
Abstract
To elucidate if microglial P2Y12 receptor (P2Y12R) mechanisms are involved in the trigeminal spinal subnucleus caudalis (Vc; also known as the medullary dorsal horn) in intraoral cancer pain, we developed a rat model of tongue cancer pain. Squamous cell carcinoma (SCC) cells were inoculated into the tongue of rats; sham control rats received the vehicle instead. Nociceptive behavior was measured as the head-withdrawal reflex threshold (HWRT) to mechanical or heat stimulation applied to the tongue under light anesthesia. On day 14 after the SCC inoculation, activated microglia and P2Y12R expression were examined immunohistochemically in the Vc. The HWRT was also studied in SCC-inoculated rats with successive intra–cisterna magna (i.c.m.) administration of specific P2Y12R antagonist (MRS2395) or intraperitoneal administration of minocycline, a microglial activation inhibitor. Tongue cancer was histologically verified in SCC-inoculated rats, within which the HWRT to mechanical stimulation of the tongue was significantly decreased, as compared with that of vehicle-inoculated rats, although the HWRT to heat stimulation was not. Microglia was strongly activated on day 14, and the administration of MRS2395 or minocycline reversed associated nocifensive behavior and microglial activation in SCC-inoculated rats for 14 d. The activity of Vc wide dynamic range nociceptive neurons was also recorded electrophysiologically in SCC-inoculated and sham rats. Background activity and noxious mechanically evoked responses of wide dynamic range neurons were significantly increased in SCC-inoculated rats versus sham rats, and background activity and mechanically evoked responses were significantly suppressed following i.c.m. administration of MRS2395 in SCC-inoculated rats as compared with sham. The present findings suggest that SCC inoculation that produces tongue cancer results in strong activation of microglia via P2Y12 signaling in the Vc, in association with increased excitability of Vc nociceptive neurons, reflecting central sensitization and resulting in tongue mechanical allodynia.
Introduction
Neuropathic and inflammatory mechanisms are involved in oral cancer pain (Mantyh 2006). Moreover, orofacial cancer pain is partially mediated by cancer-induced peripheral inflammation (Harano et al. 2010). Microglia in the trigeminal spinal subnucleus caudalis (Vc) is also activated in models of orofacial inflammation, trigeminal nerve injury, or facial cancer (Okada-Ogawa et al. 2009; Sago et al. 2012; Old et al. 2015). Activated microglia releases brain-derived neurotrophic factor binding to tropomyosin receptor kinase B in spinal dorsal horn neurons, thus contributing to neuronal hyperexcitability (Coull et al. 2005; Ren and Dubner 2007). Collectively, these findings suggest that microglia in the Vc may be activated in association with cancer growth, which contributes to the central sensitization of Vc neurons.
The P2Y12 receptor (P2Y12R) is activated by ATP binding in the spinal dorsal horn (Inoue and Tsuda 2012). It was recently reported that P2Y12R expression in microglia is involved in activation of these cells in the spinal dorsal horn following sciatic nerve injury (Kobayashi et al. 2008; Kobayashi et al. 2013). It is thus very likely that P2Y12R is the key receptor involved in microglial activation under pathologic conditions. However, it is not fully understood how P2Y12 signalingcontributes to cancer pain mechanisms. Therefore, the aimof the present study was to evaluate if microglial P2Y12R mechanisms are involved in intraoral cancer pain.
Materials and Methods
Animals
A total of 178 male Fischer rats (100 to 200 g; Japan SLC) were used in the present study. Rats were housed in a temperature-controlled room (23 °C) with a 12:12-h light:dark cycle, and water and food were offered ad libitum. This study was approved by the Animal Experimentation Committee at Nihon University (AP15D032), and the experiments were conducted according to the guidelines of the International Association for the Study of Pain (Zimmermann 1983). All efforts were made to minimize animal suffering and reduce the number of animals utilized.
Tongue Cancer Model
Rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal; Schering Plough) and placed on the warm mat. The mouth was gently opened and the tongue pulled out. A 30-µL suspension of rat squamous cell carcinoma (SCC) cells (SCC-158, JCRB0231; JCRB) was inoculated into the right edge of the tongue by a 26-gauge needle. The suspension consisted of SCC cells (5 × 106) with 30 µL of 0.01M phosphate-buffered saline (PBS).
Histologic Examination of the Tongue
On days 3, 6, 14, and 21 after the SCC or PBS inoculation (n = 8 in each), rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal) and perfused transcardially with isotonic saline. Their tongues were removed and embedded in Tissue Tek (Sakura Finetechnical). The tongue sections were cut with cryostat, the temperature of which was kept at −20 °C, and then placed on slides. Sections were moved to room temperature and then fixed with 4% paraformaldehyde for 1 h. The sections were stained with hematoxylin and eosin to assess any pathologic changes indicative of tumor formation.
Head Withdrawal Reflex Threshold Measurement
In advance, rats were sufficiently anesthetized with 2% isoflurane (Mylan), and the supply of isoflurane was subsequently cut off. An adequate level of anesthesia was defined once noxious pressure applied to the hind paw induced an identical weak flexion reflex of the hind limb (Urata et al. 2015). Then, rats were applied graded heat stimulation (PBS-inoculated: n = 5, SCC-inoculated: n = 5) or mechanical stimulation (PBS- inoculated: n = 5, SCC-inoculated: n = 5) to measure head-withdrawal reflex thresholds under the defined depth of anesthesia within 1 min before the animals started to regain consciousness. The threshold intensities were measured for evoking head-withdrawal reflex to mechanical stimulation (0 to 150 g, 10 g/s, cutoff: 150 g; Panlab) and heat stimulation (35°C to 55 °C, 1 °C/s, cutoff: 55 °C; Intercross) of the left edge of tongue (6-mm2 area, 3 mm posterior from tip of tongue). The stimulus intensity was gradually increased to elicit head withdrawal accompanying a clear tongue withdrawal response; the heat or mechanical head-withdrawal reflex threshold (HHRT or MHRT) was defined as the lowest temperature or pressure required for eliciting head withdrawal, respectively. Every time the graded heat or mechanical stimulation was performed once, rats were sufficiently anesthetized with 2% isoflurane to reestablish the defined level of anesthesia. The mechanical or heat stimulation was applied 3 times at 5-min intervals, and the mean values of the MHRT and HHRT were determined. The value at baseline (i.e., before any drug administration; see below) was defined as the 100% value.
Immunohistochemistry
Rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal) and perfused with cold 4% paraformaldehyde in 0.1M phosphate buffer; then, brainstem was removed and immersed in the same fixative for 2 d on days 3, 4, and 14 (n = 5 in each) following SCC or PBS inoculation. By the same token, brainstem was removed and immersed on day 14 in vehicle-administrated SCC-inoculated rats (n = 5), MRS2395-administrated SCC-inoculated rats (n = 5), and vehicle-administrated PBS-inoculated rats (n = 5). The brainstems (±1 mm from obex) were immersed in 20% sucrose for 12 h; then, 30-µm frontal sections were cut on a freezing microtome, and every fifth section was corrected in 0.01M PBS. Activated microglia showed dramatic morphologic changes (from ramified to ameboid) and intense increases in the expression of microglial marker Iba1. Therefore, P2Y12R and/or Iba1 immunohistochemistries were conducted to detect activated microglia and P2Y12R expression.
Free-floating tissue sections were incubated in rabbit anti-P2Y12R polyclonal antibody corresponding to amino acid residues 125 to 142 of human P2Y12 with an additional N-terminal cysteine as the immunogen (1:100, P4871; Sigma-Aldrich; Shirasaki et al. 2013) and/or goat anti-Iba1 polyclonal antiserum corresponding to C-terminus of Iba1 (1:500, 019-19741; Wako Chemicals; Bosco et al. 2011) in 0.3% TritonX-100 in 0.01M PBS containing 4% normal donkey serum overnight at 4 °C. Sections were also incubated in mouse anti-NeuN monoclonal antibody corresponding to purified cell nuclei from mouse brain (1:1,000, MAB377; Millipore; Honda et al. 2008) and rabbit anti-P2Y12R polyclonal antibody (1:100) in 0.3% TritonX-100 in 0.01M PBS containing 4% normal donkey serum overnight at 4 °C to define if P2Y12R is expressed in neurons. After rinsing with 0.01M PBS, sections were incubated in Alexa Fluor 488 donkey anti-rabbit IgG (1:200 in 0.01M PBS, R37118; Invitrogen) and Alexa Fluor 568 donkey anti-goat IgG (1:200 in 0.01M PBS, A11057; Invitrogen) or Alexa Fluor 594 anti-mouse IgG (1:200 in 0.01M PBS, A21203; Invitrogen) for 2 h at room temperature.
The sections that were reacted by no primary antibody were also incubated in Alexa Fluor 488 donkey anti-rabbit IgG (1:200), Alexa Fluor 568 donkey anti-goat IgG (1:200), or Alexa Fluor 594 anti-mouse IgG (1:200). After being rinsed with 0.01M PBS, sections were coverslipped in mounting medium (Thermo Fisher Scientific). Iba1-, P2Y12R-, and NeuN-immunoreactive (Iba1-IR, P2Y12R-IR, and NeuN-IR) cells were observed under fluorescent microscope (Keyense BZ9000; Keyense). With a computer-assisted imaging analysis system (ImageJ 1.37; National Institutes of Health; Kiyomoto et al. 2013), the density of Iba1-IR cells was measured in the superficial layer area (313.37 × 3133.69 µm2) of the dorsal portion in Vc, which has been shown to receive sensory inputs from the tongue. The density of Iba1-IR cells in Vc was measured from 1 section at the obex level, and the mean density of Iba1-IR cells was calculated in each group. All immunohistochemical measurements were conducted under a blind condition, performed by observers who did not know which treatment the animals received.
MRS2395 and Minocycline Administration
Rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal) and placed in a stereotaxic frame. A small hole was made in the occipital bone after a small incision was made in the neck skin, and a polyethylene tube (0.8 mm in diameter; Natsume) was inserted into the cisterna magna for intra–cisterna magna (i.c.m.) administration of drugs. The specific P2Y12 R antagonist, 2,2-dimethyl-propionic acid 3-(2-chloro-6-methylaminopurin-9-yl)-2-(2,2-dimethyl-propionyloxymethyl)-propylester (4.5 mM, MRS2395; Sigma-Aldrich), was dissolved with 25% dimethyl sulfoxide in 0.01M PBS. The MRS2395 (n = 6) or vehicle (n = 6) was continually administrated intracisternally for 14 d after SCC inoculation via an osmotic mini pump (0.5 µL/h, Alzet model 2002; Durect Corporation). Vehicle (n = 6) was also continually administrated for 14 d after PBS inoculation. Once-daily intraperitoneal administration of minocycline (30 mg/kg/d; Sigma-Aldrich) dissolved with 0.01M PBS (n = 5) or vehicle (n = 5) was performed for 14 successive days (days 0 to 13) after SCC inoculation. The vehicles for MRS2395 and minocycline were 20% dimethyl sulfoxide and 0.01M PBS, respectively. The MHRT was measured daily as described above.
Single-Neuron Recording
On day 14, the rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal) and rigidly secured in the stereotaxic frame. The medulla and upper cervical spinal cord were exposed, and dura was removed. During the recording, rats were immobilized by pancuronium bromide (0.6 mg/kg/h, intravenous; Schering-Plough). End-tidal CO2 concentration was kept at 3.5% to 4.5%. Body temperature was monitored and maintained at 37 °C. After tracheotomy, rats were respired artificially with oxygen (2 L/min) and isoflurane (1.0% to 2.0%). During the single-unit recording, heart rate was continually monitored. The concentration of isoflurane was appropriately increased once the heart rate was increased after the application of mechanical stimulation. Enamel-coated tungsten electrodes (FHC) were advanced into the Vc, and single neuronal activities were recorded. We used gentle brush stimulus to search the receptive field of the tongue mucosa. When the neuron responded to gentle mechanical stimulus, graded mechanical stimuli were applied to the tongue. Thus, all neurons were classified as wide dynamic range (WDR) neurons on the basis of their responses as previously described (Iwata et al. 1999). We analyzed WDR neurons receiving input from the tongue mucosa. The responses of WDR neurons to brush and to nonnoxious (10 and 30 g) and noxious (60, 90, 120, and 150 g) pinch—which was applied for 5 s at 1-min intervals—were analyzed. WDR neurons were studied in PBS-inoculated rats (n = 6) and SCC-inoculated rats (n = 8) as well as in SCC-inoculated rats receiving MRS2395 (SCC/MRS2395: n = 5) and SCC-inoculated rats receiving vehicle (SCC/vehicle: n = 5). Spikes were sorted, and spike frequencies were analyzed with the Spike II software (CED). Peristimulus time histograms (bin width = 1 s) were generated in response to each stimulus. Background (BG) activity was recorded for 1 min before application of the brush stimulus. We defined neuronal responses as occurring when the mean firing frequencies were >2 standard deviations of the mean BG activity. After neuronal recording, the recording sites in Vc were identified histologically.
Statistical Analysis
Data are expressed as mean ± SEM. Statistical analyses were performed by Student’s t test or 1- or 2-way repeated-measures analysis of variance, followed by Bonferroni’s multiple-comparison tests, where appropriate. A value of P < 0.05 was considered significant.
Results
Macroscopic Examination and Inflammatory Response of the Tongue
The thickness of SCC-inoculated tongue significantly increased on day 6 postinoculation (SCC-inoculated: 3.7 ± 0.1 mm, PBS- inoculated: 3.1 ± 0.1 mm; Appendix Fig. 1A). Moreover, the maximum sectional area of tumoral mass in SCC-inoculated tongue grew larger in a time-dependent manner (Appendix Fig. 1B). After the SCC or PBS inoculation, inflammatory cells such as monocytes, macrophages, and lymphocytes infiltrated at the tumor margins in SCC-inoculated rats and at the PBS-inoculated sites in PBS-inoculated rats (Appendix Fig. 1C). On day 3, there was no significant difference in the number of inflammatory cells between SCC- and PBS-inoculated rats (Appendix Fig. 1D). On days 6, 14, and 21, the number of inflammatory cells in SCC-inoculated rats was larger than that of PBS-inoculated rats. Moreover, the number of inflammatory cells on days 6, 14, and 21 in PBS-inoculated rats was smaller than that of day 3.
Histologic Examination of the Tongue in SCC-Inoculated Rats
Histologic examination of SCC- or PBS-inoculated tongues revealed tumor formation on days 3, 6, 14, and 21 after SCC inoculation; the large expansion due to SCC cell proliferation was seen in the tongue; and the tumor margin had reached the contralateral side of the tongue by day 21 (Fig. 1A). After PBS inoculation, no histologic changes were observed.
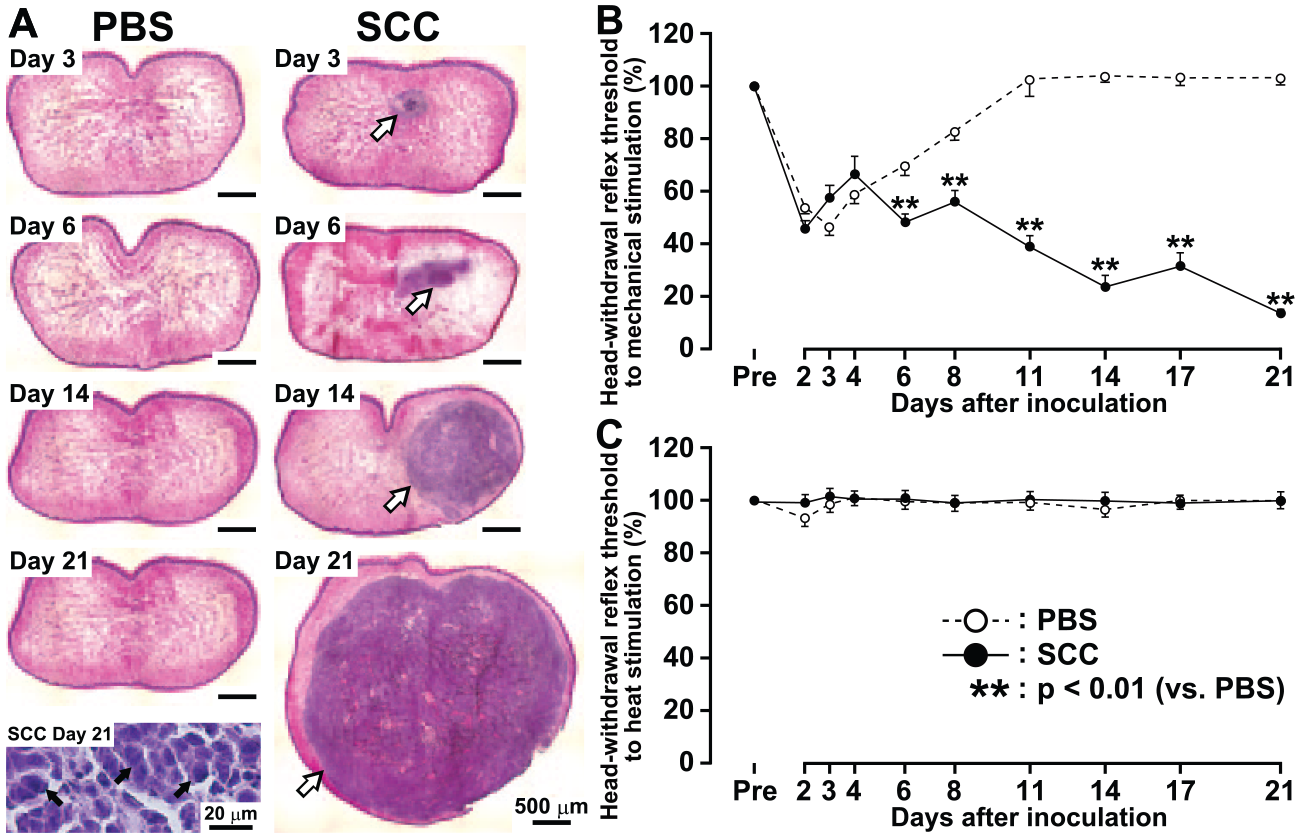
Tongue histologic observations and pain sensitivity following phosphate-buffered saline (PBS) or squamous cell carcinoma (SCC) inoculation. Histologic examination of the tongue obtained from PBS- and SCC-inoculated rats (n = 8 in each;
Mechanical and Heat Sensitivities of the Tongue Following SCC Inoculation
The MHRT of the tongue was significantly reduced on days 3 and 6 to 21 after SCC inoculation when compared with that of PBS-inoculated rats (Fig. 1B). However, the HHRT of the tongue was not changed after SCC inoculation during the experimental period (Fig. 1C).
Microglial Activation
Many Iba1-IR cells were observed in the dorsal portion of Vc on day 14 after SCC inoculation (Fig. 2A). On day 3, Iba1-IR cells in Vc were apparent in PBS- and SCC-inoculated rats (Fig. 2B, C). On day 4, the density of Iba1-IR cells was reduced in PBS- and SCC-inoculated rats (Fig. 2D, E). No difference of the density of Iba1-IR cells was observed between PBS- and SCC-inoculated rats on days 3 and 4. On day 14, the mean density of Iba1-IR cells with large soma and short processes was significantly larger in SCC-inoculated rats versus PBS-inoculated rats (Fig. 2F–H).
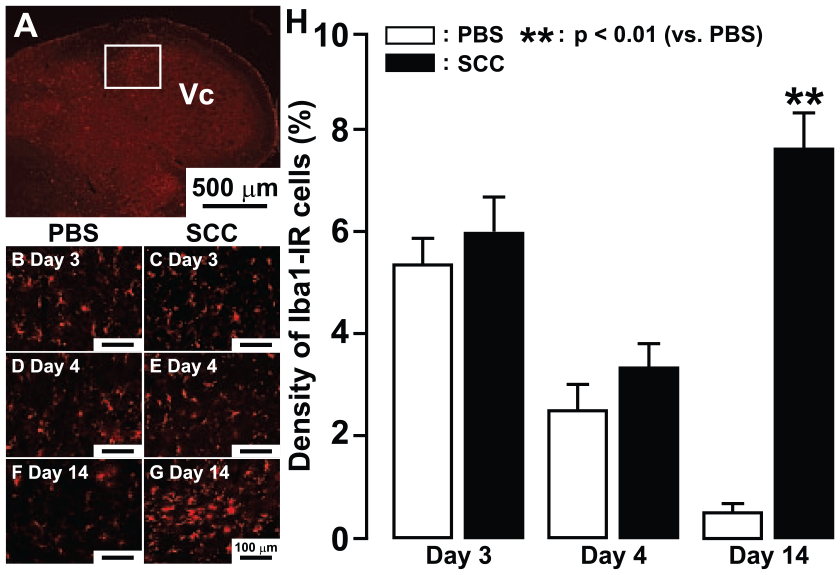
Microglial activation in Vc following phosphate-buffered saline (PBS) or squamous cell carcinoma (SCC) inoculation. Low-magnification photomicrograph of Iba1-immunoreactive (Iba1-IR) cells in the trigeminal spinal subnucleus caudalis (Vc) in PBS-inoculated rats on day 14. The mean density of Iba1-IR cells in the rectangle area in the Vc was calculated each day (
Effect of Minocycline or MRS2395 Administration on Microglial Activation and Mechanical Sensitivity
The mean density of Iba1-IR cells was significantly reduced in SCC-inoculated rats on day 14 following intraperitoneal administration of minocycline for 14 d (Fig. 3A–C). Furthermore, at 2 to 14 d, the MHRT was also significantly higher in SCC-inoculated rats receiving minocycline than in SCC-inoculated rats receiving vehicle at 2 to 14 d (Fig. 3D).
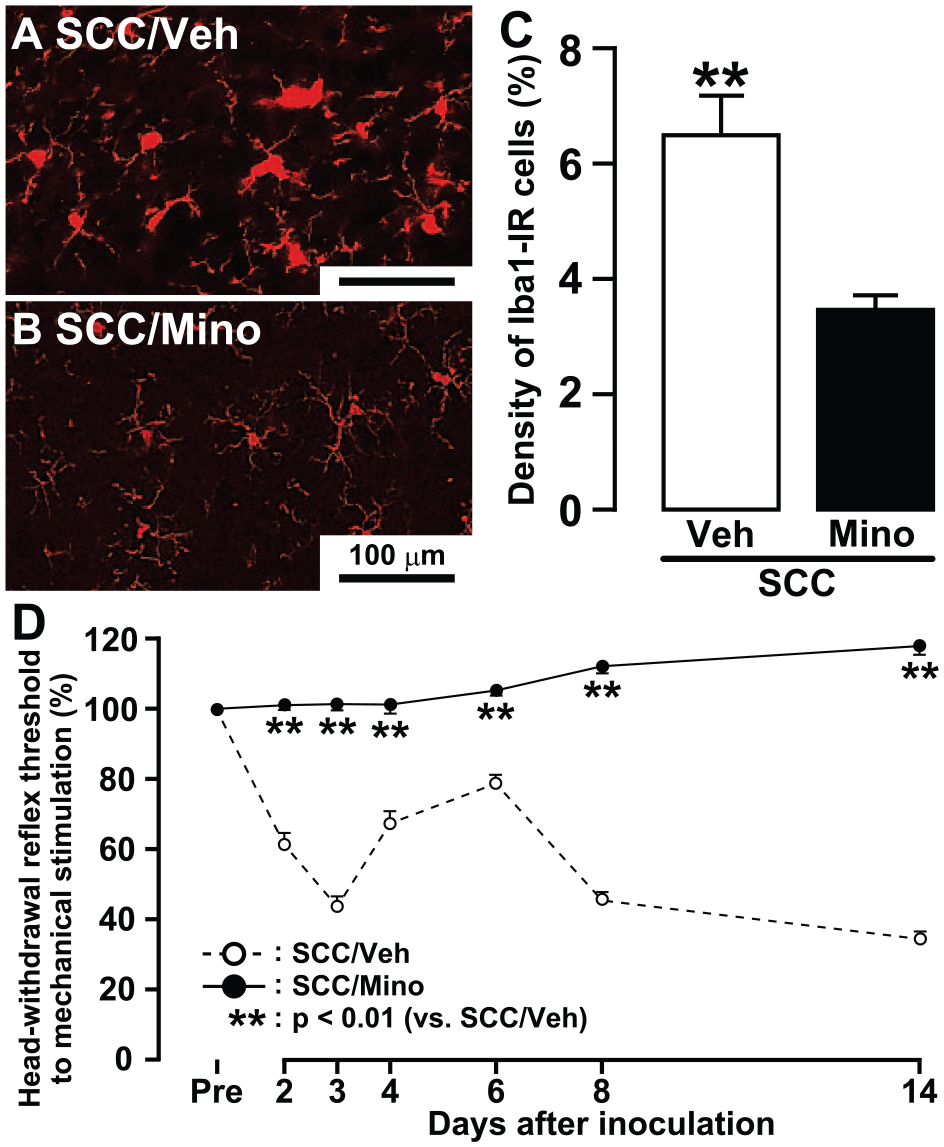
Changes in microglial activation in trigeminal spinal subnucleus caudalis and tongue mechanical sensitivity following minocycline administration in squamous cell carcinoma (SCC)–inoculated rats. Photomicrographs of Iba1-immunoreactive (Iba1-IR) cells in intraperitoneal vehicle-administrated (Veh;
NeuN-IR cells or Iba1-IR cells showed P2Y12R-IR inSCC-inoculated rats on day 14 (Fig. 4A, B). With Alexa Fluor 488 anti-rabbit IgG, Alexa Fluor 568 anti-goat IgG, or Alexa Fluor 594 anti-mouse IgG as secondary antibody, no nonspecific labeling was observed in the absence of primary antibody (Fig. 4C). Thus, we tested the effect of i.c.m. administration of specific P2Y12R antagonist MRS2395 on the expression of Iba1-IR cells and the MHRT in SCC-inoculated rats. The density of Iba1-IR cells was significantly smaller in SCC-inoculated rats receiving MRS2395 or in PBS-inoculated rats receiving vehicle as compared with SCC-inoculated rats receiving vehicle (Fig. 4D). The MHRT of the tongue was not changed during continuous i.c.m. administration of MRS2395 in SCC-inoculated rats, whereas on day 14, the MHRT was significantly lower and higher in SCC-inoculated rats administrated MRS2395 relative to PBS-inoculated rats administrated vehicle and SCC-inoculated rats administrated vehicle, respectively (Fig. 4E).
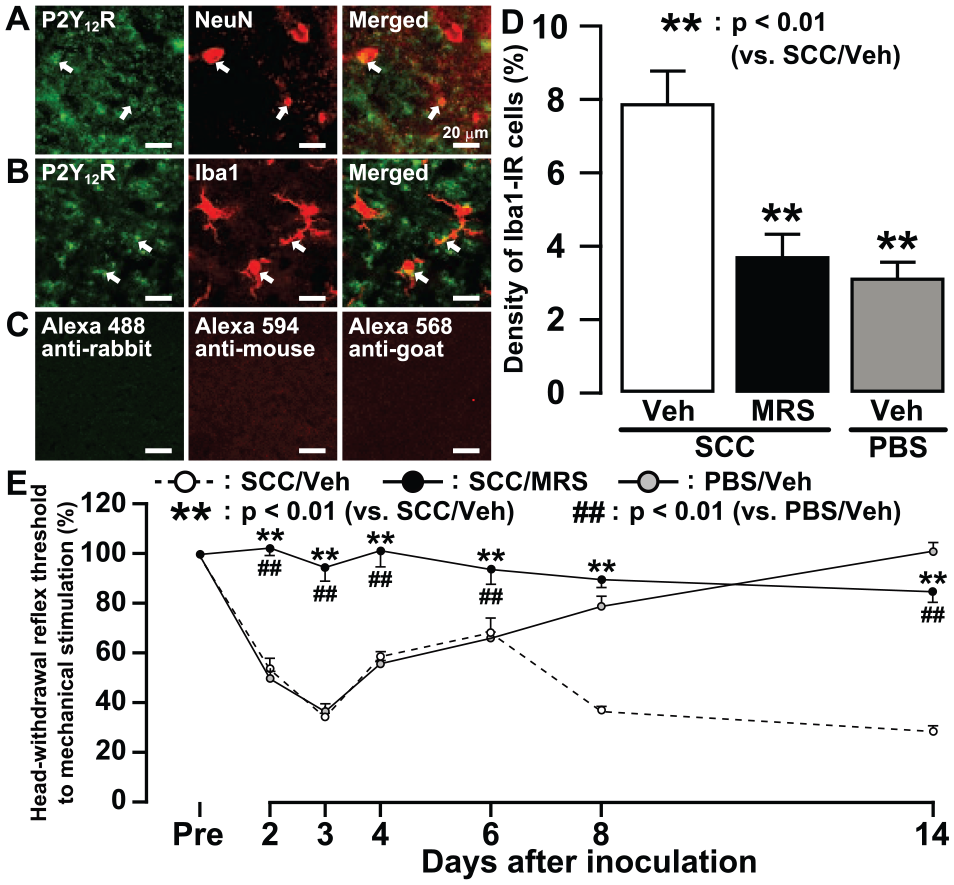
Changes in microglial activation in trigeminal spinal subnucleus caudalis (Vc) and tongue mechanical sensitivity following MRS2395 (MRS) administration in squamous cell carcinoma(SCC)–inoculated rats. Photomicrographs of P2Y12R-IR and NeuN-IR cells (
Effect of MRS2395 Administration on Vc Nociceptive Neuronal Activity in SCC-Inoculated Rats
A total of 45 WDR neurons (PBS: 12 neurons, SCC: 16 neurons, SCC/vehicle: 8 neurons, SCC/MRS2395: 9 neurons) were recorded as responding to nonnoxious and noxious mechanical stimulation of the tongue, though not heat stimulation (Fig. 5A–D). There was evidence of central sensitization in this intraoral cancer pain model, since the mean BG activity was significantly higher in SCC-inoculated rats versus PBS-inoculated rats (Fig. 5E). The mean response to 90 to 150 g of mechanical stimuli was also significantly larger in SCC-inoculated rats than in PBS-inoculated rats (Fig. 5F). Furthermore, the mean BG activity and mean responses to 60 to 150 g of mechanical stimuli were significantly smaller in SCC-inoculated rats receiving MRS2395 relative to SCC-inoculated rats receiving vehicle (Fig. 5G, H).
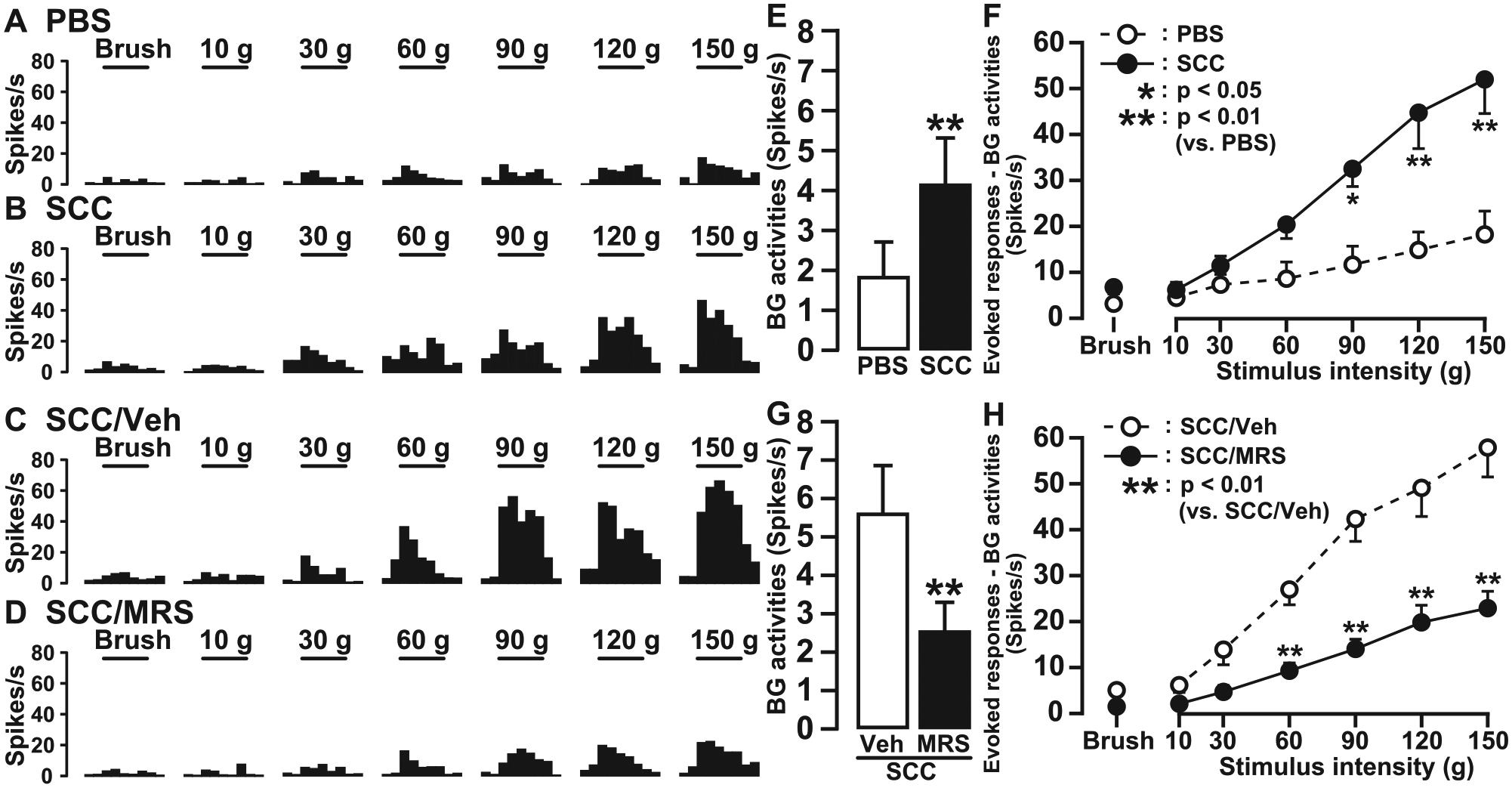
Single-neuron activities of trigeminal spinal subnucleus caudalis (Vc) wide dynamic range (WDR) neurons on day 14. Typical peristimulus time histograms of WDR neurons in phosphate-buffered saline (PBS)–inoculated rats (n = 6;
Discussion
This is the first documentation that trigeminal central sensitization of Vc nociceptive neurons accompanied by P2Y12R expression in several activated Vc microglia occurs in an intraoral cancer pain model. Further novel findings were that mechanical hypersensitivity of the tongue was apparent in SCC-inoculated rats. In addition, microglia was activated in the Vc 3 d after SCC or PBS inoculation and 14 d after SCC inoculation, and administration of MRS2395 or minocycline reversed the nocifensive behavior and microglial activation in SCC-inoculated rats. The BG activity and pinch-evoked responses of Vc nociceptive neurons receiving afferent inputs from the tongue were also significantly enhanced in the SCC-inoculated rats; such neuronal changes are indicative of central sensitization (Chiang et al. 2011). The enhancement was significantly reduced by administration of MRS2395 or minocycline. These findings suggest that tongue cancer is associated with microglial activation via P2Y12 signaling in the Vc, leading to central sensitization of Vc nociceptive neurons and tongue mechanical allodynia.
On day 14, the MHRT of the tongue was significantly lower in SCC-inoculated rats than in PBS-inoculated rats; yet, there were no changes in HHRT of the tongue. We also observed a significant decrease of MHRT in SCC- and PBS-inoculated rats on day 3. This decrement of MHRT on day 3 may be caused by the injected solutions themselves. These data are consistent with clinical observations of humans that many oral cancer pain patients complained of mechanical allodynia caused by orofacial movements (Cuffari et al. 2006; Lam and Schmidt 2011). This difference between mechanical and thermal responses in SCC-inoculated rats could be related to differences in neuropeptide involvement. It has been reported that neuropeptides such as calcitonin gene-related peptide and substance P are significantly expressed in trigeminal ganglion or dorsal root ganglion neurons following orofacial inflammation, whereas they are downregulated following nerve injury (Tsuboi et al. 2015). However, further studies are necessary to evaluate exact mechanisms underlying response differences between mechanical and heat stimuli in SCC-inoculated rats.
It has been well documented that, following trigeminal nerve injury or orofacial inflammation, Vc nociceptive neurons are strongly activated and sensitized, with increased BG activity and mechanical- and/or thermal-evoked responses (Chiang et al. 2011; Sessle 2011). A barrage of action potentials is generated in primary afferent neurons associated with peripheral nerve injury or tissue inflammation (Smith et al. 2013; Honda et al. 2014). Following hyperexcitability of primary afferent neurons, nociceptive neurons in the central nervous system become hyperactive, and microglia in Vc are strongly activated via the p38 mitogen-activated protein kinase cascade (Ikeda et al. 2012; Cao et al. 2015). In association with microglial activation, various receptors are overexpressed in microglia, such as purinergic receptors, chemokine receptors, and toll-like receptors (Ji et al. 2009; Gao and Ji 2010; Ren and Dubner 2010). We observed strong activation of the Iba1-IR cells expressing P2Y12R, and blockade of this receptor suppressed the microglial activation. Taken together, these findings indicate that P2Y12 signaling is involved in microglial activation in the Vc following tongue cancer inoculation.
Hyperactive Vc nociceptive neurons and activated microglia release various molecules involved in the activation of microglia in the Vc, such as glutamate, substance P, calcitonin gene-related peptide, and ATP (Ren and Dubner 2010; Sessle 2011). Microglial activation causes further enhancement of the excitability of Vc nociceptive neurons via functional interactions between Vc neurons and microglia. We observed the strong enhancement of the BG activity of Vc WDR neurons and mechanical-evoked responses in SCC-inoculated rats. Following administration of P2Y12R antagonist, the enhancement of Vc WDR neurons as well as microglial activation was depressed in SCC-inoculated rats. These findings suggest that Vc WDR neurons receiving mechanical-evoked afferent inputs from the tongue become hyperactive associated with microglial activation via P2Y12 signaling following SCC inoculation.
Author Contributions
T. Tamagawa, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; M. Shinoda, contributed to conception, design, data analysis, and interpretation, drafted and critically revised the manuscript; K. Honda, K. Iwata, contributed to conception, design, data acquisition, and analysis, drafted and critically revised the manuscript; A. Furukawa, contributed to conception and data acquisition, drafted and critically revised the manuscript; K. Kaji, H. Nagashima, J. Chen, contributed to conception and data analysis, drafted and critically revised the manuscript; R. Akasaka, contributed to design, data acquisition, and analysis, drafted and critically revised the manuscript; B.J. Sessle, contributed to design, data analysis, and interpretation, drafted and critically revised the manuscript; Y. Yonehara, contributed to design, data acquisition, and interpretation, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
This study was supported in part by research grants from the Sato and Uemura Funds, Nihon University School of Dentistry; a grant from the Dental Research Center, Nihon University School of Dentistry; a multidisciplinary research grant (2015) and an individual research grant from Nihon University; a Grant-in-Aid for Young Scientists (B, 26861750) from KAKENHI; and grants from the MEXT-Supported Program for the Strategic Research Foundation at Private Universities (2013-2017).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.