
Abstract
The aim of this study was to gain insights into the biology and mechanics of immediate postextraction implant osseointegration. To mimic clinical practice, murine first molar extraction was followed by osteotomy site preparation, specifically in the palatal root socket. The osteotomy was positioned such that it removed periodontal ligament (PDL) only on the palatal aspect of the socket, leaving the buccal aspect undisturbed. This strategy created 2 distinct peri-implant environments: on the palatal aspect, the implant was in direct contact with bone, while on the buccal aspect, a PDL-filled gap existed between the implant and bone. Finite element modeling showed high strains on the palatal aspect, where bone was compressed by the implant. Osteocyte death and bone resorption predominated on the palatal aspect, leading to the loss of peri-implant bone. On the buccal aspect, where finite element modeling revealed low strains, there was minimal osteocyte death and robust peri-implant bone formation. Initially, the buccal aspect was filled with PDL remnants, which we found directly provided Wnt-responsive cells that were responsible for new bone formation and osseointegration. On the palatal aspect, which was devoid of PDL and Wnt-responsive cells, adding exogenous liposomal WNT3A created an osteogenic environment for rapid peri-implant bone formation. Thus, we conclude that low strain and high Wnt signaling favor osseointegration of immediate postextraction implants. The PDL harbors Wnt-responsive cells that are inherently osteogenic, and if the PDL tissue is healthy, it is reasonable to preserve this tissue during immediate implant placement.
Introduction
In the past, a standard dental implant procedure consisted of tooth removal, followed by extraction site healing, then osteotomy site preparation and subsequent implant placement (Branemark et al. 1977). This strategy was thought to ensure maximum bone-implant contact (BIC) because the dimensions of the osteotomy (e.g., produced by drilling) could be tailor-fit to the implant (Heimke 1990). Patient demand for shorter treatment times, however, led to an increase in the use of implants that are placed immediately after tooth extraction, without a period of extraction socket healing (Hong and Oh 2017), and an obvious difference between an osteotomy and an extraction socket is that the latter has an irregular geometry. Upon implant placement, this irregular geometry creates some regions of BIC but also some regions where there are gaps between the bone and implant. Clinical data demonstrate that immediate postextraction implants can undergo osseointegration (Paolantonio et al. 2001; Anitua et al. 2016; Buser et al. 2017), but the biomechanical mechanisms responsible for this success are not entirely clear. Using a well-characterized mouse model of oral implant placement (Mouraret et al. 2013; Mouraret et al. 2014; Wang et al. 2017; Yin et al. 2016), we sought to gain insights into the unique biomechanical environments around such “immediate” implants to determine if there were methods by which immediate postextraction implant osseointegration could be improved.
In a previous study, we demonstrated that extraction sockets heal significantly faster than osteotomies (Pei et al. 2017), and the underlying reason for this faster repair was due to Wnt-responsive osteoprogenitor cells residing in the periodontal ligament (PDL) that remained attached to the socket wall after tooth extraction (Yuan et al. 2018). Osteotomies also heal, but the early phase of repair takes longer (Wang et al. 2017; Li et al. 2017; Pei et al. 2017), presumably because of the paucity of this Wnt-responsive osteoprogenitor population. Therefore, we set out to directly test whether Wnt-responsive cells in residual PDL might be a major factor contributing to osseointegration of immediate postextraction implants.
This was the starting point for our study. We also sought to characterize the mechanical environments and biological responses at the 2 distinctive environments created when placing an implant in an extraction socket—for example, the sites where there is a direct bone-implant interface and the sites where there is a gap interface between the bone and implant that may be populated by PDL remnants. In doing so, we gained critical insights into how gap interfaces with remnants of a healthy PDL are the sites of robust osteogenesis. This osteogenic environment is created in part because of low strains, coupled with the presence of Wnt-responsive cells that are poised to produce bone. We then demonstrated that the addition of a liposomal formulation of a WNT protein could stimulate the Wnt-responsive osteoprogenitor population further to accelerate new bone formation around immediate postextraction implants.
Methods and Materials
Animals and Surgery
This study complied with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines, and the Stanford Committee on Animal Research approved all protocols (No. 13146). Axin2CreERT2/+;R26RmTmG/+ mice (Nos. 018867 and 007576, respectively) and Axin2LacZ/+ mice (No. 11809809) were purchased from Jackson Laboratories and housed in a temperature-controlled environment with 12-h light/dark cycles.
Tamoxifen delivery, bilateral maxillary first molar extractions, liposomal WNT3A (L-WNT3A) delivery, and in vivo bone labeling are described in the Appendix. For sample preparation, staining, histomorphometric measurements, and finite element model, also see the Appendix.
Statistical Analyses
Results are presented as the mean ± SD values of independent replicates. Student’s t test was used to quantify differences described in this article. P ≤ 0.05 was significant.
Results
Around Immediate Postextraction Implants, Gap Interfaces Constitute a Low-Strain Pro-osteogenic Environment
To place an immediate postextraction implant, a maxillary first molar was extracted; then, on the palatal aspect of the palatal root socket, an osteotomy was produced and an implant was placed (Appendix Fig. 1). This procedure intentionally created 2 types of interfaces: one interface allowed for direct BIC, and the other had a gap between the implant and bone, which in some cases contained remnants of PDL (asterisks, Fig. 1A).
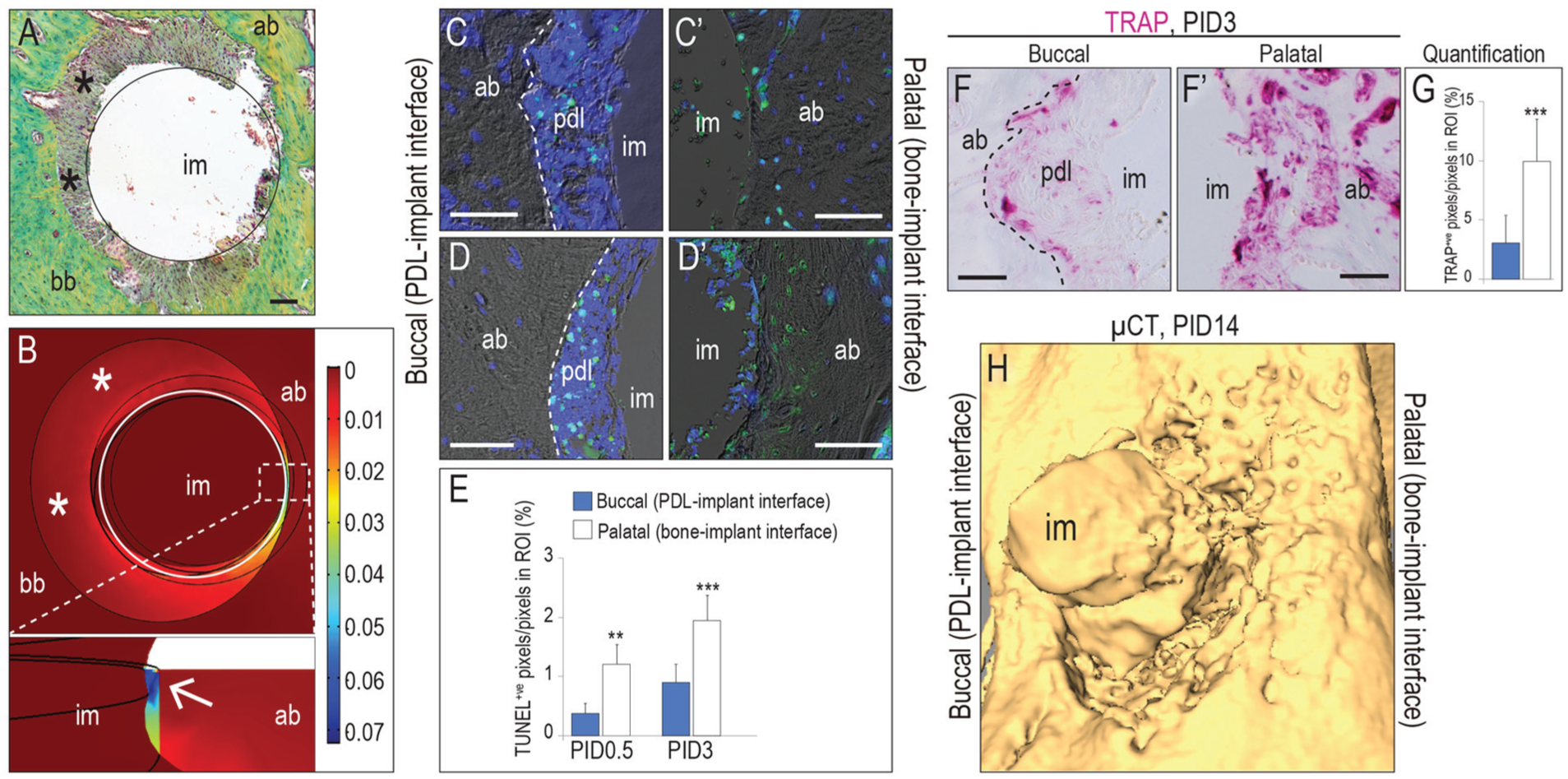
Around immediate postextraction implants, gap interfaces constitute a low-strain pro-osteogenic environment. (
We used finite element modeling to map the distribution of strains in the peri-implant environment. These strains were produced by the insertion torque (~0.1 N·cm) used during implant placement (Cha et al. 2015; Wang et al. 2017; Chen et al. 2017) and by loading. All implants were positioned subocclusally, at the height of the gingiva (Appendix Fig. 1D); therefore, the finite element model assumed nominal (nonmasticatory) levels of loading (Brunski 1999; Cha et al. 2015; Dorogoy et al. 2017). Computational data demonstrated low strains on the buccal aspects of implants, coinciding with the regions where PDL-filled gaps existed (asterisks, Fig. 1B). However, high strains existed on the palatal aspects of the implants where the bone and implant were in direct contact (arrow in magnified area, Fig. 1B).
The biological sequelae of these strain distributions were assessed on postimplant day (PID) 0.5 with TUNEL and DAPI to detect dead and dying osteocytes. On the buccal aspect, the only apoptotic cells detected were in the PDL remnants (Fig. 1C), whereas on the palatal aspect, apoptotic osteocytes were plentiful (Fig. 1C′). We also assessed the distribution of dead and dying osteocytes on PID3; again, on buccal aspects, apoptosis was limited to cells in the PDL remnants (Fig. 1D), whereas abundant apoptotic osteocytes were detected on the palatal aspects (Fig. 1D′; quantified in Fig. 1E, n = 6). These data established a direct relationship between high interfacial strains and distribution of dying and dead osteocytes around an immediate postextraction implant.
Necrotic bone triggers bone resorption (Jilka et al. 2013). In keeping with this observation, the PDL-implant interface exhibited minimal tartrate-resistant acid phosphatase (TRAP) activity on PID3, and that TRAP activity was restricted to the PDL remnants (Fig. 1F). At the bone-implant interface, however, TRAP activity was detectable throughout in the alveolar bone (Fig. 1F′; quantified in Fig. 1G, n = 6). These early differences in the pattern of bone resorption had longer-term consequences: Image analyses on PID14 indicated that relative to buccal surfaces where PDL remnants were preserved, palatal surfaces where the implant was in direct contact with bone showed significant crestal recession (Fig. 1H). Conversely, these data demonstrated that peri-implant sites with lower interfacial strain had significantly less osteocyte death and thus less bone resorption and bone recession.
Osteoprogenitor Cells Residing in the PDL Support Peri-implant Bone Formation
Low peri-implant strain environments are known to support bone formation (Cha et al. 2015). To test whether the same relationship existed around immediate postextraction implants, we evaluated buccal and palatal aspects of the implants (Fig. 2A) at a time point when new bone is detectable in the peri-implant environment. For example, on PID7, osteogenesis was underway on the buccal aspects of implants where PDL-implant interfaces had existed (n = 8; Fig. 2B). On the palatal aspects (which had been the bone-implant interface), only fibrous interfacial tissue was evident (n = 8; Fig. 2B′). Alkaline phosphatase activity was robust on the buccal aspects (Fig. 2C) and minimal on the palatal aspects (Fig. 2C′). Immunostaining for osterix (Fig. 2D, D′) and type I collagen (Fig. 2E, E′) proteins demonstrated that the former PDL-implant interfaces were populated by osteoprogenitor cells at a time when bone-implant interfaces were still lacking them.

Osteoprogenitor cells residing in the periodontal ligament (PDL) support peri-implant bone formation. (
Analyses on PID14 confirmed that the PDL-implant interface was more osteogenic than the bone-implant interface: the buccal surfaces of implants were surrounded by mature bone (n = 8; Fig. 2F), while on palatal surfaces, small gaps existed between the bone and inner diameter of the implant that filled with fibrous tissue (n = 8; Fig. 2F′). Picrosirius red staining confirmed more mature bone on buccal aspects (Fig. 2G) as compared with palatal aspects of the implants (Fig. 2G′). Osteocalcin+ve cells were embedded in the mature bone matrix at former PDL-implant interfaces (Fig. 2H). At former bone-implant interfaces, however, osteocalcin+ve cells were found associated only with preexisting alveolar bone (Fig. 2H′).
Vital dye double-labeling experiments were used to validate new bone formation. Calcein was delivered 1 d before the implant placement surgery, followed by alizarin red on PID12, 2 d prior to sacrifice. Amorphous calcein labeling was observed in the preexisting alveolar bone (Fig. 2I, I′), regardless of whether the buccal or lingual surfaces were examined. The pattern of alizarin red label, however, was different: new alizarin red–labeled interfacial bone consistently demonstrated new bone formation at former PDL-implant interfaces (Fig. 2I), whereas at former bone-implant interfaces, the new alizarin red–labeled bone was not yet juxtaposed to the implant (Fig. 2I′).
Progeny of a Wnt-Responsive Population Is Responsible for Peri-implant Bone Formation
A Wnt-responsive population associated with PDL remnants after tooth extraction contributes to socket healing (Yuan et al. 2018). We hypothesized that this Wnt-responsive population also contributes to osseointegration of an implant placed in a fresh extraction socket. To identify and follow the fate of these Wnt-responsive cells during osseointegration, we employed a Wnt reporter stain, Axin2CreErt2/+;R26mTmG/+ (van Amerongen et al. 2012). In this line, exposure to tamoxifen induces a genetic recombination event that results in a subset of Wnt-responsive cells expressing green fluorescent protein (GFP). The recombination event occurs only when tamoxifen is present; after tamoxifen has been cleared from the body, any increase in the number of GFP+ve cells reflects an expansion in the original Wnt-responsive population.
Using this strategy, we labeled a subset of Wnt-responsive cells in the intact PDL (n = 4; Fig. 3A). A similar distribution of GFP+ve cells were found around immediate postextraction implants (n = 6; Fig. 3B). Most GFP+ve cells coexpressed periostin, regardless of whether they were part of a functional PDL (Fig. 3C) or were in the PDL remnants around immediate postextraction implants (Fig. 3D).

Progeny of a Wnt-responsive population is responsible for peri-implant bone formation. (
The most significant difference between the population of GFP+ve cells in the intact PDL versus the GFP+ve population around an implant was that in the latter case, the cells were mitotically active. Few, if any, Wnt-responsive cells were actively proliferating in the intact PDL, as shown by the lack of Ki67/GFP coimmunostaining (Fig. 3E), whereas Ki67-immunopositive cells were abundant around the immediate implants (Fig. 3F). Therefore, damage to the PDL provoked a strong repair response in the Wnt-responsive population.
This mitotic activity resulted in a rapid expansion of GFP+ve population by PID7 (Fig. 3G). GFP+ve cells were largely restricted to the buccal aspects and virtually nonexistent on the palatal aspects of the same implants (Fig. 3G′). At this point, the GFP+ve population primarily consisted of osteoblasts, as demonstrated by osterix and osteocalcin expression (Fig. 3H, I). Compared with their high expression levels on the buccal aspects, these osteogenic proteins were expressed at very low levels on the palatal aspects of the same immediate postextraction implants (Fig. 3H′, I′). These data collectively signaled that bone formation occurred first in the low-strain pro-osteogenic regions where Wnt-responsive cells and their progeny were plentiful.
By PID14, on the buccal aspects of the implant, the GFP+ve osteocalcin+ve population was largely encased in mineralized matrix (Fig. 3J, K). On the palatal aspects, a GFP+ve osteocalcin+ve population was now in evidence (Fig. 3J′, K′) but not yet surrounded by a collagenous matrix. Histologically, the buccal aspects of the implants were encased in mature (yellow) bone matrix (Fig. 3L), while the bone on the palatal aspects was still in an immature (green/blue) state of mineralization (Fig. 3L′). Collectively, molecular profiling coupled with lineage tracing data demonstrated that a PDL-derived, Wnt-responsive population directly contributed to the osseointegration of an immediate postextraction implant.
Wnt Signaling Accelerates Peri-implant Bone Formation
In our experimental model, primary stability was provided by the bone-implant interface, and yet this region was also the slowest to show evidence of new bone formation. We wondered if, by increasing Wnt signaling around the entire implant, we could accelerate osseointegration on all aspects of an immediate postextraction implant. To test this possibility, we delivered a stabilized form of L-WNT3A protein or a control liposome formulation containing phosphate-buffered saline (L-PBS) to the extraction socket prior to implant placement.
On PID3, compared with L-PBS-treated implants, more Wnt-responsive cells were found in the peri-implant environment around immediate postextraction implants treated with L-WNT3A (n = 8; compare Fig. 4A, A′), which verified that the L-WNT3A was bioactive. The L-WNT3A stimulus protected osteocytes from programmed cell death, since minimal apoptosis was detected (compare Fig 4B, B′; Appendix Fig. 2). TRAP activity was also diminished by the WNT3A stimulus (Fig. 4C, C′), in keeping with the relationship between dying osteocytes and resorption of necrotic bone. Alkaline phosphatase activity was also stronger around L-WNT3A-treated implants (Fig. 4D, D′).
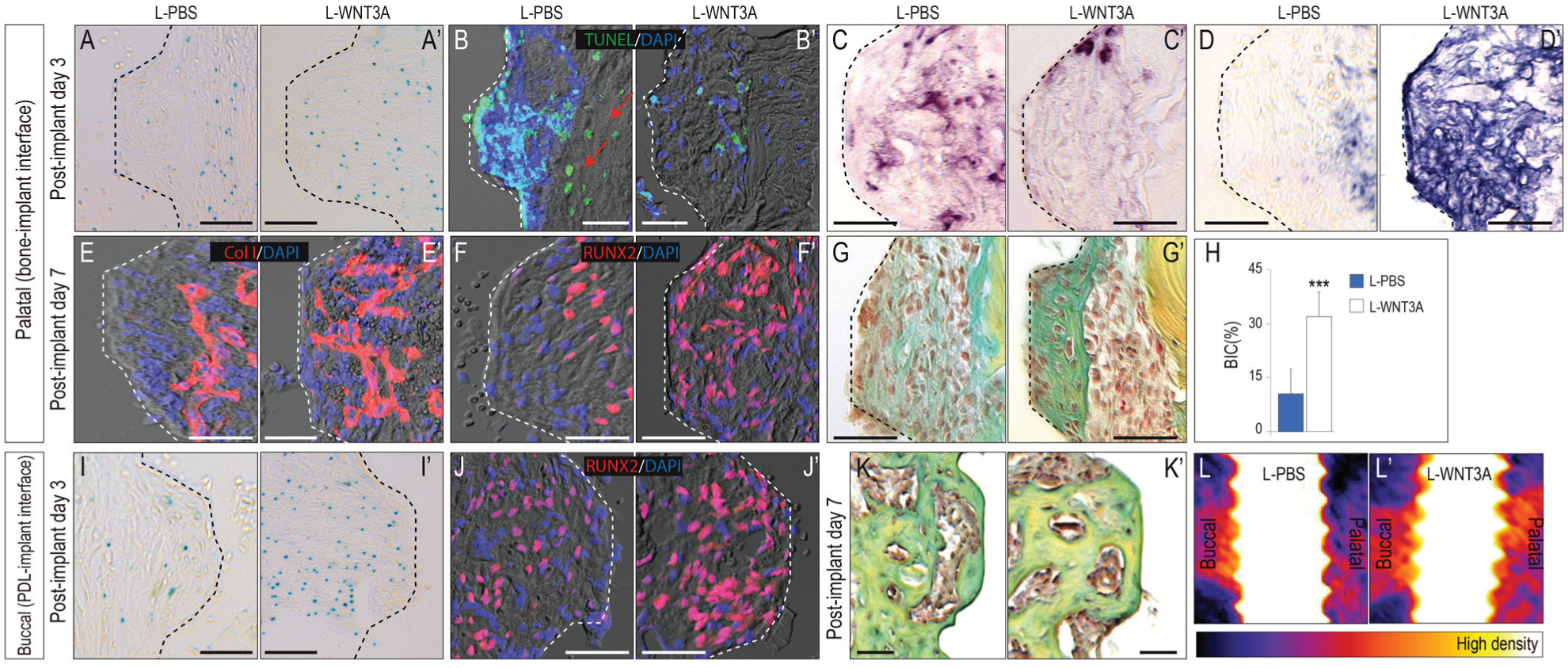
Wnt signaling accelerates peri-implant bone formation. On the palatal aspects of implants, (
New bone formation in an extraction socket is typically evident by day 7 (Yuan et al. 2018). On PID7, type I collagen expression and Runx2+ve cells were minimal on the palatal aspects in L-PBS group (Fig. 4E–G). With L-WNT3A treatment, however, type I collagen expression and Runx2+ve cells were abundant at the interface between bone and implant, with new bone formed at the bone-implant interface (Fig. 4E′–G′). On PID7, the BIC was 9% in the L-PBS group yet 32% in the L-WNT3A group (Fig. 4H). These data demonstrated that the bone formation at the bone-implant interface could be accelerated by adding exogenous L-WNT3A.
On the buccal aspects of implants, which already exhibited superior bone formation rates, L-WNT3A accelerated osseointegration still further, by increasing Xgal+ve, Runx2+ve osteogenic cell number (Fig. 4I, I′, J, J′) and promoting bone formation (Fig. 4K, K′). Imaging analyses with micro–computed tomography confirmed superior peri-implant bone formation in the L-WNT3A group on buccal and palatal aspects (Fig. 4L, L′). Overall, our data demonstrated that a low-strain pro-osteogenic environment exists around immediate postextraction implants, specifically at the sites where a gap exists between the implant and bone. This pro-osteogenic environment can be created by preserving the healthy PDL remnants or re-created by adding exogenous L-WNT3A at the time of implant placement.
Discussion
The environment around an immediate postextraction implant is unique from that which exists around a “delayed” implant—for example, one that is placed into an osteotomy that has been prepared in a healed extraction site. From a biological perspective, the former peri-implant environment contains PDL remnants, while the latter is devoid of them (Pei et al. 2017; Yuan et al. 2018). These PDL remnants are important because, at least in a healthy state, they directly contribute cells for peri-implant osseointegration (Figs. 2, 3; see Pei et al. 2017).
From a mechanical perspective, peri-implant compressive strains are created by implant insertion (Fig. 1); coupled with heat transfer during drilling, a zone of apoptosis is produced that circumscribes an implant (Cha et al. 2015). The same happens around some regions of an immediate postextraction implants—for example, at those aspects of the implant surface that are in direct contact with bone (Fig. 1). Those aspects of the implant that are facing the gap, however, experience very low strain, and as a consequence, osteocyte death is limited. When osteocyte death is limited, so is bone resorption, which is why the buccal aspects of immediate postextraction implants showed minimal cavitation. On the palatal aspects where strains were high, osteocyte death was widespread, and a bony void was common with robust osteoclastic activity. Collectively, these results suggest that as a clinical practice, producing a low-strain environment and retaining healthy PDL remnants after tooth extraction will contribute to faster osseointegration of an implant.
PDL Remnants Left Behind after Tooth Extraction Contain Osteoprogenitor Cells That Support Osseointegration of Immediate Postextraction Implants
PDL harbors Wnt-responsive osteoprogenitor cells that are responsible for healing an extraction socket. The act of extraction causes these cells to switch from a quiescent to a rapidly proliferative state and then differentiate into osteoblasts to produce new alveolar bone matrix (Yuan et al. 2018). Here, we show that this same Wnt-responsive population is also directly responsible for the osseointegration of implants placed in the tooth extraction socket (Fig. 3).
The clinical implication of these data is that the PDL should be preserved during tooth extraction and implant placement because it harbors cells that can directly contribute to bone formation and/or osseointegration, but there is a caveat: in our study, mice had healthy PDLs, whereas in clinical practice, tooth extractions are often associated with periodontal disease. If the PDL is unhealthy, is it still worth retaining it as a source of potential osteoprogenitor cells? Some studies suggested that periodontitis impedes osteogenic differentiation of PDL cells (Liu et al. 2011; Liu et al. 2015). Whether this holds true in vivo after removal of the affected tooth is not known, and despite their many advantages, rodent models are not readily amenable to this line of inquiry. The murine genomic response to inflammation appears to correlate poorly with analogous patient conditions (Seok et al. 2013). Other animal models whose chronic inflammatory responses are more similar to humans may be needed to address this question.
Wnt Signaling Prevents Osteocyte Apoptosis and Promotes Osteogenic Differentiation
Heat and strain together result in a zone of osteocyte apoptosis. Osteocyte death triggers bone resorption, which around an implant leads to a dip in implant stability. The sites where PDL remnants were retained were remarkably free of osteocyte apoptosis; instead, the only cell death observed was restricted to the pliable PDL (Fig. 1). The presence of the PDL remnants signaled that the alveolar bone had been subjected to neither drilling nor compression-induced strain. Both factors contributed to the viable status of the peri-implant bone.
In cases where PDL remnants must be removed, we found that pretreatment of the extraction socket with L-WNT3A maintained osteocyte viability (Fig. 4). Other investigators reported that increased Wnt/β-catenin signaling is associated with decreased osteocyte apoptosis (Bonewald and Johnson 2008). Moreover, WNT proteins demonstrated an antiapoptotic effect on osteoblast progenitors and differentiated osteoblasts through Scr/ERK and phosphatidylinositol 3-kinase/AKT pathway (Almeida et al. 2005). Investigators also showed that lithium chloride, which can elevate Wnt signaling, led to the differentiation of human PDL fibroblasts along an osteogenic lineage (Heo et al. 2010). Furthermore, in an osteoporotic rat model, antisclerostin therapy, which can elevate Wnt signaling, enhanced BIC by stimulating bone formation and inhibiting bone resorption (Virdi et al. 2015). These studies used an indirect method to activate Wnt signaling; nonetheless, both showed enhanced bone formation. Here, we used the WNT ligand itself, which has advantages when it comes to specificity and potency (Minear et al. 2010; Dhamdhere et al. 2014; Yin et al. 2016). The mechanism of L-WNT3A action on osseointegration (Fig. 4) may be to convert PDL fibroblasts directly into osteoprogenitor cells, and/or it may function to recruit cells to the Wnt responsive status. In either case, L-WNT3A appears to be sufficient to induce bone formation either from fibrous tissue (Yin et al. 2016) or from the fibrous PDL (Fig. 4) for purposes of accelerated implant osseointegration.
Conclusion
Wnt-responsive cells contribute to new bone formation around immediate postextraction implants, and this Wnt-responsive population initially resides in the PDL. Therefore, if clinical observations support it, PDL remnants should be preserved in extraction sockets. Delivery of L-WNT3A to the extraction socket at the time of implant placement significantly accelerates osseointegration, which provides another opportunity protecting existing osteocytes from death and simultaneously stimulating osteoprogenitor cells to give rise to new bone in the immediate postextraction implant environment.
Author Contributions
X. Yuan, X. Pei, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; Y. Zhao, Z. Li, C.H. Chen, U.S. Tulu, B. Liu, L.A. Van Brunt, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; J.B. Brunski, contributed to conception, design, data analysis, and interpretation, critically revised the manuscript; J.A. Helms, contributed to conception, design, data analysis, and interpretation, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
DS_10.1177_0022034518765757 – Supplemental material for Biomechanics of Immediate Postextraction Implant Osseointegration
Supplemental material, DS_10.1177_0022034518765757 for Biomechanics of Immediate Postextraction Implant Osseointegration by X. Yuan, X. Pei, Y. Zhao, Z. Li, C.H. Chen, U.S. Tulu, B. Liu, L.A. Van Brunt, J.B. Brunski and J.A. Helms in Journal of Dental Research
Footnotes
Acknowledgements
We thank Liao Wang and Yan Wu for their invaluable suggestions on establishing the mouse model.
This research project was supported by a grant from the National Institutes of Health (R01DE024000-11; J.A.H. and J.B.B.), the National Natural Science Foundation of China (81601613, X.P.), and the Science and Technology Support Program of Sichuan Province (2016FZ0085; X.P.).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.