
Abstract
This study aimed to investigate the role of PIN1 on the hepatic differentiation of human dental pulp stem cells (hDPSCs) and its signaling pathway, as well as the potential therapeutic effects of hDPSC transplantation and PIN1 inhibition on CCl4 (carbon tetrachloride)–induced liver fibrosis in mice. The in vitro results showed that hepatic differentiation was suppressed by infection with adenovirus-PIN1 and promoted by PIN1 inhibitor juglone via the downregulation of Wnt3a and β-catenin. Compared with treatment with either hDPSC transplantation or juglone alone, the combination of hDPSCs and juglone into CCl4-injured mice significantly suppressed liver fibrosis and restored serum levels of alanine transaminase, aspartate transaminase, and ammonia. Collectively, the present study shows for the first time that PIN1 inhibition promotes hepatic differentiation of hDPSCs through the Wnt/β-catenin pathway. Furthermore, juglone in combination with hDPSC transplantation effectively treats liver fibrosis, suggesting that hDPSC transplantation with PIN1 inhibition may be a novel therapeutic candidate for the treatment of liver injury.
Keywords
Introduction
Currently, cell-based therapies are emerging as an alternative to whole organ transplantation (Schuppan and Afdhal 2008; Wallace et al. 2008). Stem cell therapies based on hematopoietic stem cells, bone marrow–derived mesenchymal stem cells, and adipose tissue–derived mesenchymal stem cells have been successfully used in preclinical animal studies and clinical studies on patients with acute liver failure or chronic liver disease with end-stage cirrhosis (Kisseleva et al. 2010; Cho et al. 2011). However, methods used to harvest stem cells for these purposes are highly invasive and painful for the donor. Furthermore, the number and differentiation potential of stem cells decline with increasing age (Martens et al. 2013). Therefore, one alternative transplantable source of hepatic progenitors for the treatment of liver disease is subject to intensive investigation.
Human dental pulp contains human dental pulp stem cells (hDPSCs), which can differentiate into cells of odontogenic, adipogenic, osteogenic, chondrogenic, myogenic, and neurogenic lineages in vitro (Iohara et al. 2006). One advantage of hDPSCs is that they can be easily obtained noninvasively from teeth without any further surgical intervention with respect to the donor. The successful transplantation or engraftment of hDPSCs into adult animals, such as mice, rats, and rabbits, has been reported (de Mendonca et al. 2008; Huang et al. 2008; Gomes et al. 2010). Importantly, hDPSCs can differentiate into hepatocyte-like cells, which exhibit hepatocytic morphology (Ishkitiev et al. 2010; Ishkitiev, Calenic, et al. 2012; Ishkitiev, Yaegaki, et al. 2012). Taken together, these reports suggest that hDPSCs are an alternative source for cell therapy–based treatments of liver cirrhosis.
PIN1, or protein interacting with NIMA (never in mitosis A)–1, is a peptidyl-prolyl isomerase that can alter the conformation of phosphoproteins, thereby affecting protein function (Lu et al. 2007). PIN1 regulates a variety of cellular events for cell cycle progression, differentiation, and stress responses (Ryo et al. 2003). We recently reported that PIN1 inhibition can promote the odontogenic differentiation of hDPSCs (Lee et al. 2014) but suppress osteoclast differentiation (Cho et al. 2015).
There are many kinds of liver diseases, including hepatitis (viral infection–mediated inflammation of liver), fascioliasis (parasitic infection–mediated hepatic disease), and steatohepatitis (fatty liver disease, alcoholic/nonalcoholic). Although liver transplantation has long been regarded as an effective treatment strategy, the growing knowledge of cell biology has exploited an alternative stem cell based–cell therapy for patients with liver diseases (Vosough et al. 2011; Kadyk et al. 2015). One of the most common liver diseases, nonalcoholic steatohepatitis (NASH), is managed solely by lifestyle changes (e.g., diet and exercise) because it currently has no obvious treatment methods in spite of the increasing prevalence rate of NASH in the developed countries (Nakatsu et al. 2012).
PIN1–/– mice are highly resistant to NASH, suggesting that PIN1 inhibition could provide a novel therapeutic strategy for it (Nakatsu et al. 2012). In contrast, PIN1–/– mice exhibit more liver necrosis after ischemia/reperfusion injury when compared with wild-type mice (Kuboki et al. 2009), and PIN1 overexpression attenuates acute liver injury (liver necrosis) induced by carbon tetrachloride (CCl4) in mice (Risal et al. 2012). Although results from these studies are inconsistent depending on types of liver injury, PIN1 seems to play a critical role in liver disease and/or regeneration.
To provide a novel strategy for liver fibrosis, this study aims to investigate the role of PIN1 in the hepatic differentiation of hDPSCs with its underlying signaling pathways and to determine whether hepatocyte-like cells derived from hDPSCs and PIN1 inhibition can be useful for liver repair or regeneration in the CCl4-injured liver fibrosis of mice.
Materials and Methods
Cell Culture of hDPSCs
Human dental pulp tissues were obtained from clinically healthy permanent teeth (premolars) extracted during orthodontic treatment. All the human sample experiments were approved by the institutional review board and the research ethics committee of Kyung Hee University. hDPSCs were isolated and cultured as previously described (Ishkitiev et al. 2010; Lee et al. 2014). In brief, extracted primary cells cultured within the first 5 passages were expanded in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum with 1% antibiotic and subsequently submitted to magnetic-activated cell sorting to isolate hDPSCs. CD34+/c-kit+/STRO-1+ cells were defined as hDPSCs. To induce hepatic differentiation, primary hDPSCs were cultured in hepatogenic medium (HM) containing DMEM, 2% fetal calf serum, and 20 ng/mL of rh-hepatocyte growth factor for 5 d, followed by a mixture of 10 ng/mL of Oncostatin M, 10 nmol/L of dexamethasone, and 1% ITS-X for an additional 15 d, as described previously (Ishkitiev et al. 2010). Hepatic differentiation of hDPSCs was induced in the presence or absence of juglone or vehicle (ethanol, <1% final concentration).
Preparation of Recombinant PIN1 Adenovirus
Adenoviruses encoding PIN1 (Ad-PIN1) or β-galactosidase that were constructed with the ViraPower Adenovirus Expression System (Invitrogen) were kindly provided by Professor Byung-Hyun Park (Jeonbuk National University). Adenoviral infection was carried out at a multiplicity of infection of 100 in OPTI-MEM (Gibco BRL Co.) for 5 h before commencement of hepatogenic induction.
Periodic Acid–Schiff Staining
Cells were fixed in 4% paraformaldehyde for 3 min and then oxidized in 1% periodic acid for 5 min, washed with phosphate-buffered saline, and treated with Schiff’s reagent for 15 min. Staining intensities were determined with ImageJ software (National Institutes of Health).
RNA Isolation and Reverse Transcription Polymerase Chain Reaction
Total RNA was isolated with TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. Reverse transcription was performed with AccuPower RT PreMix (Bioneer). The resulting cDNA was amplified with AccuPower PCR PreMix (Bioneer). Primer information and polymerase chain reaction (PCR) conditions are listed in previous studies (Ishkitiev et al. 2010; Ishkitiev, Yaegaki, et al. 2012). PCR products were resolved on 1.5% agarose gels and stained with ethidium bromide.
Western Blot Analysis
Whole cell lysates and nuclear extracts were prepared. Proteins (30 µg per lane) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. Membranes were then treated with blocking solution for 1 h and incubated with primary and horseradish peroxidase–conjugated secondary antibodies for 1 h at 37 °C. Proteins were visualized with enhanced chemiluminescence (Amersham).
Immunofluorescence Staining
Cells were fixed in 4% paraformaldehyde for 30 min. Primary antibodies against stage-specific embryonic antigen-4 (SSEA-4) and STRO-1 were incubated in blocking buffer for overnight at 4 °C. Alexa Fluor 488–conjugated secondary antibody (Molecular Probes) was incubated in 1% bovine serum albumin / phosphate-buffered saline for 1 h at 4 °C. For flow cytometry, 10,000 events were acquired with a FACSCalibur apparatus (BD Biosciences). Data were analyzed with CellQuest research software (BD Biosciences). Unstained cells and cells stained only with secondary antibody served as negative controls. Cells examined by confocal microscopy were mounted on glass slides and observed with a Cell Voyager CV1000 (Yokogawa Electric Corporation).
Induction of Hepatic Fibrosis and Cell Transplantation
Five-week-old BALB/c nude mice (male) were purchased from Central Lab Animal, Inc. Experimental protocols were approved by the Ethics Committee for Animal Care and Use of Kyung Hee University, Seoul, Korea. Liver fibrosis was induced by CCl4 injection (twice weekly for 4 wk) into the peritoneum at a dosage of 1 mL/kg of body weight of CCl4 dissolved in a 1:10 volume of castor oil (Sigma-Aldrich). Chemical marker PKH26-labeled hepatocyte-like cells derived from hDPSCs (1 × 105 cells/100 µL) were transplanted into each mouse via tail vein infusion. Juglone (1 mg/kg) was injected intraperitoneally twice weekly for an additional 3 wk (Fig. 5A).
Detection of Transplanted Human Cells in Mice
To trace the transplanted hDPSCs in CCl4-injured mice, cells were labeled with a PKH26 red fluorescent cell linker kit (Sigma-Aldrich) according to the manufacturer’s instructions and observed with a Typhoon 8600 variable mode imager (GE Healthcare). The presence of grafted human cells in the livers of CCl4-injured mice was assessed by immunohistochemical examination. A Histostain-Plus rabbit primary kit (Zymed Laboratories, Inc.) was used to visualize immunoactions. Human cytokeratin 18 (CK18)–specific antibody was purchased from Santa Cruz Biotechnology (sc-51583).
Blood and Histologic Analysis
Mice from all experimental groups were sacrificed 7 wk after commencing the experiment. Blood was collected, and the concentrations of alanine aminotransferase (ALT), aspartate aminotransferase (AST), and ammonia were measured with a BioVision assay kit. Specimens were fixed in 10% neutral-buffered formalin, embedded in paraffin wax via conventional techniques, sectioned at 4-μm thickness, and stained with hematoxylin and eosin. Heidenhain’s Azan trichrome staining and periodic acid–Schiff (PAS) staining were also performed to examine liver collagen deposition and glycogen storage, respectively. Fibrosis areas were measured with ImageJ 1.44 image analysis software and a CX41 microscope (Olympus) equipped with a DP21 digital camera (Olympus). The amount of collagen deposition was calculated from 25 randomized and nonoverlapping areas at 20× magnification. The percentage of fibrosis in the liver was calculated by dividing the total blue-stained area by the total liver area.
Hydroxyproline Content Assay
Hepatic hydroxyproline content in whole liver samples was quantified colorimetrically; 30 mg of freeze-dried liver samples were hydrolyzed in 6N HCl at 110 °C for 16 h. Hydroxyproline concentration was calculated from a standard curve prepared with high-purity hydroxyproline (Sigma Aldrich) and expressed as mg/g (hydroxyproline/liver).
Statistical Analysis
Comparisons of parameters among groups were made by 1-way analysis of variance combined with Bonferroni correction. All values are presented as the means ± SD. A P value <0.05 was considered statistically significant.
Results
Characterization of the Hepatic Differentiation of hDPSCs
To characterize the hepatic differentiation of hDPSCs, cells were treated with HM for up to 21 d; then their morphology and protein expression of hepatic markers were analyzed. After 21 d of culture, hDPSCs cultured in HM changed from an elongated, spindle shape to a more cuboidal or polygonal shape (Fig. 1A). Protein levels of the early hepatic marker albumin and hepatocyte nuclear factor 1α were increased, with the highest expression 5 d after HM treatment, after which levels decreased. The expression of protein levels of the mature hepatic marker CK18 was increased in a time-dependent manner, and another mature hepatic marker, CCAAT/enhancer binding protein α, was increased up to 10 d and slightly decreased thereafter (Fig. 1B). To further confirm the expression of hepatic marker in hDPSCs, hepatic differentiated hDPSCs were examined by immunocytochemistry (Fig. 1C). Intense positive staining for albumin, hepatocyte nuclear factor 1α, CK18, and CCAAT/enhancer binding protein α were detected at 21 d of HM-induced differentiated hDPSCs.
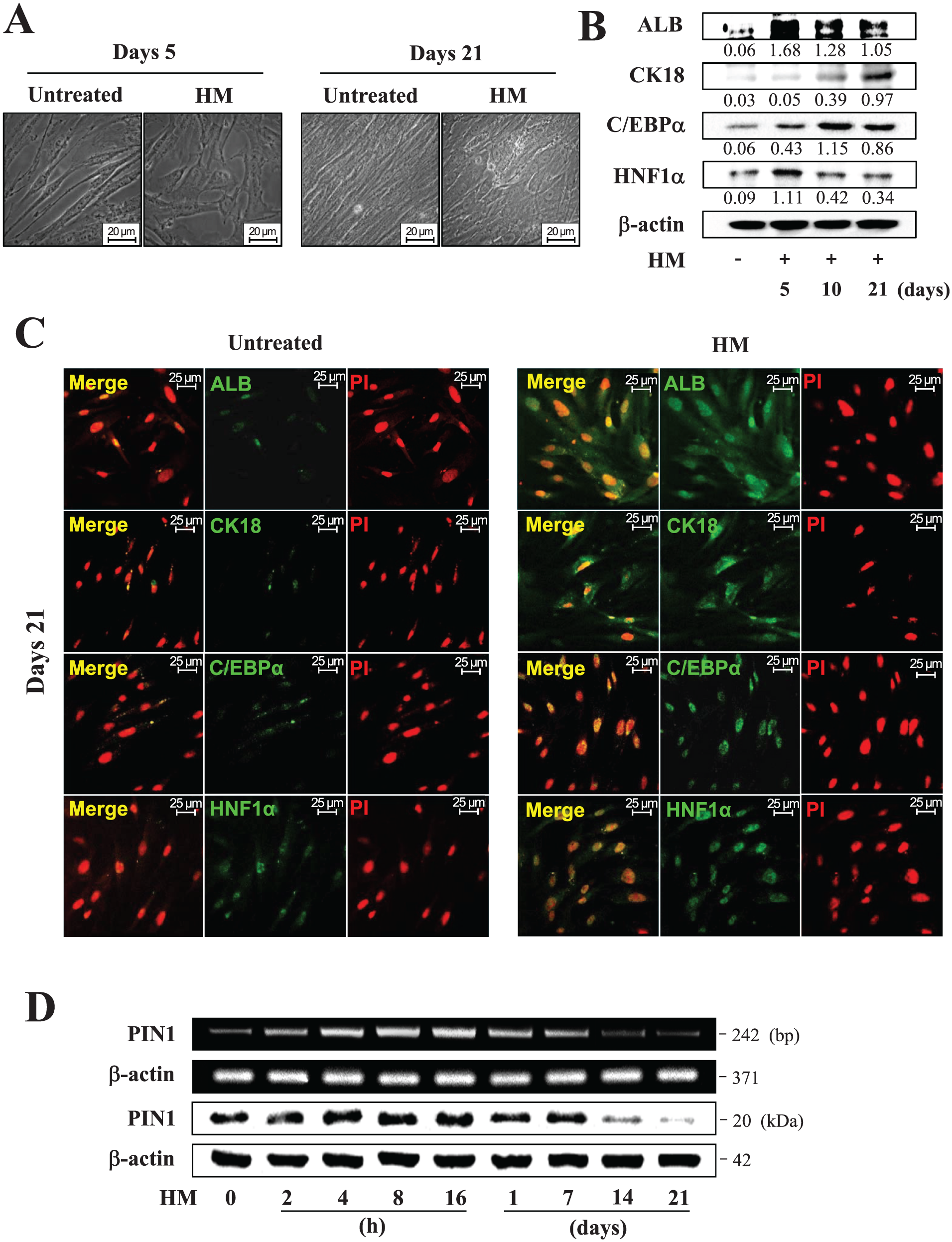
Characterization of hepatic differentiation of human dental pulp stem cells (hDPSCs). Cells were treated with hepatogenic medium (HM) for 21 d. (
PIN1 mRNA and Protein Expression during Hepatic Differentiation of hDPSCs
To investigate PIN1 mRNA and protein expression during hepatic differentiation of hDPSCs, cells were cultured in HM for 21 d and analyzed by conventional reverse transcription PCR and Western blotting (Fig. 1D). PIN1 mRNA and protein expression increased up until 16 h but gradually decreased thereafter. A rapid decrease in PIN1 was observed after 7 d.
Effects of PIN1 Inhibition and Overexpression on Hepatic Differentiation of hDPSCs
To examine the role of PIN1, juglone (5-hydroxy-1,4-naphthoquinone; a potent inhibitor of PIN1) was treated during the hepatic differentiation of hDPSCs. Juglone has been suggested as a reactive molecule that specifically inhibits the enzymatic activity of PIN1 (Lee et al. 2014; Costantino et al. 2016). Glycogen storage ability, which was assessed by PAS staining of hDPSC-derived hepatocyte-like cells, and expressions of hepatic marker were markedly stimulated by juglone in a dose-dependent manner (Fig. 2A, B). In Figure 2B, the relative expression of PIN1 upon juglone treatment was difficult to detect because the expression was markedly decreased after 7 d of hepatic differentiation (as shown in Fig. 1D). However, PIN1 shows a meaningful decrease in the presence of a high concentration of juglone (2 µM). STRO-1 and SSEA-4 are the best-known markers for mesenchymal stem cells (Yu et al. 2007; Kawanabe et al. 2012). Of note, pretreatment with juglone enhanced the HM-induced decrease of STRO-1- and SSEA-4-positive cells (Fig. 2C). On the contrary, PIN1 overexpression by Ad-PIN1 blocked HM-induced glycogen storage and downregulated mRNA expression of hepatic markers in hDPSCs (Fig. 2D, E).
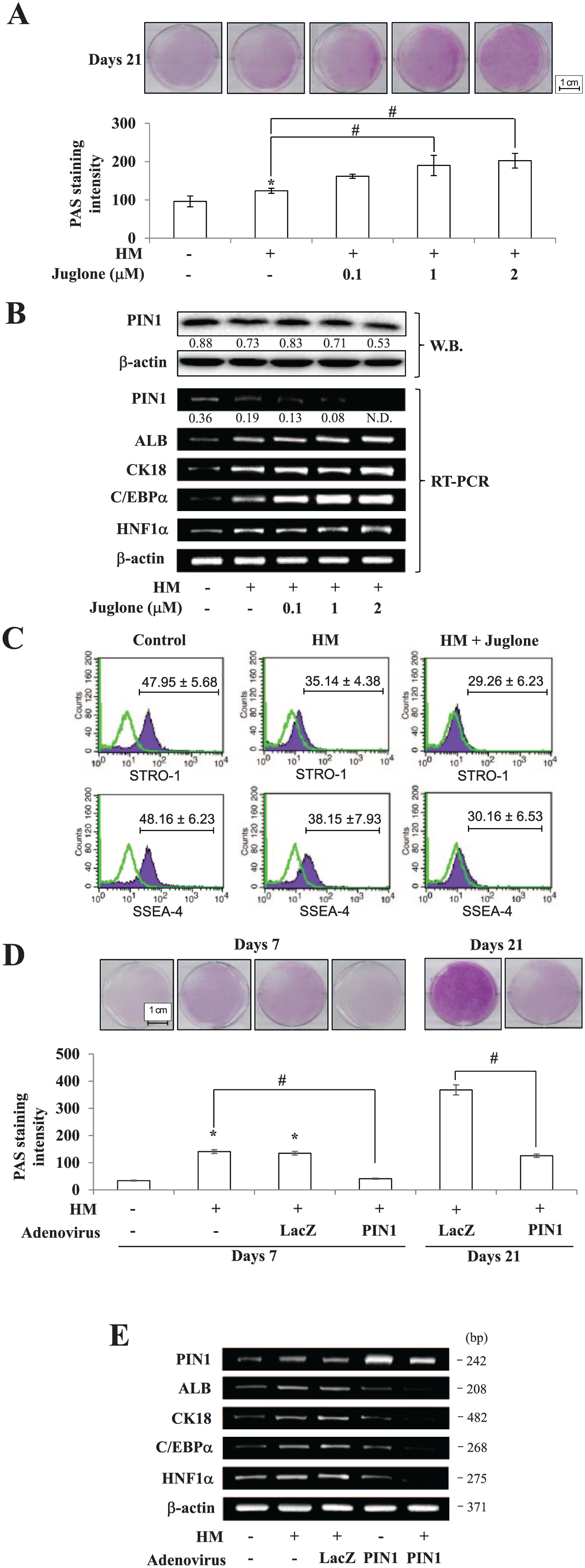
Effects of PIN1 on hepatic differentiation. To inhibit PIN1, cells were treated with juglone during the period of hepatic differentiation. Differentiation was assessed by (
Effects of PIN1 Inhibition and Overexpression on the Wnt3a/β-Catenin Pathway
To better understand the molecular mechanism of PIN1-mediated hepatic differentiation, we evaluated Wnt/β-catenin signaling in HM-treated hDPSCs upon PIN1 inhibition and overexpression. Among the Wnt genes examined, HM increased Wnt3a expression and subsequent total β-catenin protein expression in a time-dependent manner, whereas the expression of Wnt1 and Wnt5a remain unchanged (Fig. 3A). The presence of juglone further enhanced HM-mediated Wnt3a upregulation, while forced expression of PIN1 dramatically diminished the increase in Wnt3a (Fig. 3B). To further corroborate that PIN1 triggers a Wnt3a decrease during hepatocyte differentiation, PIN1-overexpressing hDPSCs were allowed to differentiate into hepatocytes in the presence or absence of rh-Wnt3a (Fig. 3C). In accordance with Western blot results, overexpression of PIN1 reduced the hepatocytic glycogen storage ability of hDPSCs, which was rescued by exogenous treatment with rh-Wnt3a. Similarly, restoration of hepatocyte marker genes upon Wnt3a treatment in PIN1-overexpressing cells further confirmed the significant role of PIN1 in Wnt3a reduction (Fig. 3D). In addition, DKK-1, an inhibitor of the Wnt signaling pathway, further enhanced Ad-PIN1-mediated inhibition of hepatic differentiation.
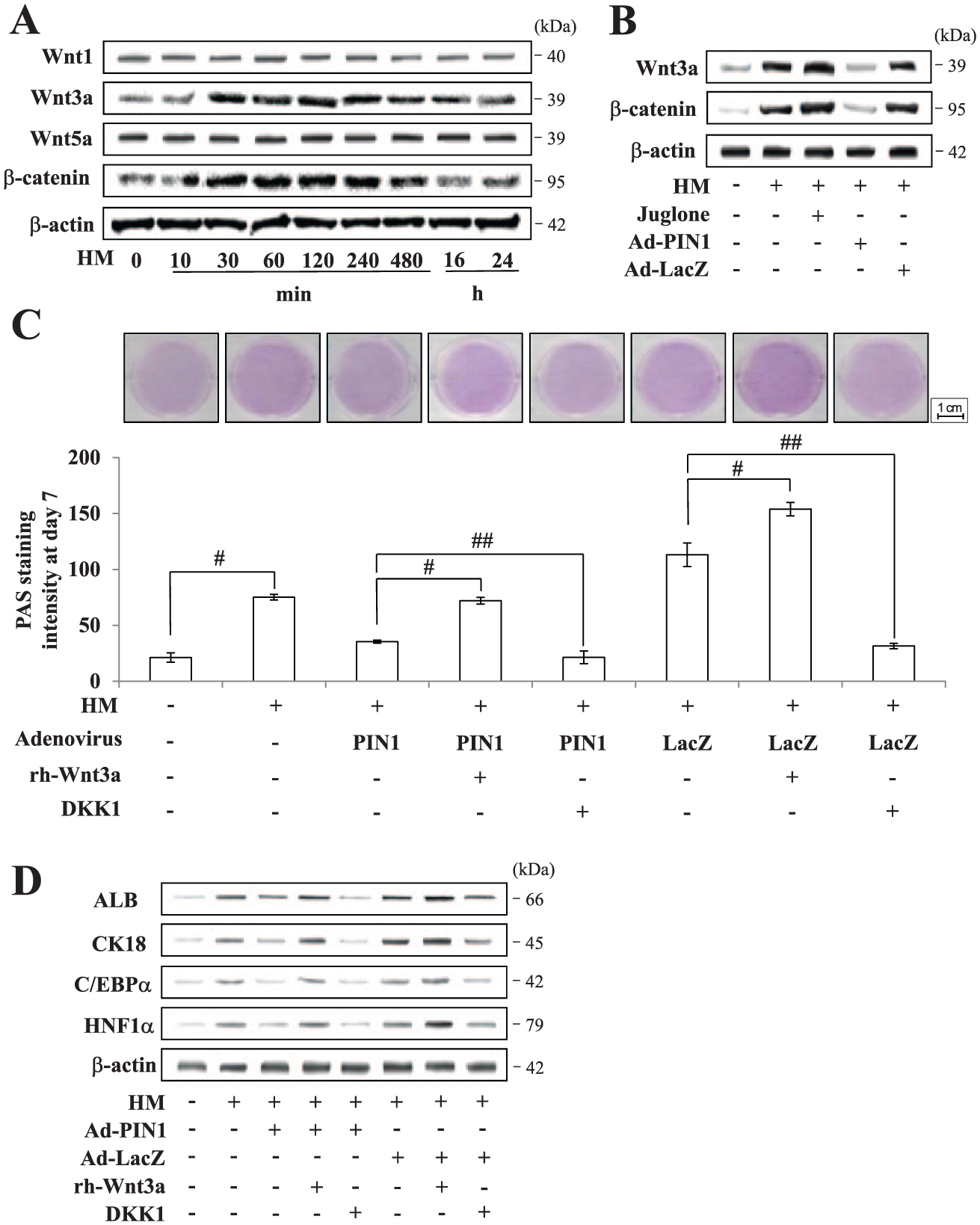
Effects of PIN1 on the Wnt3a/β-catenin pathway during hepatic differentiation in human dental pulp stem cells. (
Confirmation of Cell Engraftment in the CCl4-Injured Mouse Liver
To determine whether human cells were present in the livers of transplanted mice, PKH26-labeled hDPSCs were transplanted into CCl4-injured mice (Fig. 4A). PKH26-derived fluorescence was not detected in the cryosectioned liver that did not receive transplanted cells. In contrast, significant fluorescence was observed in the liver receiving hDPSCs (Fig. 4B). The distribution of human cells in mouse liver tissue was further confirmed by immunohistochemistry for antihuman major histocompatibility complex class I (Fig. 4C). Furthermore, we performed additional immunohistochemistry using the antibody that detects human but not mouse proteins. Human-specific CK18 was positively stained in the liver tissue of the mice injected with hDPSCs (Fig. 4D). Therefore, we conclude that the differentiated hDPSCs act as hepatocytes in the murine liver after cell transplantation.
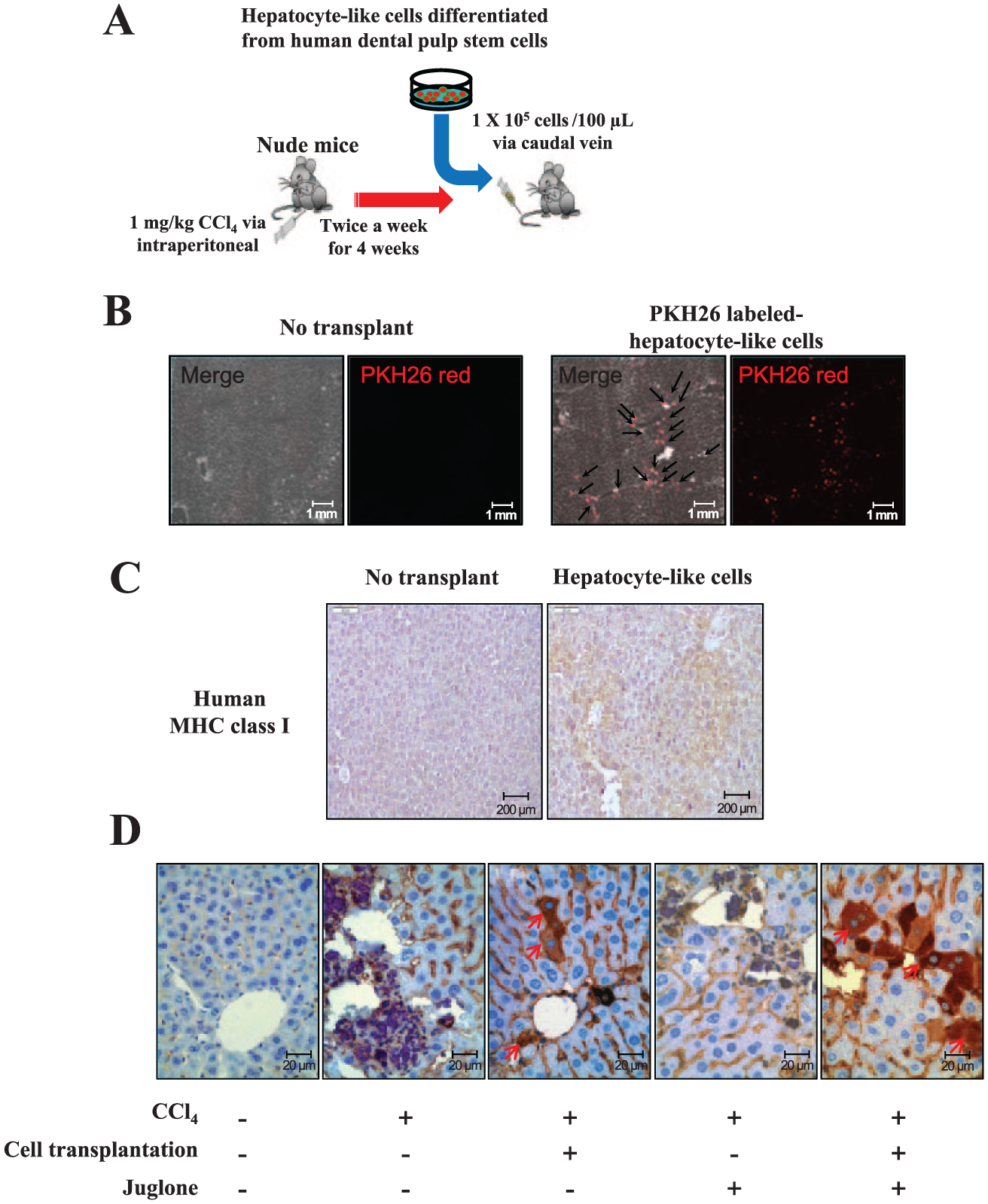
Schematic diagram of an animal model for hepatic fibrosis and cell transplantation. (
In Vivo Therapeutic Effects of hDPSC Transplantation versus hDPSCs Plus Juglone into Mice with Liver Fibrosis
Since PIN1 inhibition by juglone promoted hepatic differentiation in hDPSCs, we next investigated the effects of juglone and hepatocyte-like cell transplantation in CCl4-induced liver-injured mice by histologic examination and determined the serum levels of ALT, AST, and ammonia (Fig. 5A). Following 3 wk of treatment with CCl4, liver fibrosis and destruction of sinusoidal structures were observed in sections stained with hematoxylin and eosin, PAS, and Heidenhain’s Azan trichrome (Fig. 5B). Blue-stained (fibrotic) areas decreased in mouse livers following hDPSC engraftment or juglone treatment versus treatment with CCl4 alone. Histomorphometric examination of liver tissues showed that juglone in combination with hDPSC engraftment has a significant antifibrotic effect, as evidenced by a decrease in liver fibrosis versus the juglone-alone or hDPSC engraftment–alone group (based on Azan trichrome staining; Fig. 5C). Serum ALT, AST, and ammonia levels significantly increased in response to CCl4 as compared with the controls but were significantly reduced in mice treated with either juglone or hDPSCs alone (Fig. 5D–F). Combined treatment of both juglone and hDPSCs significantly decreased CCl4-induced increase of ALT, AST, and ammonia. Furthermore, hDPSC engraftment plus juglone treatment have a significant antifibrotic effect, as evidenced by the significant decrease in liver collagen (hydroxyproline content) as compared with CCl4, CCl4 plus juglone alone, and CCl4 plus hDPSCs alone (Fig. 5G).
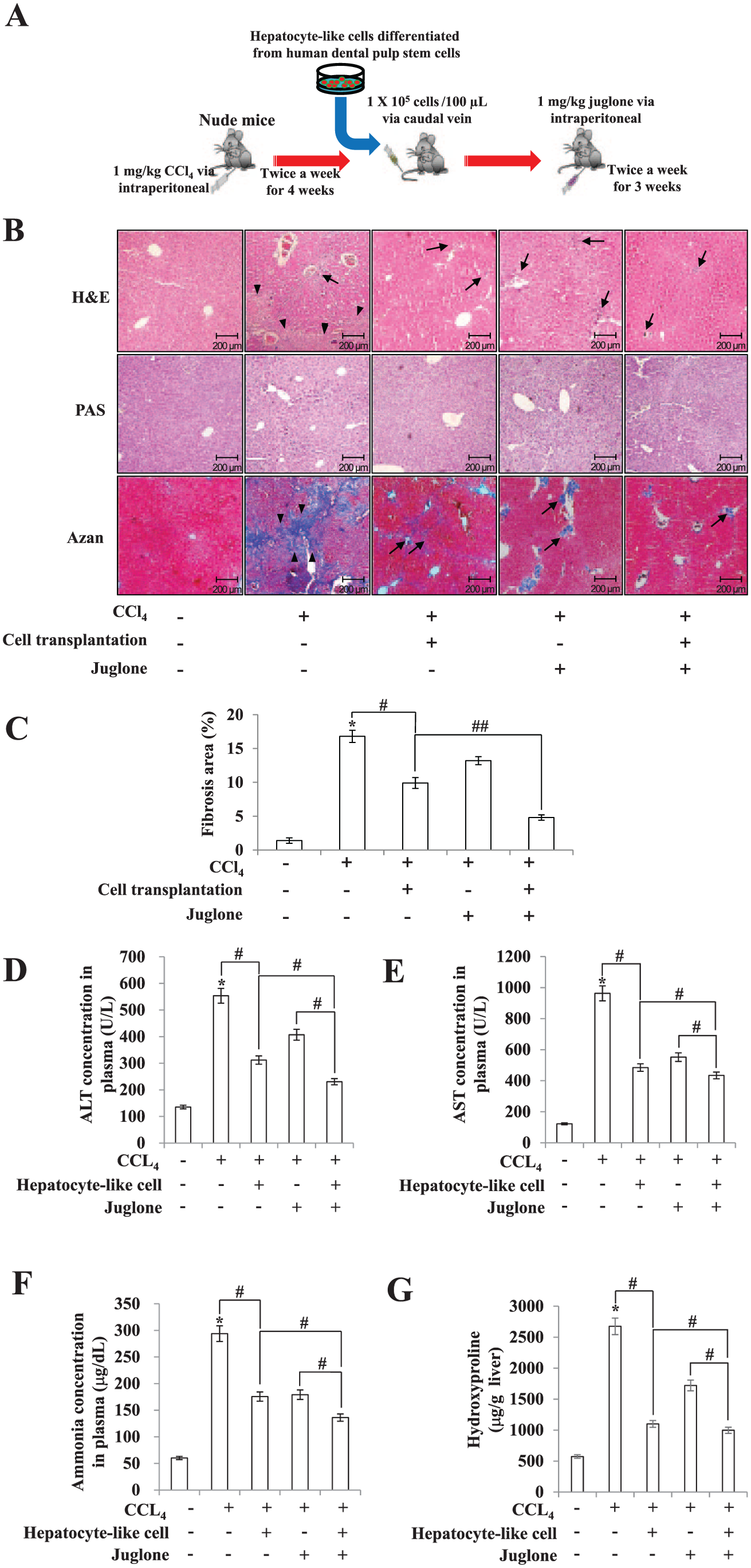
Therapeutic effects of human dental pulp stem cell transplantation and juglone in carbon tetrachloride (CCl4)–induced liver fibrosis in nude mice. (
Discussion
Consistent with findings from previous studies, we report that hDPSCs can differentiate into cells that exhibit a hepatic phenotype in 20 to 22 d (Ishkitiev et al. 2010; Patil et al. 2014), and we verified that hDPSCs cultured in HM express high mRNA and protein levels of hepatic markers and contain PAS-positive glycogen granules. The present study also showed that PIN1 mRNA and protein expression increased in a time-dependent manner up until 16 h following hepatic induction but began to decline thereafter and was consequently suppressed for the remainder of hepatic differentiation of hDPSCs.
Wnt/β-catenin signaling is known to be a major player in the hepatocyte/liver developmental process (Thompson and Monga 2007; Monga 2014). In this study, expression levels of Wnt3a and β-catenin were enhanced in the presence of juglone but inhibited in the presence of Ad-PIN1, which is consistent with our report on the odontogenic differentiation of hDPSCs (Lee et al. 2014). In addition, the addition of rh-Wnt3a significantly reversed the inhibitory effect of Ad-PIN1on hepatic differentiation. These data support a role for PIN1 in the negative regulation of hepatocyte differentiation by inhibiting Wnt3a expression.
Engraftment is a multistep process involving directed migration of inoculated cells. The fluorescent dye PKH26, which binds mainly to the cell membrane, has been used as a cell tracer to locate transplanted cells in hosts (Li et al. 2013). In the present study, hDPSC engraftment in livers of immunocompromised mice was demonstrated by the presence of positive cells for PKH26, human major histocompatibility complex class I, and human CK18 at 3 wk posttransplantation. Our findings indicate that hDPSCs can migrate to the liver injury site and participate in liver injury healing by differentiating into hepatocytes. However, there could be another mechanism of liver injury healing by hDPSCs. Besides differentiating into hepatocytes, hDPSCs could induce hepatic differentiation of liver stem cells in the recipient mice, or hDPSCs could provide some kind of cytokines that promote liver injury healing. The precise underlying mechanism of hDPSC-mediated liver injury healing needs to be investigated in further studies.
Besides hDPSCs, several types of stem cells were evaluated as a transplantable cell sources for stem cell–based liver therapy: bone marrow–derived mesenchymal stem cells, hematopoietic stem cells, and adipose tissue–derived mesenchymal stem cells (Vosough et al. 2011; Kadyk et al. 2015). Each stem cell has its own characteristic advantages and limitations for cell-based therapy of liver repair. One of the greater advantages of DPSCs than other stem cells will be its simple and noninvasive obtaining protocol. The role of PIN1 on hepatocyte-like differentiation of other stem cells was not fully comprehended. However, recent studies report the resistance of PIN1 knockout mice to NASH (Nakatsu et al. 2012) and the attenuation of hepatocellular carcinoma upon PIN1 knockdown (Shinoda et al. 2015). These reports suggest the relevance of PIN1 to liver homeostasis, although further studies are required to understand the precise mechanisms.
Experimental liver injury by CCl4 in nude mice is a widely used animal model to examine hepatic fibrosis by intraperitoneal administration (Cho et al. 2011). As a result of hepatic injury, liver fibrosis, the number of glycogen-laden cells, hepatic hydroxyproline contents, and serum ALT, AST, and ammonia levels showed a marked increase after CCl4 administration; however, these increases were attenuated by hDPSCs or the PIN1 inhibitor juglone. Furthermore, the combination of hDPSCs and juglone reduced CCl4-induced fibrosis scores. Taken together, these data demonstrate that cotreatment with hDPSC transplantation and juglone effectively reduces experimental hepatic fibrosis.
In summary, this study is the first to demonstrate that PIN1 inhibition promotes hepatic differentiation of hDPSCs via the Wnt/β-catenin pathway. Moreover, a combination therapy of hDPSC transplantation with juglone demonstrates additive effects on CCl4-induced hepatic fibrosis. Thus, the combination of hDPSC transplantation and PIN1 inhibition may represent a potential new therapeutic approach for the treatment of hepatic fibrosis and cirrhosis.
Author Contributions
H.J. Kim, contributed to design and data interpretation, drafted the manuscript; Y.A. Cho, Y.M. Lee, S.Y. Lee, and W.J. Bae, contributed to conception and data acquisition, drafted the manuscript; E.C. Kim, contributed to conception, design, data acquisition, and analysis, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Footnotes
This work was supported by a grant from the National Research Foundation of Korea, funded by the Ministry of Science, ICT and Future Planning (2012R1A5A2051384), and by a Midcareer Researcher Program grant through the National Research Foundation of Korea, funded by the Ministry of Education, Science and Technology (2012004117).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.