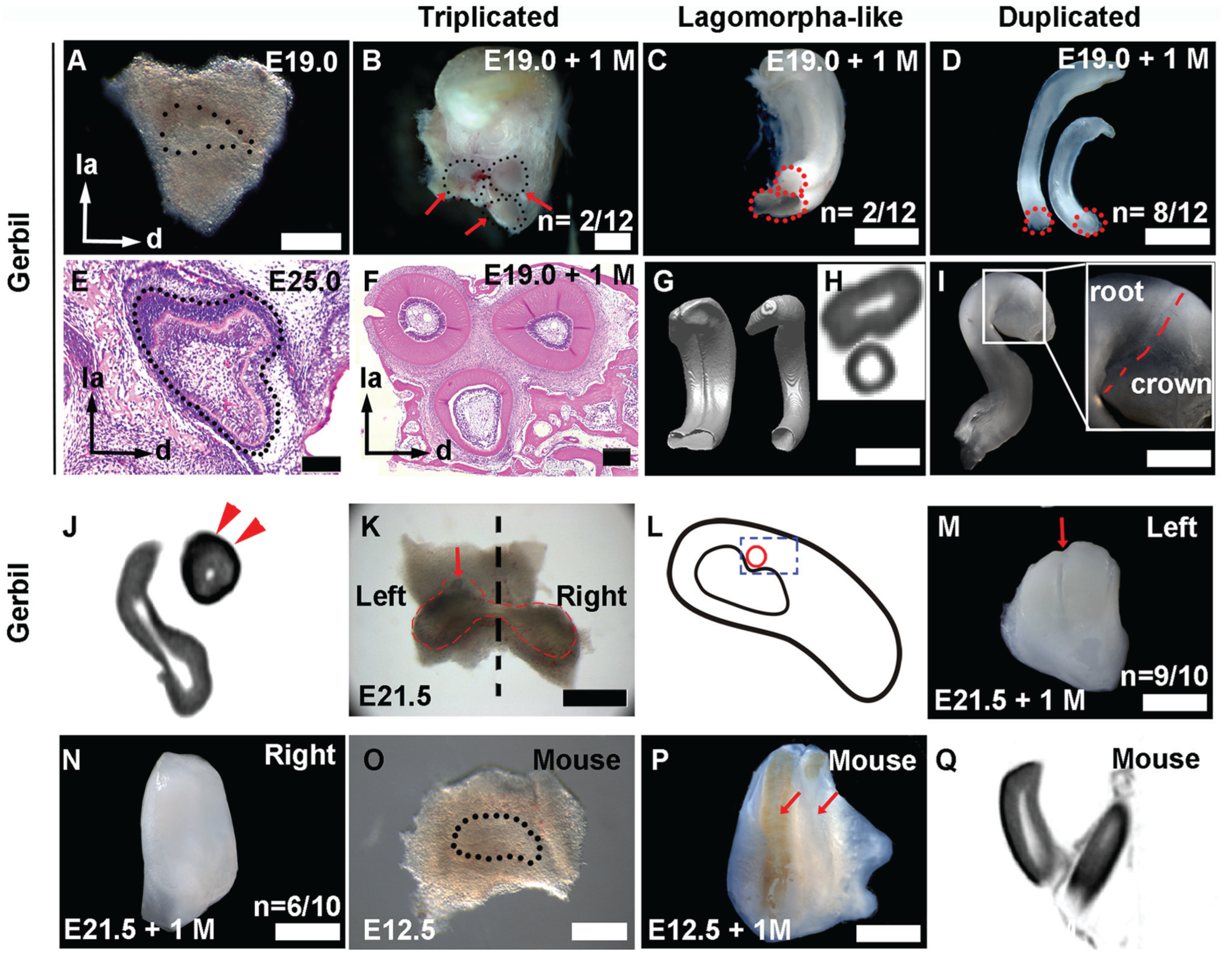
Reindividualization of rudimentary teeth from the maxillary incisor tooth germs. (
In contrast, when dissected the incisor tooth germs at the cap stage, a stage when the labially expressed Shh domain disappeared, most of the explants (Fig. 3K, left) developed into typical grooved incisors of the gerbil (Fig. 3M; n = 9/10), losing the reindividualization capacity. This result suggests that the integration of the dental primordia occurred before the cap stage in vivo. To investigate the relationship between the minute tooth and the groove formation, we also removed the minute tooth and the labial distal rudimentary primordium from the incisor tooth germ at E21.5 (Fig. 3K, right), as indicated by a schematic diagram (Fig. 3L). Impressively, the right incisor tooth germ developed into an incisor without a groove (Fig. 3N, n = 6/10). The above results indicate that the labial face of the functional incisor is contributed by the integration of mesial and distal rudimentary primordia, as their boundary was occupied by the minute tooth at the bud stage, resulting in the formation of a groove.
Compared with the observation in mouse (Hovorakova et al. 2013), the 2 Shh-expressed domains in gerbil were separated by the Shh-negative minute tooth. To test whether this subtle difference has effects on teeth revitalization, we trimmed the incisor tooth germ of the mouse at E12.0 (Fig. 3O). Most of the explants developed into duplicated incisors (Fig. 3P, Q; n = 9/10) after ex vivo culture, indicating that at least 2 dental primordia integrate to form a smooth incisor in the mouse upper jaw.
Discussion
During the development of the gerbil upper incisor, 3 epithelial swellings and 3 Shh transcription domains represent multiple dental primordia (Fig. 4A), similar to the observations in the mouse (Hovorakova et al. 2013). One distinctive difference in the incisor development between gerbil and mouse is the existence of the minute tooth in the gerbil incisor from the bud stage. A morphologically similar structure, called a “deciduous incisor” (Fitzgerald 1973) or “prelacteal tooth” (Hovorakova et al. 2013) in mouse, does not appear as late as E15.5. In this study, we found mesenchymal condensation, Bmp4 transcription, and Shh disappearance in the minute tooth, evidencing a vestige of the tooth. In addition, we observed that there were 2 Shh transcriptional patches adjacent to the minute tooth. Above all, we demonstrated that there are 3 vestigial primordia and 1 functional primordium in each upper quadrant of the gerbil. This finding is similar to the observation that at least 4 primordia integrate to form the incisor bud (Peterkova et al. 1993), and 2 labial Shh-expressed projections form a notch to accommodate the prelacteal tooth in the mouse (Hovorakova et al. 2013; Peterkova et al. 2014).
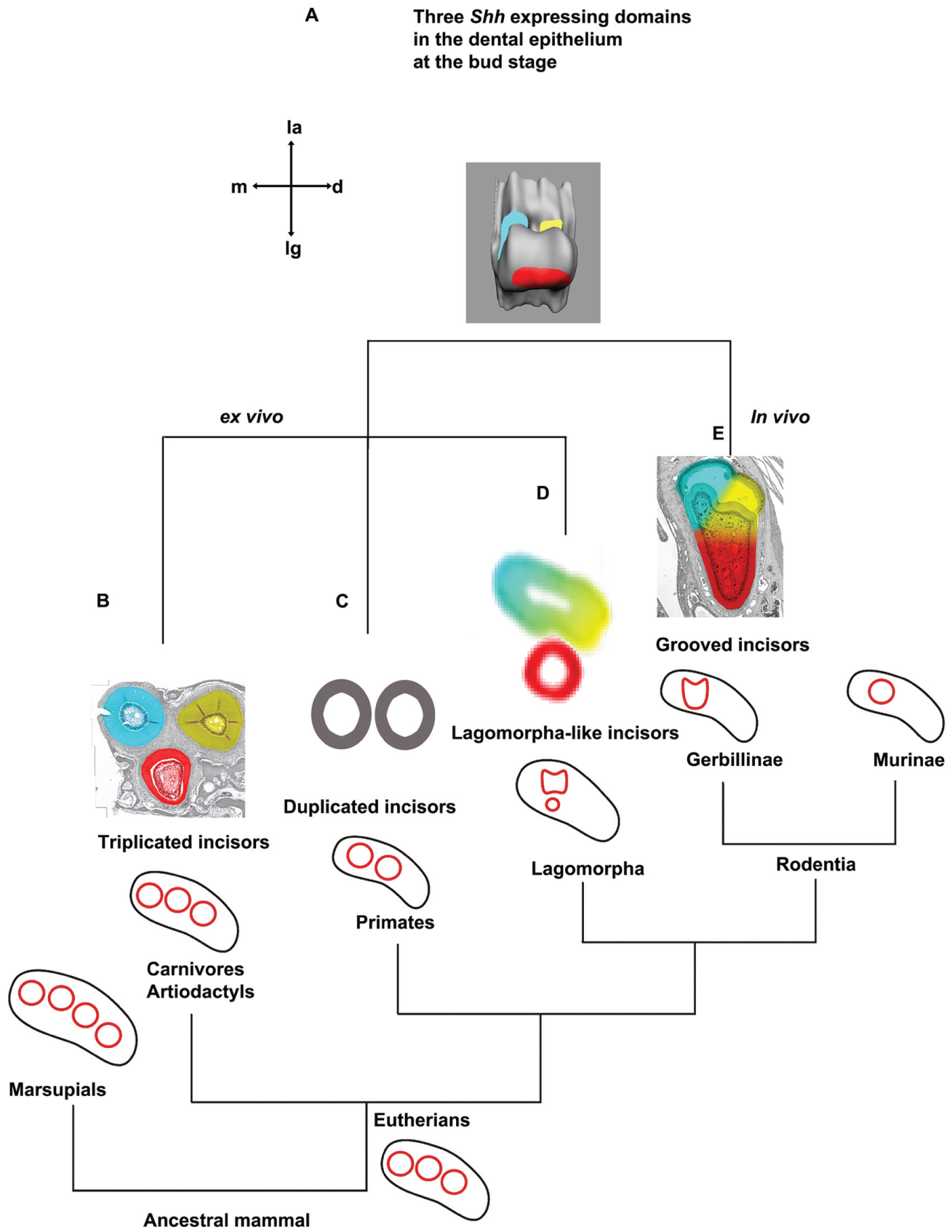
Various phenotypes of incisors produced from 1 gerbil tooth germ recapitulate the upper incisor patterns in the phylogenetic tree of mammals. (
We revitalized the rudimentary dental primordia in the gerbil by minimizing the surrounding mesenchyme. The revitalized incisor teeth from a single tooth germ have 3 phenotypes. The first type is triplicated incisors (Fig. 4B), in a configuration that resembles the shape of grooved incisors in vivo (Fig. 4E). The second type is duplicated incisors (Fig. 4C), which have been commonly observed in mice with a single gene mutation in Sostdc1, Lrp4, or Sprouty (Munne et al. 2009; Ohazama 2010; Charles et al. 2011). The third type is Lagomorpha-like incisors (Fig. 4D). The 3 phenotypes correspond to incisor patterns in the ancestral Placentals (Rose 2009) (Fig. 4B), the orders of Primates (Swindler 2002) (Fig. 4C), and Lagomorpha (Long 2008) (Fig. 4D), respectively. These results highlight how multiple dental primordia are integrated and participate in the morphogenesis of functional incisors, as well as aid in understanding incisor evolution.
Several mechanisms have been used to interpret the supernumerary incisors in mice: ectopic Wnt activity in the mesenchyme (Ohazama et al. 2008), development of a replacement tooth (Munne et al. 2009), splitting of the incisor primordium (Charles et al. 2011), and failure of integration of the ancestral dental primordia (Hovorakova et al. 2011). Our reindividualization experiments supported the integration mechanism, similar to the integration involved in the first molar formation in mouse (Prochazka et al. 2010). Various phenotypes generated in the trimming experiments could be attributed to the different activities of BMP and Wnt signaling in the dental mesenchyme, which can cause the fusion or separation of the incisors in the mouse (Murashima-Suginami et al. 2007; Munne et al. 2010; Hu et al. 2012; Yuan et al. 2015).
Furthermore, in β-catΔPrx/lacZ mice, the Shh-expressed domain split at E13.0 and the incisor morphologically duplicated at E13.5 (Fujimori et al. 2010). Accordingly, our trimmed incisor tooth germs of gerbils could be individualized only before the cap stage, consistent with the observation in mice (Munne et al. 2009). All of these emphasize that the stages before the cap stage are crucial for integration. We therefore proposed that the minute tooth emerged at the bud stage and is prone to preventing the complete integration of labial dental primordia. In the case of the mouse, the minute tooth appeared at E15.5, after the integration of the dental primordia, and thus may not affect the integration process. Nevertheless, the boundary of the dental primordia may still be susceptible, resulting in the groove that was observed in the mutants or the old wild-type mice (Ohazama et al. 2010).
Impressively, the reindividualized incisors from gerbils deposited enamel on the incisor tips, different from the typical enamel pattern in Rodentia but similar to the pattern in the order of Primates. Interestingly, Sprouty mutant mice maintain enamel on the labial surface of the supernumerary incisors (Charles et al. 2011), while other mice with a genetic alteration of Wnt or BMP signaling lack enamel on the supernumerary incisors (Klopcic et al. 2007; Munne et al. 2009; Blackburn et al. 2015). In our trimmed experiments, the removal of the tooth germ–surrounding mesenchyme might have caused the disruption of the antagonistic actions between the mesenchymal BMP4 and activin, which underlie the asymmetric distribution of the enamel between the incisor labial and lingual surfaces (Wang et al. 2004).
The gradient reindividualization of the incisor might correspond to a reversal of the integration of the rudimentary dental primordia that occurred during evolution. The grooved incisor is probably an intermediary step in the evolution of multiple incisors to a single incisor. The evidence we provided in this study will aid in our understanding of dental anomalies in a number of human syndromes and can be applied in modeling controlled tooth regeneration.
Author Contributions
L. Li, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; Q. Tang, contributed to conception, design, data analysis, and interpretation, drafted and critically revised the manuscript; H.-S. Jung, contributed to conception and design, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
Acknowledgements
We thank Zhaoming Wu (HKU) for revising the manuscript.
This research was supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (No. HI14C3266); by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2014R1A2A1A11050764); and by the Bio & Medical Technology Development Program of the NRF funded by the Korean government (MSIP) (No. 2012M3A9B4028738).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.