
Abstract
Dentin-pulp regeneration is closely linked to the presence of nerve fibers in the pulp and to the healing mechanism by sprouting of the nerve fiber’s terminal branches beneath the carious injury site. However, little is known about the initial mechanisms regulating this process in carious teeth. It has been recently demonstrated that the complement system activation, which is one of the first immune responses, contributes to tissue regeneration through the local production of anaphylatoxins such as C5a. While few pulp fibroblasts in intact teeth and in untreated fibroblast cultures express the C5a receptor (C5aR), here we show that all dental pulp fibroblasts, localized beneath the carious injury site, do express this receptor. This observation is consistent with our in vitro results, which showed expression of C5aR in lipoteichoic acid–stimulated pulp fibroblasts. The interaction of C5a, produced after complement synthesis and activation from pulp fibroblasts, with the C5aR of these cells mediated the local brain-derived neurotropic factor (BDNF) secretion. Overall, this activation guided the neuronal growth toward the lipoteichoic acid–stimulated fibroblasts. Thus, our findings highlight a new mechanism in one of the initial steps of the dentin-pulp regeneration process, linking pulp fibroblasts to the nerve sprouting through the complement system activation. This may provide a useful future therapeutic tool in targeting the fibroblasts in the dentin-pulp regeneration process.
Introduction
The success of dentin-pulp regeneration is directly correlated to the presence of nerve fibers in the dentin-pulp complex and to the sprouting of their terminal branches into the surviving pulp beneath the injured site (Kimberly and Byers 1988; Arai 1991). This nerve sprouting can enhance inflammation and healing processes by immune cell recruitment and vascular permeability at the injured site (Byers and Nähri 1999; Hanoun et al. 2015). It has long been established that tissue regeneration is closely linked to inflammation, as demonstrated by a significant tissue regeneration inhibition when corticosteroids are used in case of myocardial infarction (Kloner et al. 1978). In accordance with these data, it has been demonstrated that dentin-pulp regeneration is directly associated with the presence of nerve fibers in pulp tissue since an occlusal exposure induces more pulp necrosis in denervated than innervated teeth (Byers and Taylor 1993). The duration and extent of dental nerve sprouting depend on the nature and intensity of the injury and seem to be regulated by the spatiotemporal modulation of growth factor expression in the pulp (Taylor et al. 1988; Woodnutt et al. 2000). However, although dental nerve sprouting appears to have a critical importance in dentin-pulp regeneration, very little is known about initial molecular events controlling this process.
Recently, it has been demonstrated that the complement system, a major component of innate immunity and inflammation, is activated at the injured site of carious teeth (Chmilewsky, Jeanneau, Laurent, et al. 2014). While complement is known to be activated from plasma proteins, it is also efficiently activated in human teeth from proteins produced by pulp fibroblasts (Chmilewsky, Jeanneau, Laurent, et al. 2014). In addition, it became evident that complement, especially through the active fragment C5a, participates in tissue regeneration, as demonstrated in the liver (Mastellos et al. 2011), bone (Ignatius et al. 2011), teeth (Chmilewsky et al. 2013; Chmilewsky, Jeanneau, Laurent, et al. 2014), and cardiac tissues (Lara-Astiaso et al. 2012). Interestingly, few in vivo and in vitro studies demonstrated that C5a, through its interaction with the C5a receptor (C5aR), is directly involved in the modulation of several growth factors expression. Thus, in a retinal fibrosis mouse model, C5a-C5aR interaction induced a significant production of platelet-derived growth factor and transforming growth factor β (Boor et al. 2007). Also, it has been demonstrated that C5a stimulates the expression of vascular endothelial growth factor by retinal Müller cells in vitro (Cheng et al. 2013). Based on these data, we hypothesized that the complement activation through C5aR could be one of the initial signals in dental nerve sprouting under carious decays.
Materials and Methods
Materials
See Appendix.
Molar Collection
Human immature third molars, freshly extracted for orthodontics reasons, and carious teeth were obtained in compliance with French legislation (informed patients’ consent and institutional review board approval of the protocol used). Teeth were fixed and routinely processed as previously described (Téclès et al. 2005).
Immunohistochemistry
Before tooth sections were stained, antigen retrieval and saturation were performed as previously described (Chmilewsky, Jeanneau, Laurent, et al. 2014). Then tooth sections were incubated for 1 h with anti-C5aR (20 µg/mL), anti–β-III-tubulin (1 µg/mL), or their respective isotype control.
C5aR immunostaining was done by incubating sections for 45 min with Alexa Fluor 488 anti-mouse immunoglobulin G (IgG) (2 µg/mL) and 4′,6-diamidino-2-phenylindole (DAPI) (1 µg/mL) counterstain.
β-III-tubulin immunostaining was done by incubating sections for 2 h with horseradish peroxidase (HRP)–conjugated anti-rabbit (1:200). Diaminobenzidine (DAB, 0.5 mg/mL) was used to visualize the reaction.
Some sections were stained with hematoxylin-eosin.
Cell Cultures
Human pulp cells were prepared from immature third molars at the 2/3-root formation stage by the explant outgrowth method and cultured as previously described (About et al. 2000). Human neurons (HNs) were cultured in neuronal medium supplemented with neuronal growth supplement, 100 µg/mL streptomycin, and 100 U/mL penicillin.
Immunofluorescence Staining
Pulp cells were seeded overnight at 12.5 × 103 cells/cm2 on 8-well culture chambers before incubation in serum-free medium ± lipoteichoic acid (LTA) (1 µg/mL). After 6- or 24-h incubation, cells were fixed with 4% paraformaldehyde (10 min), permeabilized with phosphate-buffered saline (PBS)/0.3% Tween-20/0.3% glycine (30 min), and saturated with 3% bovine serum albumin (BSA) (1 h). Then, cells were incubated 1 h with a mix of anti-C5aR (10 µg/mL) + anti–fibroblast surface protein (FSP) (20 µg/mL), anti-C5aR (5 µg/mL) + anti–phospho-C5aR(pSer338) (10 µg/mL), or their respective control isotypes. Finally, cells were incubated for 40 min with a mix of Alexa Fluor 594 anti-mouse IgG, Alexa Fluor 488 anti-rabbit IgG (2 µg/mL), and DAPI (1 µg/mL). The coverslips were sealed and photographs taken (DMI6000 B microscope; Leica).
In-Cell Western Assay
Pulp cells were seeded at 15 × 103 cells/cm2 in 96-well plates. At subconfluency, cells were incubated in serum-free medium ± LTA (1 µg/mL) and ± W54011 (10 nmol/L) for 24 h. Then, cells were immediately fixed with 100% cold methanol (15 min) and saturated with 5% BSA (1.5 h). Cells were incubated overnight at 4°C with anti-C5aR (5 µg/mL), anti–phospho-C5aR(pSer338) (10 µg/mL), or their respective control isotypes. Cells were then washed (0.05% Tween-20/PBS) and incubated with respective IRDye-680RD secondary antibody (1 h). After 5 washes, plates were scanned at 700 nm (Odyssey CLx).
Neurotrophin Quantifications
Pulp cells were seeded at 15 × 103 cells/cm2 in 12-well plates. At subconfluency, cells were incubated in serum-free medium (300 µL) ± LTA (1 µg/mL) and ± W54011 (10 nmol/L). After 6, 24, and 48 h, brain-derived neurotrophic factor (BDNF) concentrations in supernatant were determined by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer’s instructions. Results were normalized to the cell viability (determined by the 3-[4, 5-dimethylthiazolyl-2]-2, 5-diphenyltetrazolium bromide [MTT] assay as described in Chmilewsky, Jeanneau, Laurent, et al. 2014).
Neurite Outgrowth Assay
Neurite outgrowth was investigated using AXon Investigation Systems (AXIS), as described (Fig. 1; for detailed protocol, see Appendix).
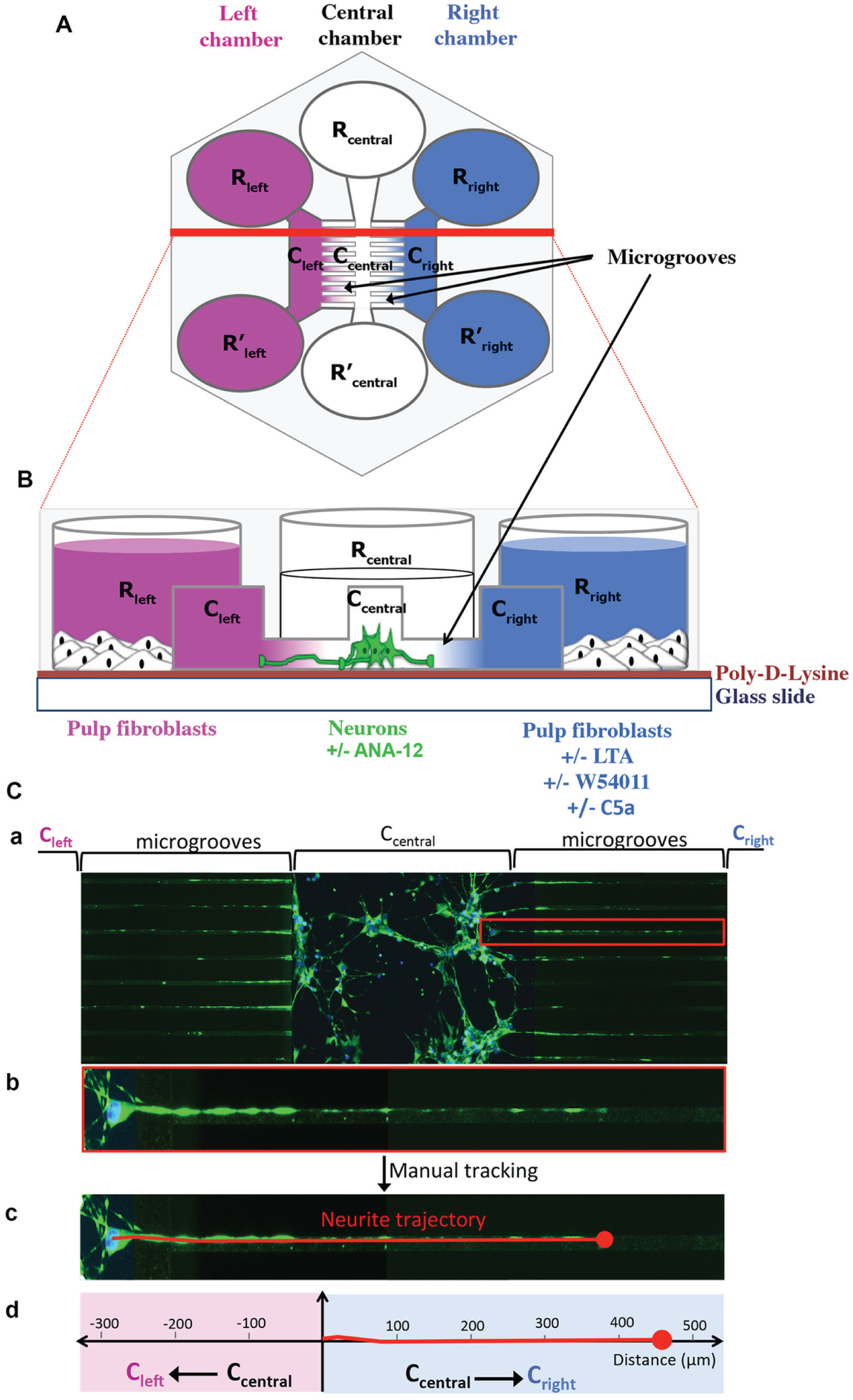
Schematic representations of an Axon Investigation System (AXIS). (
Statistical Analysis
All experiments were repeated at least 3 times (n = 3), and statistical significance was determined using the Student’s t test. Data were expressed as means ± SD and considered significant for P < 0.05.
Results
C5aR Is Expressed in Carious Teeth and Colocalized with Nerve Terminal Endings
We first investigated the expression of C5aR both in intact (Fig. 2A) and carious tooth sections (Fig. 2B). While few or no C5aR expression was detected in intact tooth sections (Fig. 2C, D), an intense staining was specifically observed in the inflamed pulp beneath the carious site (Fig. 2E, F). A higher magnification reveals that all pulp cells localized beneath the injured site expressed C5aR (Fig. 2F) while very little or no C5aR expression was detected in pulp far from injury site (Fig. 2G, H). Nerve fiber localization was investigated on the same tooth sections through β-III-tubulin staining, a differentiated neuron marker (Tao et al. 2015). In intact teeth, β-III-tubulin was mainly localized beneath the odontoblast cell layer (Fig. 2I, J). In carious teeth, a densification of nerve extensions was detected beneath the injured site (Fig. 2K). Indeed, an intense staining characteristic of transverse and longitudinal sectioned nerve structures was clearly observed at higher magnifications (Fig. 2L). Labeling at a distance from the carious site was similar to that in intact teeth (Fig. 2M, N). These results show that C5aR expression is correlated to nerve fiber sprouting in carious teeth.
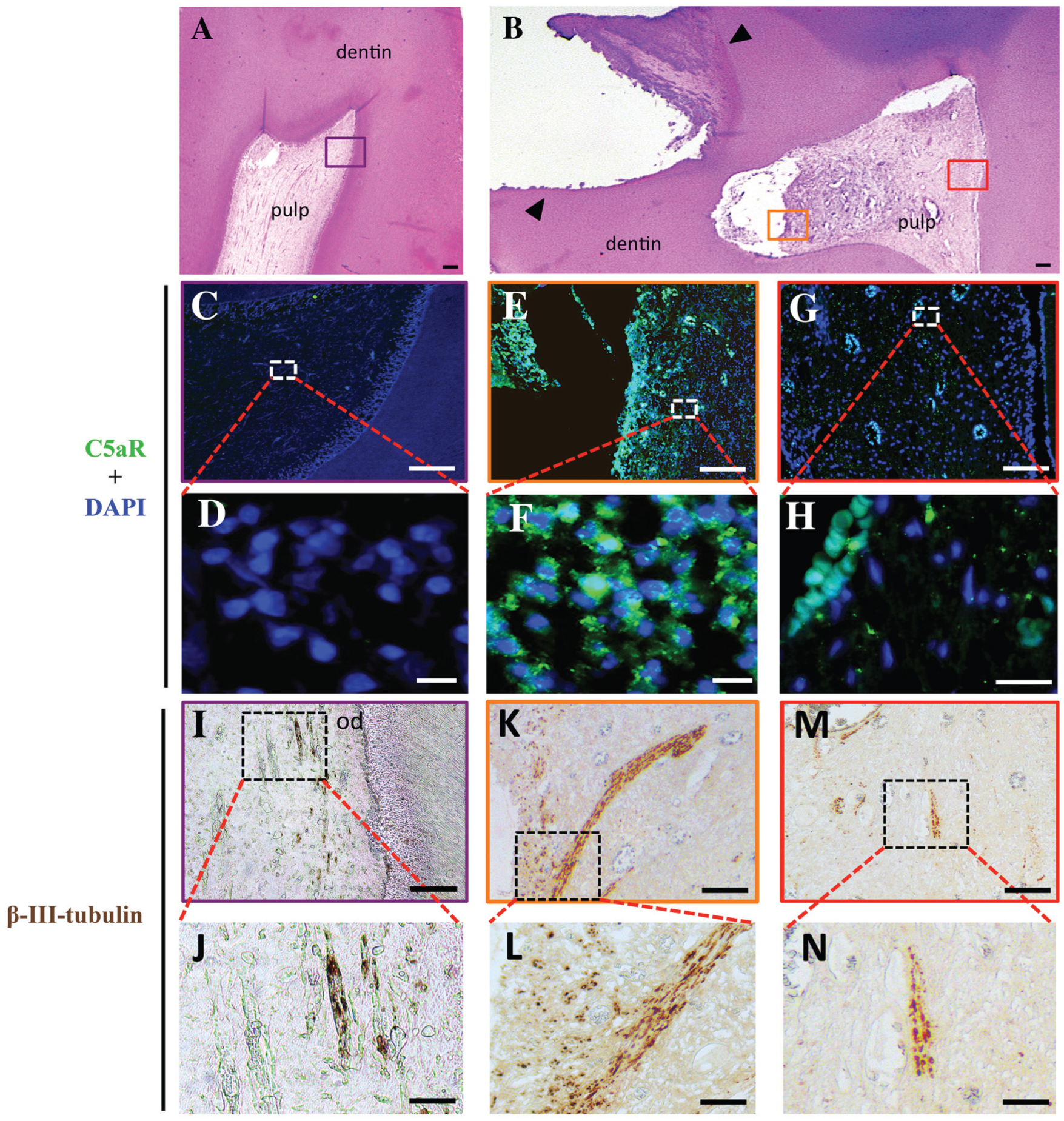
Expression of C5aR and β-III-tubulin in vivo. Immunohistochemistry was used to investigate the expression of C5aR and β-III-tubulin in intact (
C5aR Is Expressed in LTA-Stimulated Pulp Fibroblasts
Since Gram-positive bacteria constitute the predominant cariogenic bacteria (Martin et al. 2002), LTA was used to simulate the presence of these bacteria in the pulp. All pulp cells used in our experiments expressed the FSP marker (Fig. 3Aa–c); hereafter, these cells will be designated as pulp fibroblasts. Consistent with our in vivo results, we observed that few pulp fibroblasts expressed C5aR (Fig. 3Ad). After 6-h and 24-h LTA stimulation, all pulp fibroblasts expressed the C5aR (Fig. 3Ae, f). Merge images show that only LTA-stimulated fibroblasts expressed C5aR (Fig. 3Ag–i). Higher magnification of C5aR staining (Fig. 3Aj–l) indicated that 6 h after LTA stimulation, C5aR was mainly detected around the nucleus (Fig. 3Ak), whereas after 24 h, it was principally localized at cell membranes and in the cytoplasm (Fig. 3Al). Moreover, our in-cell Western assays (Fig. 3Ba, b) revealed that the C5aR staining in pulp fibroblasts was significantly higher 6 and 24 h after LTA stimulation than in untreated conditions. Taken together, these results indicate that there is a new expression of C5aR in LTA-stimulated fibroblasts.
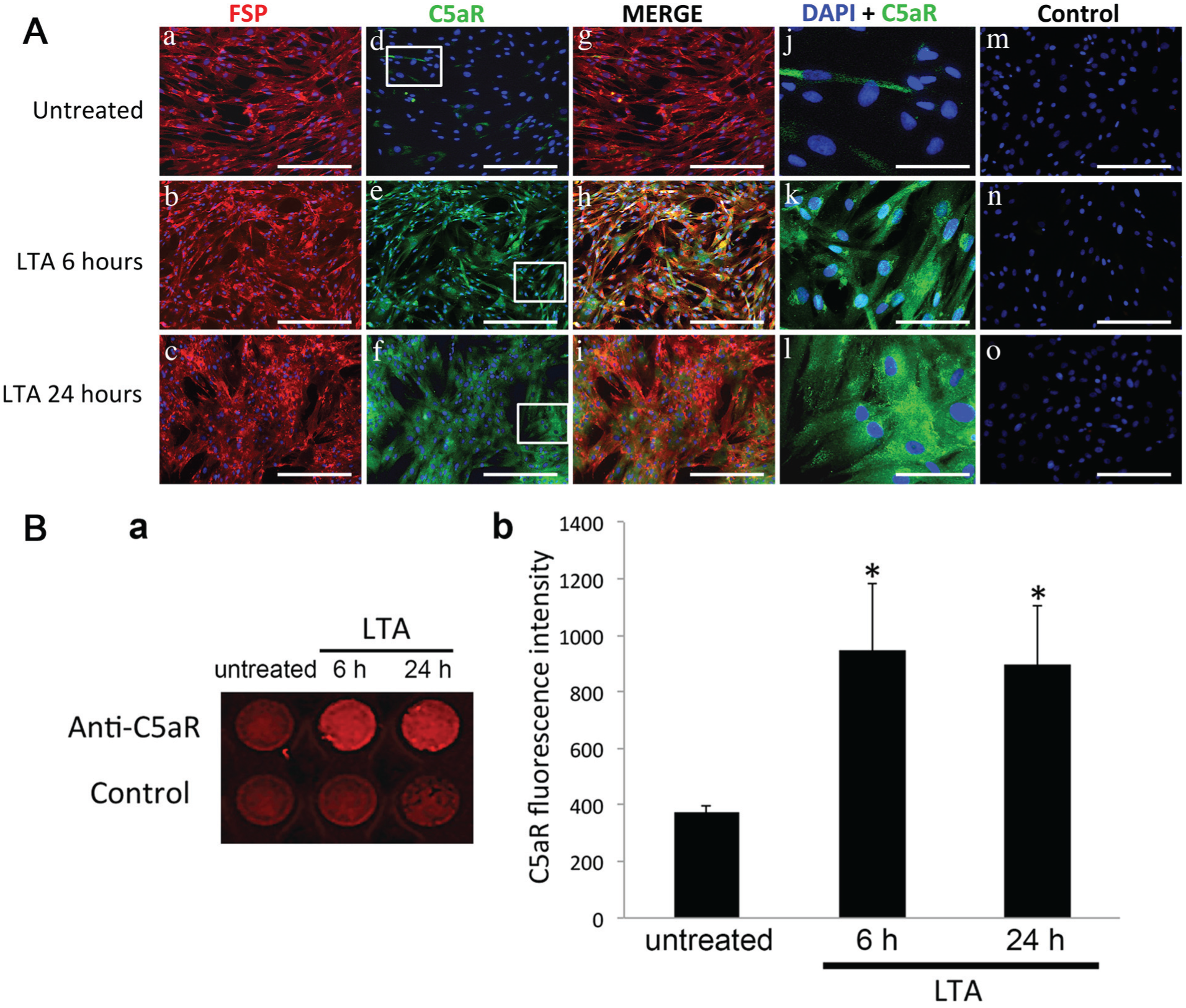
C5aR expression in human pulp cells. (
C5aR Is Activated in LTA-Stimulated Pulp Fibroblasts
We next decided to investigate the ability of C5a generated by LTA-stimulated pulp fibroblasts (Chmilewsky, Jeanneau, Laurent, et al. 2014) to activate their own C5aR. Unlike untreated fibroblasts (Fig. 4Aa–d), an intense phospho-C5aR and C5aR staining was detected in all pulp fibroblasts 24 h after LTA stimulation (Fig. 4Af–i). C5aR activation decreased when LTA-stimulated fibroblasts were incubated with a C5aR-specific antagonist (Fig. 4Ak–n). In-cell Western results (Fig. 4Ba, b) confirmed that C5aR was activated in LTA-stimulated cells, since the phospho-C5aR intensity was significantly higher than in untreated fibroblasts (Fig. 4Bb: black histograms vs. red line) at 6 and 24 h. The coincubation of LTA-stimulated fibroblasts with W54011 significantly reduced the phospho-C5aR intensity (Fig. 4Bb: blue vs. black histograms) at the same delays. Since all experiments have been realized in serum-free media (to avoid the presence of any complement molecules), the obtained results show that the C5a generated by pulp fibroblasts under LTA-stimulation activates their newly expressed C5aR.
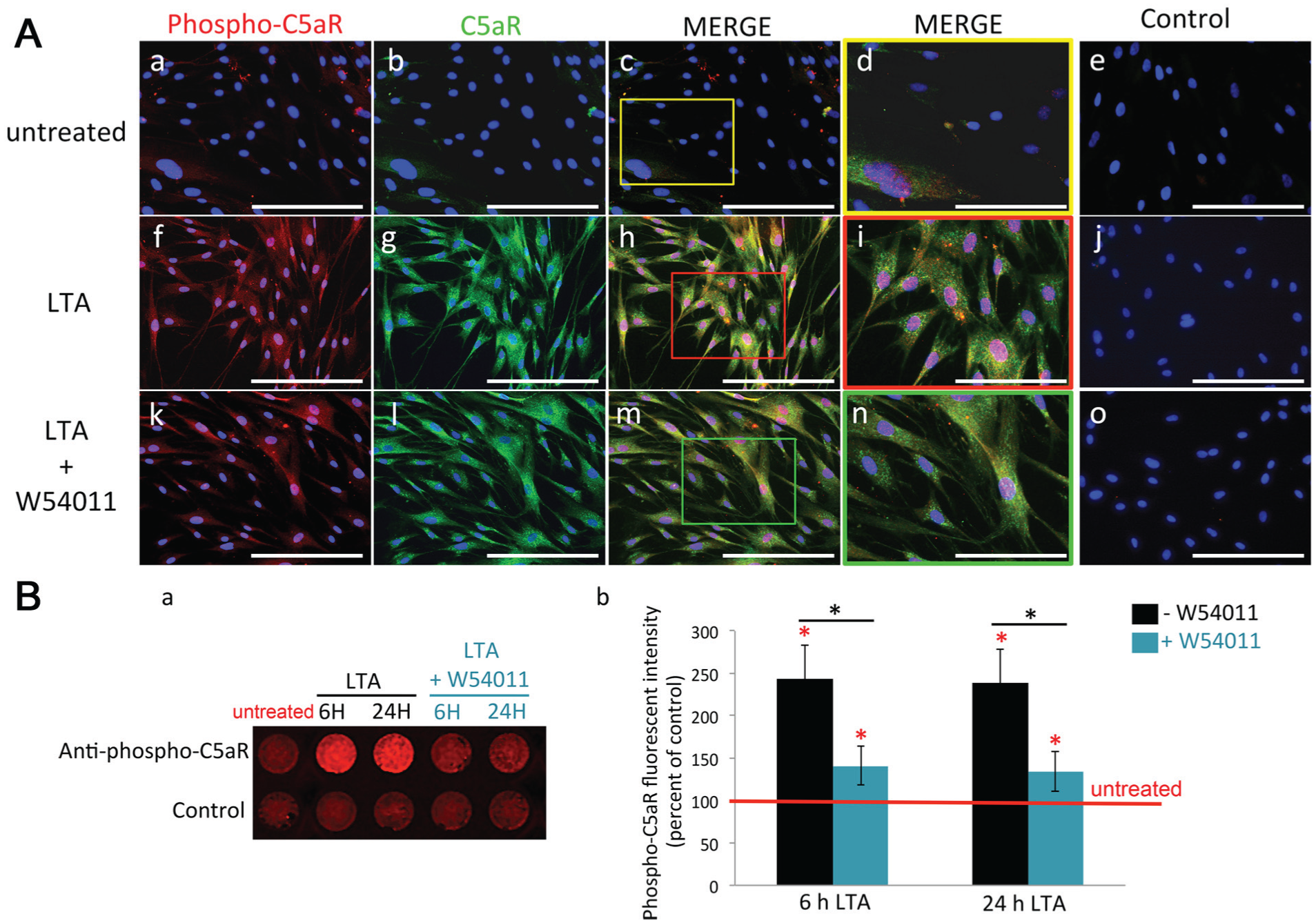
C5aR activation in human pulp fibroblasts. (
C5aR Is Required for the Modulation of BDNF Secretion in LTA-Stimulated Fibroblasts
We next investigated the potential implication of C5aR in BDNF secretion (Fig. 5A). LTA stimulation led to a significant increase of BDNF secretion at 6, 24, and 48 h (blue vs. pink histograms). Interestingly, the coincubation of LTA-stimulated fibroblasts with a C5aR-specific antagonist drastically decreased the secretion of this factor at 6, 24, and 48 h (green vs. blue histograms). These results show that C5aR activation is directly implicated in the modulation of BDNF secretion by LTA-stimulated fibroblasts. Surprisingly, incubation of LTA-stimulated fibroblasts with an additional source of C5a for 6, 24, or 48 h had no effect on BDNF secretion (black vs. blue histograms). These data suggest that the C5a, generated by LTA-stimulated fibroblasts (Chmilewsky, Jeanneau, Laurent, et al. 2014), is sufficient to activate the newly expressed C5aR efficiently. Moreover, in agreement with our previous results showing that only few fibroblasts express C5aR, the stimulation of these cells with recombinant C5a did not affect the basal BDNF secretion level (pink vs. gray histograms). These results show that the activation of the newly expressed C5aR is directly implicated in BDNF secretion by LTA-stimulated fibroblasts.
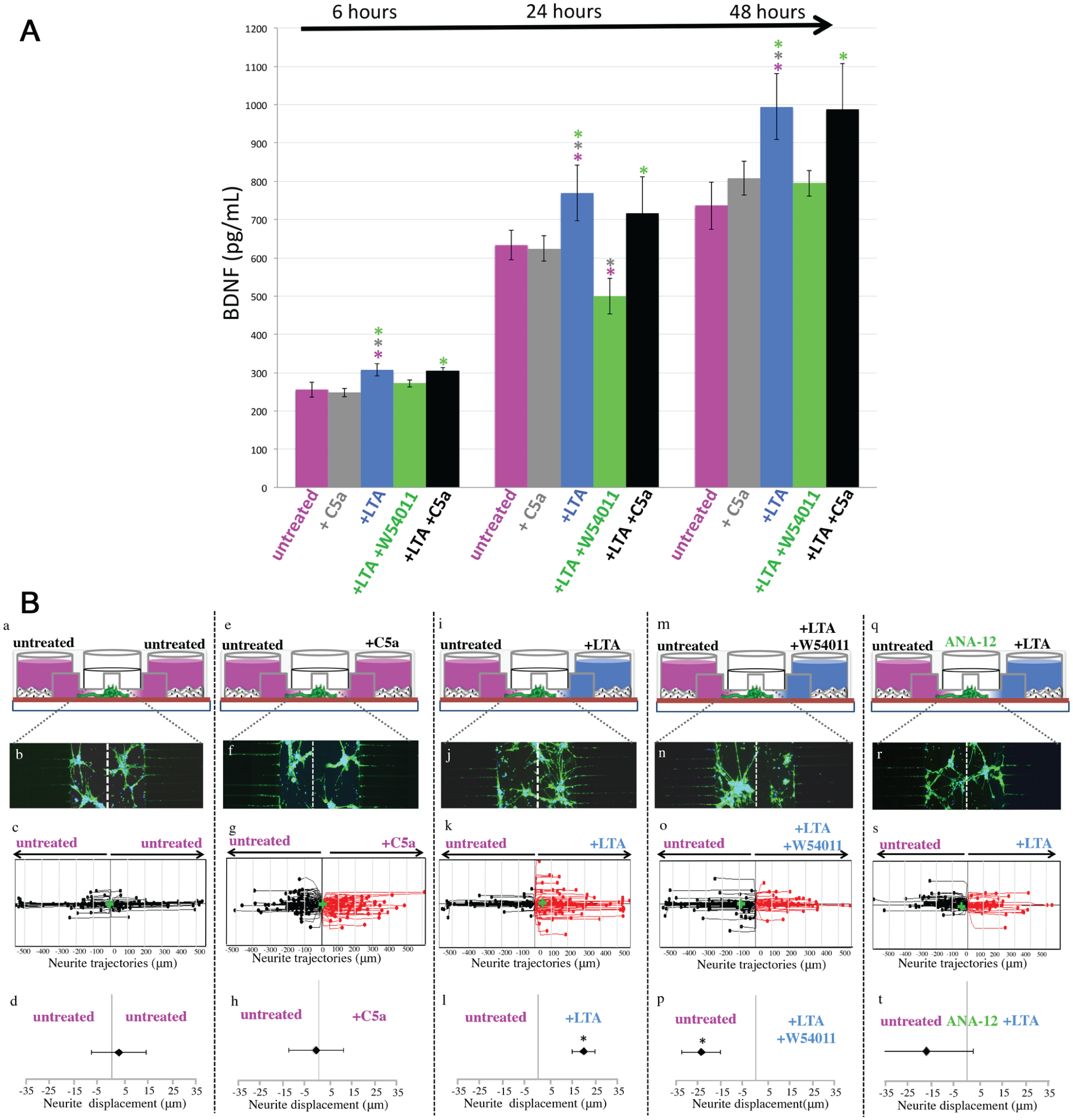
The interaction between C5a and lipoteichoic acid (LTA)–stimulated pulp fibroblasts controls the neurite outgrowth through the brain-derived neurotropic factor (BDNF) modulation. (
C5aR of LTA-Stimulated Fibroblasts Is Responsible for Neurite Outgrowth via Local BDNF Secretion
When pulp fibroblasts were cultured in the 2 peripheral chambers (Fig. 5Ba), the neurite outgrowth inside the microgrooves over a 48-h period was random and β-III-tubulin staining was the same on both sides of the central channel and the microgrooves (Fig. 5Bb, c). The statistical analysis demonstrated that global neurite outgrowth was in the center of the graph (Fig. 5Bd). In accordance with our previous results, which show that C5aR was not expressed by untreated fibroblasts, no effect was observed when the cells in one of the lateral chambers were stimulated only with C5a (Fig. 5Be–h). However, when pulp fibroblasts in one of the lateral chambers were stimulated with LTA (Fig. 5Bi), the β-III-tubulin staining was mainly detected in the stimulation side (Fig. 5Bj); the neurite trajectories (Fig. 5Bk) and the statistical analysis (Fig. 5Bl) showed that the global neurite outgrowth was significantly shifted toward the LTA stimulation side. The neurite outgrowth was drastically reduced with the C5aR-specific antagonist (Fig. 5Bm) and seemed to be redirected into the opposite side (Fig. 5Bn, o). This was confirmed by the shift of the global neurite outgrowth into the direction of untreated fibroblasts (Fig. 5Bp). These results demonstrated that the neurite recruitment in the direction of the LTA-injured site was dependent on the activation of pulp fibroblasts C5aR. Interestingly, the coincubation of neurons with an antagonist of tropomyosin receptor kinase B (TrkB) (Fig. 5Bq), a neuron’s receptor known to induce neurite outgrowth through its interaction with BDNF (Shih et al. 2013), reduced the neurite outgrowth to the LTA-stimulated side. The β-III-tubulin staining (Fig. 5Br) and representative neurite trajectories (Fig. 5Bs) appeared equally distributed toward untreated and LTA-stimulated cells. Indeed, the statistical analysis showed no significant displacement of the global neurite outgrowth (Fig. 5Bt), indicating that the neurite recruitment by LTA-stimulated fibroblasts was dependent on local BDNF secretion.
Discussion
In this study, we identified C5a/C5aR as a key initial signal in dental nerve sprouting in carious teeth. Our study reveals that pulp fibroblasts express C5aR in case of carious injury both in vivo and after LTA stimulation in vitro. We demonstrate that this newly expressed C5aR can be activated by C5a generated from complement activation by pulp fibroblasts. We provide direct evidence of C5aR critical implication in BDNF secretion modulation by pulp fibroblasts leading to a significant neurite outgrowth enhancement toward the injured site.
C5a is a major component of innate immunity, playing a critical role in the inflammatory process (Cruvinel et al. 2010). This fragment, which is quickly generated by the complement activation, is known to exert a chemotactic action on inflammatory cells through its interaction with C5aR (Klos et al. 2009; Ward 2009; Ricklin et al. 2010). Consistent with these data, we detected a high level of C5aR in the inflammatory areas of carious teeth. An observation of these teeth sections at higher magnifications revealed that all pulp cells localized beneath the injured sites and expressed C5aR, suggesting a potential C5aR expression by nonimmune cells in case of carious injury. Interestingly, many nonimmune cells from various tissues express C5aR either constitutively or in response to stimuli (Monk et al. 2007). Thus, our study adds the human pulp fibroblast as a new nonimmune cell able to express C5aR in response to external stimulations, since C5aR has mainly been detected all around the nucleus 6 h after LTA stimulation and in the cytoplasm and at the cell membrane after stimulation for 24 h. These data are supported by in-cell Western assays, which revealed a significant C5aR expression in pulp fibroblasts after LTA stimulation for 6 and 24 h. C5aR expression by various nonimmune cells suggested that the complement was implicated in other processes than inflammation. Indeed, several studies demonstrated that complement, particularly C5a, constitutes an important mediator of tissue regeneration (Ignatius et al. 2011; Mastellos et al. 2011; Lara-Astiaso et al. 2012). More recently, it has been demonstrated that C5a induces the recruitment of pulp progenitor cells (Chmilewsky et al. 2013; Chmilewsky, Jeanneau, Laurent, et al. 2014; Chmilewsky et al. 2015).
It is clearly established that dentin-pulp regeneration is a locally regulated process by the microenvironment modification (About 2013; Chmilewsky, Jeanneau, Dejou, et al. 2014). In case of carious injury, the acidic dissolution of dentin induces the liberation of extracellular matrix molecules and several growth factors directly implied in the dentin-pulp regeneration (Sloan and Smith 1999; Sloan et al. 2000). Moreover, the odontoblasts have also been identified as a source of regeneration signals in case of carious injury (Botero et al. 2006). However, deep carious injuries lead to the disintegration of odontoblasts beneath the injured sites (Fitzgerald et al. 1990). Interestingly, fibroblasts, which represent the major cell population within the pulp tissue, have previously been identified as a source of regeneration signals in case of traumatic injury (Tran-Hung et al. 2008). However, their ability to secrete regeneration signals in case of carious injury has not been given much attention. Our work highlights that BDNF secretion by cultured pulp fibroblasts was significantly enhanced after their incubation with LTA at 6, 24, and 48 h. Previous studies demonstrated that C5a directly affects the secretion of several growth factors in various nonimmune cells (Boor et al. 2007; Cheng et al. 2013). In accordance with these data, we demonstrated that C5aR acts as a positive regulator for BDNF as confirmed by W54011, which inhibited BDNF secretion by LTA-stimulated fibroblasts. Recently, human pulp fibroblast has been identified as the first nonimmune cell able to produce all complement molecules required for efficient complement activation after LTA stimulation, implying the production of C5a (Chmilewsky, Jeanneau, Laurent, et al. 2014). Here, we show, through phospho-C5aR staining in serum-free conditions, that the C5a generated by complement activation from molecules produced by LTA-stimulated fibroblasts activates its newly expressed C5aR. This is confirmed by W54011, which drastically decreases the C5aR-active form in LTA-stimulated fibroblasts. Moreover, BDNF quantification demonstrated that the C5a generated by LTA-stimulated fibroblasts (Chmilewsky, Jeanneau, Laurent, et al. 2014) is sufficient to control BDNF secretion level efficiently. Indeed, the use of an additional source of C5a to LTA-stimulated fibroblasts had no effect on BDNF secretion. In addition, the absence of a C5a effect on BDNF secretion by untreated cells confirmed that C5aR is involved in BDNF modulation only after LTA stimulation.
Neurotrophins are essential for the development and functional maintenance of the nervous system, participating in axonal growth and nerve sprouting (Bothwell 2014). It became evident that their upregulation after injury constitutes a critical step in tissue regeneration (Gordon 2009). The possibility to culture 2 different cell types inside the same AXIS devices and to submit neurons to 2 different stimulation gradients (Park et al. 2006) allowed us to evaluate the effects of C5aR activation by LTA-stimulated pulp fibroblasts on the neurite outgrowth. We demonstrated that neurite outgrowth occurs in the direction of LTA-stimulated fibroblasts. These data are consistent with our in vivo results showing a high level of nerve fibers on the injured pulp of deep carious teeth. So, the present study provides, for the first time, evidence of an efficient interaction between pulp fibroblasts and nerve sprouting during the dentin-pulp regeneration. Interestingly, our results revealed that C5aR controls the neurite outgrowth induced by LTA-stimulated fibroblasts, since C5aR functional inhibition reduces drastically the neurite outgrowth in the direction of the LTA stimulation. This is correlated with the nerve sprouting and C5aR colocalization observed beneath the injuries of human carious teeth. BDNF is known as a major neurite outgrowth inducer through its receptor TrkB (Shih et al. 2013; Yang et al. 2015). Our results suggest that the neurite outgrowth observed after LTA stimulation of pulp fibroblasts is guided by BDNF. Indeed, the coincubation of neurons with a TrkB-inhibitor prevented this neurite outgrowth. This could explain why, when LTA-stimulated fibroblasts were coincubated with W54011, the neurites outgrew in the direction of untreated cells. Indeed, BDNF quantification revealed that the inhibition of a C5a-C5aR interaction drastically reduced the BDNF secretion by LTA-stimulated pulp fibroblasts. Thus, the BDNF secreted by LTA-stimulated cells in the presence of W54011 detected at 24 h was inferior to that secreted by untreated fibroblasts. These results also suggest that C5aR activation might be implicated in the regulation of other molecules implied in neurite outgrowth. Although poorly understood, previous studies suggested that nerve growth factor (NGF) upregulation could be responsible for dental nerve sprouting (Byers et al. 1992). Here, we bring additional data in the comprehension of this event by demonstrating that LTA-stimulated fibroblasts mediated the enhancement of neurite outgrowth via the BDNF upregulation and its orchestration by the C5aR. This work is in concordance with previous studies, demonstrating in vitro that pulp cells secrete neurotrophins, which are known to induce nerve growth from the trigeminal ganglion into the pulp in vivo (Lillesaar et al. 1999).
Our findings highlight a new mechanism in dentin-pulp regeneration, linking, for the first time, pulp fibroblast to the nerve sprouting through the complement activation. In this mechanism, the BDNF modulation by human pulp fibroblasts localized beneath the injured site, through the C5a-C5aR interaction, could locally induce a neurite outgrowth/increased nerve sprouting, which is required for efficient dentin-pulp regeneration (Byers and Taylor 1993). Thus, these results may provide a useful future therapeutic tool in targeting the fibroblasts in dentin-pulp regeneration.
Author Contributions
F. Chmilewsky, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; I. About, contributed to design, data analysis, and interpretation, drafted and critically revised the manuscript; S.-H. Chung, contributed to data interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
Acknowledgements
We thank Dr. Jean-Charles Gardon for providing the third molars and Dr. David A. Reed from the department of Oral Biology, University of Illinois at Chicago, for kindly providing the microscope (Leica DM2000) used for generated histologic pictures.
This study was supported by start-up funds from University of Illinois at Chicago College of Dentistry (to S.-H. Chung).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.