
Abstract
Sirt1 promotes odontoblastic gene expression in human dental pulp cells, whereas the inhibition of Sirt1 downregulates the expression of those genes. To investigate whether the overexpression of Sirt1 in mesenchymal stem cells (MSCs) driven by Prx1 promoter could rescue the dentin formation defects in Bmi1-deficient (Bmi1−/−) mice, we established the MSCs overexpressing Sirt1 in Bmi1 knockout mice (Sirt1TGBmi1−/−). First, we used Prx1-Cre/ROSAnTnG mice to demonstrate that Prx1 linage cells exist mainly in the pulp horns at 4 wk of age. Second, we found that 4-wk-old Sirt1TG mice had increased tooth volume as compared with wild-type (WT) littermates. The expression level of Sirt1 was significantly higher in dental papilla mesenchymal cells of Sirt1TG mice than WT mice. Furthermore, we demonstrated that the tooth mineralization, dental volume, dentin sialoprotein–immunopositive areas, odontoblastic gene expression, and percentage of proliferating BrdU-positive cells were significantly elevated in the Sirt1TG mice and dramatically reduced in the Bmi1–/– mice versus the WT littermates at 4 wk of age. However, the areas of predentin and the percentage of TUNEL-positive apoptotic cells were significantly reduced in the Sirt1TG mice but dramatically increased in the Bmi1−/− mice as compared with the WT littermates. All these parameters were rescued in the Sirt1TGBmi1−/− mice versus the Bmi1−/− mice. Finally, by using dental papilla mesenchymal cells, we found that the overexpression of Sirt1 rescued the reduced cell proliferation and differentiation and increased the cell apoptosis caused by Bmi1 deficiency, which was associated with increased p53 deacetylation. Therefore, this study indicates that Sirt1 is a potential therapeutic target for promoting dentin formation in an anabolic approach to the treatment of dental developmental defects.
Keywords
Introduction
Sirtuin 1 (Sirt1), a member of the sirtuin family, is an NAD+-dependent protein deacetylase that plays important roles in modulating gene transcription, energy metabolism, cell proliferation, and apoptosis (Houtkooper et al. 2012; Nogueiras et al. 2012). In addition to histone deacetylation, Sirt1 deacetylates a number of protein substrates, including p53, FOXO3a, PPARγ, and NF-κB (Nogueiras et al. 2012), to perform its biological functions. Sirt1 heterozygous mice (Sirt1+/−) show a significantly decreased volume of long bones and vertebrae as compared with that in wild-type (WT) mice at 12 wk of age (Cohen-Kfir et al. 2011). Osteoblast-specific Sirt1 deletion achieved by crossing Sirt1-flox/flox mice with ColI-Cre transgenic mice resulted in decreased bone mass caused by reduced bone formation (Edwards et al. 2013). In vitro studies showed that increased Sirt1 promotes the expression of odontoblastic genes in human dental pulp cells, including genes encoding alkaline phosphatase (ALP), osteocalcin (OCN), dentin matrix protein 1 (DMP1), and dentin sialophosphoprotein (DSPP), whereas the inhibition of Sirt1 downregulates the expression of those genes (Kim et al. 2012; Jang et al. 2015). However, no in vivo study has demonstrated the function of Sirt1 in dentin formation.
B-lymphoma Mo-MLV insertion region 1 (Bmi1) was initially identified as an oncogene that regulates cell proliferation and transformation (Park et al. 2004). Bmi1 plays an important but complex role in the self-renewal and differentiation of mesenchymal stem cells (MSCs; Molofsky et al. 2005; Zhang et al. 2010; Oguro 2011). Both Bmi1 and Sirt1 belong to the polycomb group proteins and have common downstream target genes, including p16, p19, and p53 (Martinez and Cavalli 2006). Using Bmi1 gene–deficient (Bmi1−/−) mice, we previously reported that Bmi1 plays an important role in dentin formation by promoting cell proliferation and maintaining the redox balance (Yin et al. 2016). However, we only partly rescued the defects in dentin formation in the Bmi1−/− mice by supplying the antioxidant N-acetylcysteine to their drinking water for 2 wk from 3 wk of age. We inferred that at 3 wk old, Bmi1−/− mice already displayed severe defects in dentin formation, and it was difficult to correct these phenotypes after they had been established. We also found that Bmi1 deficiency caused significant downregulation of Sirt1 protein; resveratrol, a Sirt1 activator, markedly promoted osteoblast differentiation in bone marrow MSCs from Bmi1−/− mice (Zhang et al. 2010). Therefore, we attempted to rescue dentin formation defects in Bmi1−/− mice with genetic methods, such as overexpressing the Sirt1 gene in Bmi1−/− mice by crossing the Bmi1−/− mice with the Sirt1-transgenic mice.
Paired-related homeobox gene 1 (Prx1) is a marker of MSCs, which are widely used for mesenchymal lineage–specific knockdown or overexpression of target genes (Elefteriou and Yang 2011). However, there is a lack of clear evidence that Prx1 transcripts are expressed in dental pulp cells. We therefore used Prx1-Cre/ROSAnTnG mice to trace Prx1 linage cells in postnatal dental tissue. Furthermore, we established the MSCs overexpressing Sirt1 in Bmi1 knockout mice (Sirt1TGBmi1−/−) and compared their dentin formation phenotypes with their WT, Sirt1TG, and Bmi1−/− littermates.
Materials and Methods
Animals
Prx1-Cre/ROSAnTnG mice were obtained by crossing Prx1-Cre male mice with ROSAnTnG female mice. Sirt1TG mice were originally generated in our laboratory (Yan et al. 2015). Briefly, pECE-Sirt1 plasmid corresponding to Sirt1 transcript and PNCHS4-Prx1-cre-NLS plasmid containing Prx1 promoter transcript were amplified, digested with NotI/SacI enzymes, and ligated with each other; then, the plasmid was microinjected into the pronucleus of a fertilized egg. The injected embryos were subsequently transferred into the oviducts of pseudopregnant surrogate mothers. Founder mice were born harboring the injected foreign gene in their genomes. This line of Sirt1TG mice overexpresses Sirt1 in MSCs, driven by the Prx1 promoter. Bmi1−/− mice generated in a C57BL/6J background were obtained from Dr. Anton Berns (van der Lugt et al. 1994). Sirt1TG mice and Bmi1+/− mice were crossed to generate Sirt1TGBmi1+/− double mutant mice, and Sirt1TGBmi1+/− mice and Bmi1+/− mice were crossed to generate WT, Sirt1TG, Bmi1−/−, and Sirt1TGBmi1−/− mice. All animal procedures were conducted in accordance with approved guidelines of the Committee of Nanjing Medical University for Animal Resources (approval ID1601253).
Histology
Three-day-old or 4-wk-old mice were used, as specified in the figures. Mice were given 2 injections of 5-bromo-2′-deoxyuridine (BrdU; 1 mg/mouse/injection, intraperitoneal) spaced 16 h apart and were euthanized 2 h after the second injection. The mouse mandibles were removed, fixed in 4% paraformaldehyde, decalcified in 14% EDTA, and embedded in Tissue-Tek for frozen sections (Sun et al. 2017) or embedded in paraffin for paraffin sections (Sun et al. 2016). Frozen sections were directly used for fluorescence microscopy or subjected to BrdU and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. Paraffin sections were stained with hematoxylin and eosin (H&E), analyzed histochemically for total collagen with Sirius red, or analyzed immunohistochemically as described in the online Appendix.
Cell Culture
Dental papilla mesenchymal cells (DPMCs) were isolated from the first molars of 3-d-old mutant and WT mice with tying forceps under microscopy and digested with 3 mg/mL of collagenase type I (Sigma-Aldrich) and 4 mg/mL of dispase (Sigma-Aldrich) for 60 min at 37 °C (Wang et al. 2013). The cells were grown to confluence in α-minimal essential medium containing 10% fetal bovine serum (ScienCell Research Laboratories). Third-passage cells were used for cell characterization (Appendix Fig. 1), confirmation of the Prx1 expression, and other experiments.
Statistical Analysis
Results are given as means ± SEM. Statistical analysis was performed with GraphPad Prism 5 software (GraphPad Software Inc.). Comparisons between 2 groups were made with a 2-tailed unpaired Student’s t test. Comparisons of 4 groups were made with 1-way analysis of variance, followed by Dunnett’s post hoc multiple-comparison test. P < 0.05 was considered statistically significant.
Additional information may be found in the online Appendix.
Results
Genetic Tracing of Postnatal Prx1+ Dental Stem Cells
We first used Prx1-Cre/ROSAnTnG mice to trace Prx1 linage cells in the postnatal dental tissue. In Prx1-Cre/ROSAnTnG mice, all Prx1-Cre+ cells and their descendants in dental tissue express GFP, whereas all Prx1-Cre– cells and their descendants express tdTomato. We found many Prx1-Cre+ cells located in the pulp horns at 3 days or 4 weeks after birth (Fig. 1A, C). There were about 10% dental pulp cells labeled with GFP in the region of pulp horns on postnatal day 3 (Fig. 1A), whereas there was about 20% GFP+ cells on postnatal week 4 (Fig. 1C). Surprisingly, the remaining regions of dental pulp, especially apical papilla, had a lack of expression of Prx1 (Fig. 1B, D) on postnatal day 3 and postnatal week 4 (Fig. 1B, D). In Figure 1B and D, only some of Prx1-Cre+ cells were located on the surface of the trabecular bone.
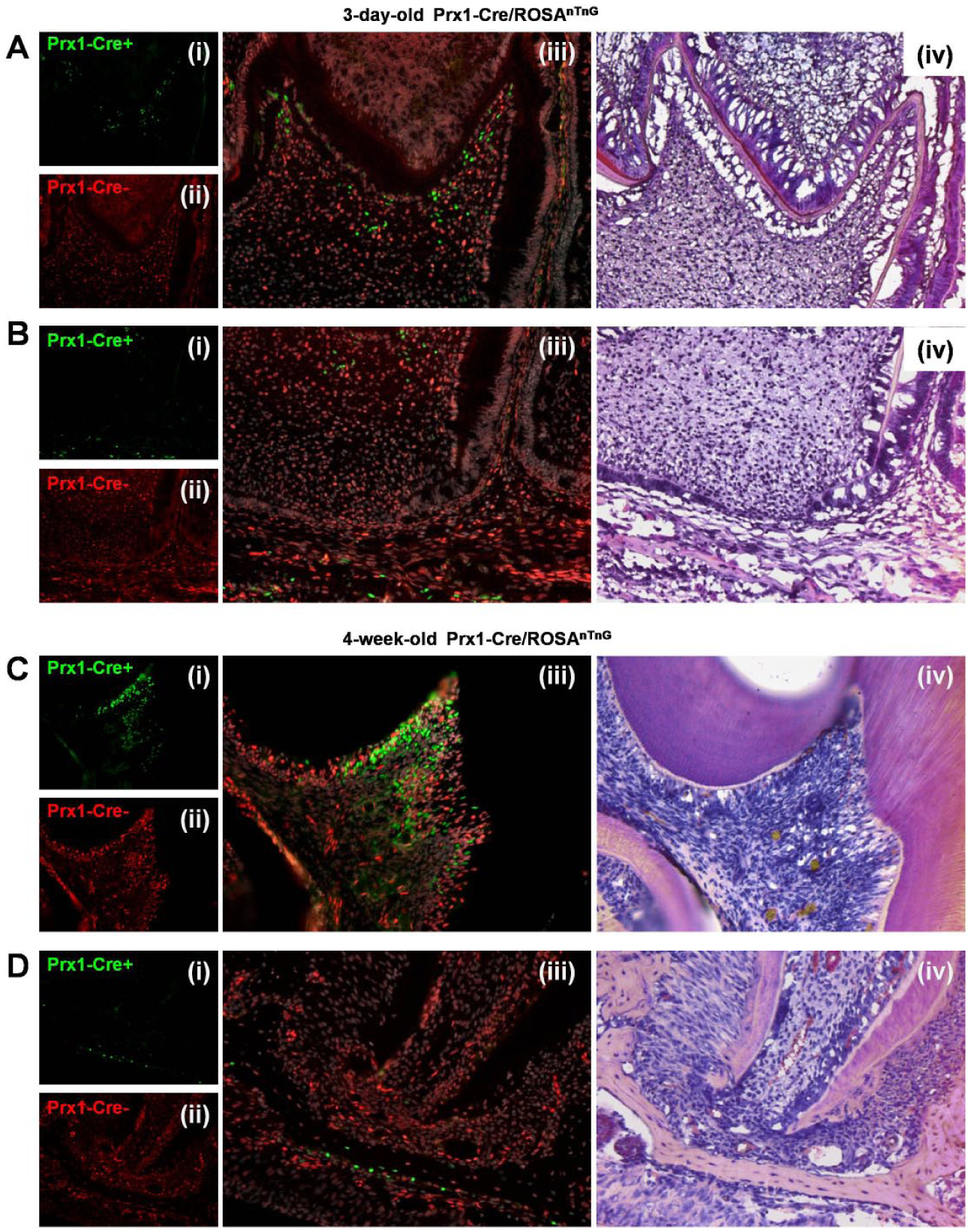
Genetic tracing of postnatal Prx1+ dental stem cells. Frozen sections of first molar from (
Effect of Overexpression of Sirt1 in MSCs on Tooth Mineralization and Tooth Volume in Bmi1−/− Mice
Since Prx1 cells exist in dental tissue postnatally, we used Sirt1TG mice with overexpressed Sirt1 in MSCs, which were driven by the Prx1 promoter, and their WT littermates to compare their dental phenotypes. H&E staining and histomorphometric analysis showed that the dental volume of the incisors and the first molars were increased in the Sirt1TG mice versus their WT littermates (Fig. 2A). We next cultured DPMCs from WT and Sirt1TG mice, which are the mesenchymal lineage cells. We found that the Prx1 protein expression level was equal between the DPMCs from WT and Sirt1TG mice, while the Sirt1 protein expression level was increased significantly in Sirt1TG DPMCs as compared with DPMCs from the WT littermates (Fig. 2B). Protein from spleen cells (not mesenchyme derived) was included as negative control for Prx1 and Sirt1 expression (Fig. 2B).
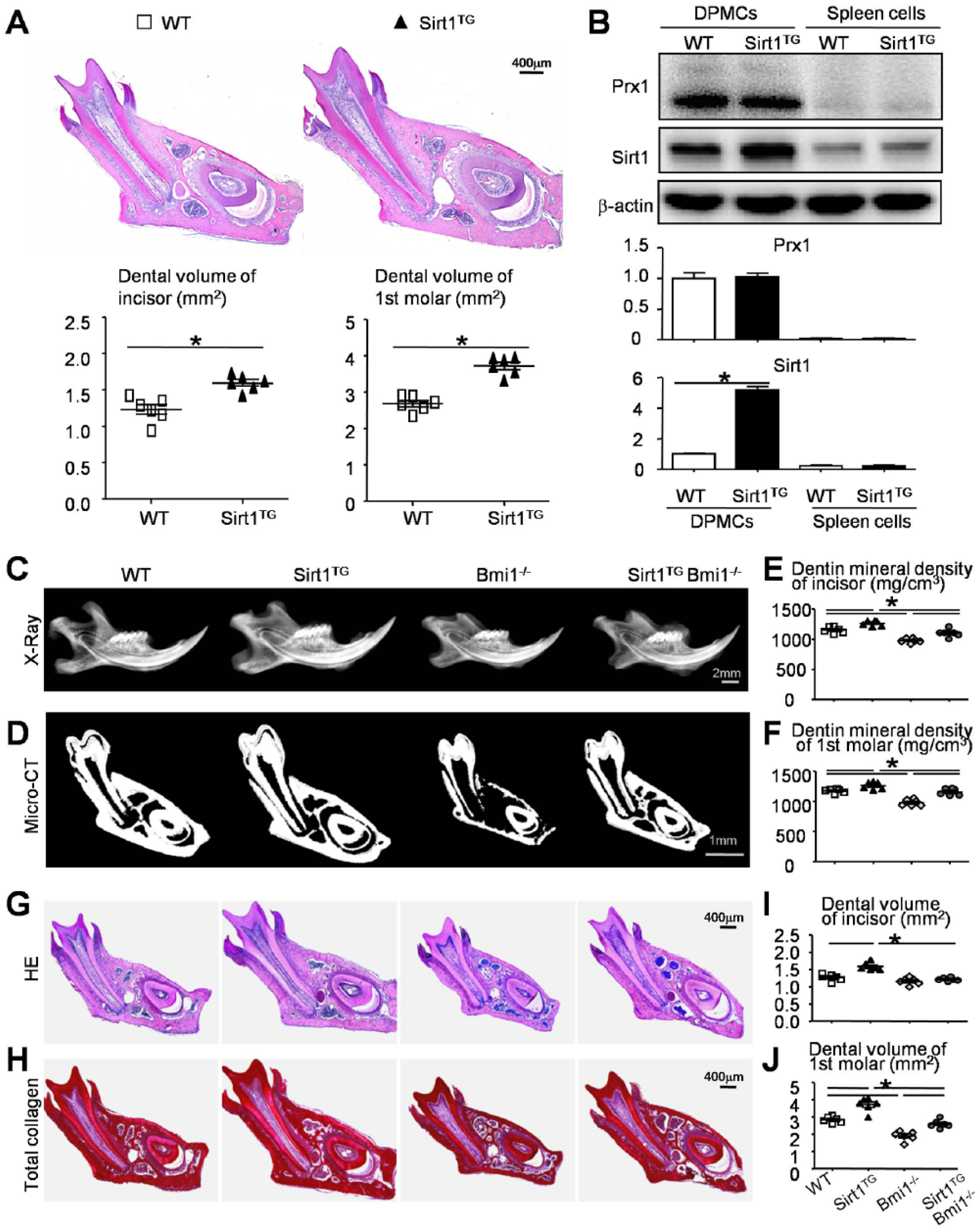
Effect of overexpression of Sirt1 in mesenchymal stem cells on tooth mineralization and tooth volume in Bmi1−/− mice. Four-week-old wild-type (WT), Sirt1TG, Bmi1−/−, and Sirt1TGBmi1−/− mice were used. (
Given the promotional effect of overexpressed Sirt1 on tooth volume, we hypothesized that overexpression of Sirt1 in MSCs may rescue or partly rescue dentin formation defects caused by Bmi1 deficiency (Yin et al. 2016). Radiography and micro–computed tomography scanning were performed to determine the tooth mineralization between WT and mutant mice. When compared with their WT littermates, the mineralized dentin density of the incisors and molars were increased in the Sirt1TG mice, reduced in the Bmi1−/− mice, but not significantly altered in the Sirt1TGBmi1−/− mice (Fig. 2C–F). Both parameters were significantly greater in Sirt1TGBmi1−/− mice than in Bmi1−/− mice (Fig. 2C–F).
H&E staining, histochemically for total collagen, and a histomorphometric analysis showed that the dental volume of incisors was increased in the Sirt1TG mice (Fig. 2G–I). However, the dental volume of incisors had no significant differences between the Bmi1−/− and Sirt1TGBmi1−/− mice in sections through the first molars and incisors (Fig. 2G–I). Notably, the dental volume of the first molars was increased in the Sirt1TG mice but dramatically reduced in the Bmi1−/− mice as opposed to their WT littermates. However, the dental volume of the first molars was increased in the Sirt1TGBmi1−/− mice versus their Bmi1−/− littermates, and it was rescued to near levels as their WT littermates (Fig. 2G, H, J).
Effect of Sirt1 Overexpression in MSCs on Predentin Maturation and Dentin Formation in Bmi1−/− Mice
We then determined the alterations of predentin maturation and dentin formation in 4 genotype mice. The thickness of predentin in the first molars and the immunoreactivity for DSP were assessed with histology and immunohistochemistry, respectively. The ratio of the areas of predentin to dentin was reduced in the Sirt1TG mice but increased in the Bmi1−/− mice versus their WT littermates (Fig. 3A, C). The DSP immunoreactivity was increased in the Sirt1TG mice but decreased in the Bmi1−/− mice when compared with their WT littermates (Fig. 3B, D). Both parameters were improved in the Sirt1TGBmi1−/− mice versus their Bmi1−/− littermates (Fig. 3A–D).
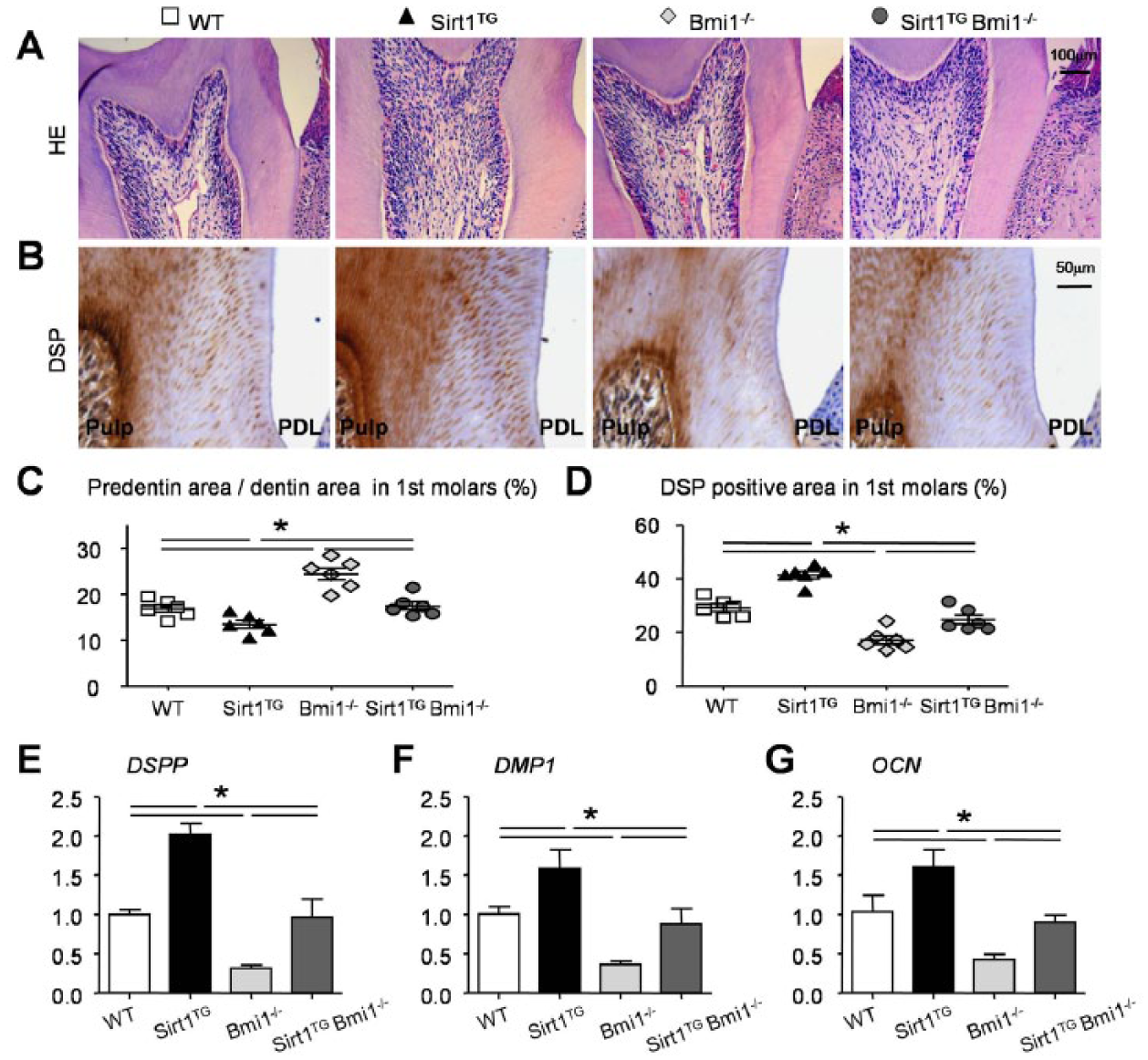
Effects of overexpression of Sirt1 in mesenchymal stem cells on predentin maturation and dentin formation in Bmi1−/− mice. Four-week-old wild-type (WT), Sirt1TG, Bmi1−/−, and Sirt1TGBmi1−/− mice were used. Paraffin sections through the root wall of the first molars were (
We also examined the expression of genes involved in dentin formation. RNA was isolated from the molars, and real-time reverse transcription polymerase chain reaction was performed. The expression of genes, including DSPP, DMP1, and OCN, was significantly upregulated in the Sirt1TG mice but reduced in the Bmi1−/− mice as compared with their WT littermates. However, they were increased in the Sirt1TGBmi1−/− mice versus Bmi1−/− mice (Fig. 3E–G).
Effect of Sirt1 Overexpression in MSCs on Cell Proliferation and Apoptosis in Bmi1−/− Mice
We next used the RNA isolated from molars to confirm Sirt1 gene expression and the expression of genes involved in cell proliferation and apoptosis. The Sirt1 and PCNA mRNA levels were significantly upregulated in the Sirt1TG mice but reduced in the Bmi1−/− mice as compared with their WT littermates. However, they were increased in the Sirt1TGBmi1−/− mice as opposed to the levels in the Bmi1−/− mice (Fig. 4A, B). Caspase3 gene expression was reduced in the Sirt1TG mice but increased in the Bmi1−/− mice versus their WT littermates. However, it was significantly reduced in the Sirt1TGBmi1−/− mice against that in the Bmi1−/− mice (Fig. 4C).
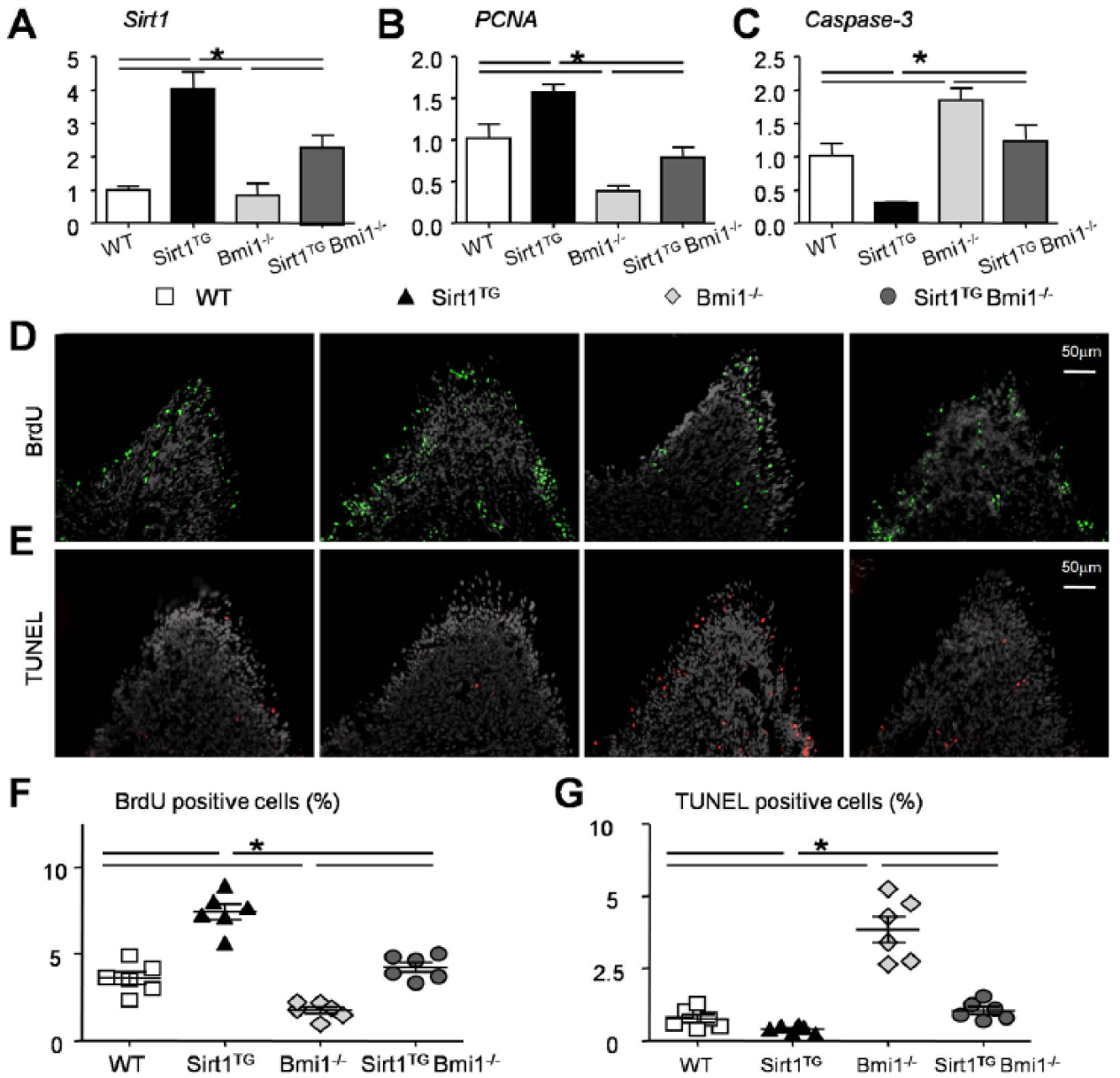
Effects of overexpression of Sirt1 in mesenchymal stem cells on cell proliferation and apoptosis in Bmi1−/− mice. Four-week-old wild-type (WT), Sirt1TG, Bmi1−/−, and Sirt1TGBmi1−/− mice were used. Comparison of (
To confirm the data, the proliferation and apoptosis of cells were examined further by BrdU and TUNEL assays, respectively. BrdU- or TUNEL-positive cells were mainly determined to be in the pulp horns, where Prx1 cells were located the most (Fig. 1C). BrdU-positive cells were increased in the Sirt1TG mice, reduced in the Bmi1−/− mice, but not significantly altered in the Sirt1TGBmi1−/− mice relative to their WT littermates (Fig. 4D, F). The percentage of apoptotic cells in the pulp horns was reduced in the Sirt1TG mice and increased in the Bmi1−/− mice but not significantly altered in the Sirt1TGBmi1−/− mice relative to their WT littermates (Fig. 4E, G). Both parameters were improved in the Sirt1TGBmi1−/− mice as compared with their Bmi1−/− littermates (Fig. 4D–G).
Effect of Sirt1 Overexpression in MSCs on p53 Deacetylation, Cell Proliferation, Apoptosis, and Differentiation in Bmi1−/− Mice
According to the literature, Sirt1 binds and deacetylates p53, thus regulating cell proliferation and apoptosis (Kume et al. 2006; Hori et al. 2013; Lee and Gu 2013). Acetylation is required for p53 function (Appella and Anderson 2001). To investigate the underlying mechanism, we performed a Western blot analysis of extracts of the molars from the mutant and WT mice. Bmi1 expression was undetectable in the Bmi1−/− and Sirt1TGBmi1−/− mice, as expected (Fig. 5A). The expression of Acetyl-p53 (K370), Acetyl-p53 (K379), BAX, and p21 was reduced in the Sirt1TG mice and increased in the Bmi1−/− mice but not significantly altered in the Sirt1TGBmi1−/− mice relative to the levels in their WT littermates (Fig. 5A). In contrast, the expression of Sirt1 and Bcl-2 (B-cell lymphoma 2) was increased in the Sirt1TG mice and reduced in the Bmi1−/− mice but not significantly altered in the Sirt1TGBmi1−/− mice relative to the expression levels in WT littermates (Fig. 5A). All the parameters were improved in the Sirt1TGBmi1−/− mice when compared with their Bmi1−/− littermates (Fig. 5A). To confirm these data, a Sirt1 activator, resveratrol, was used to treat DPMCs from WT and Bmi1−/− mice. Data revealed that in Bmi1−/− DPMCs, Sirt1 activation decreased the acetylation of p53, rescued Bax and p21 expression levels, but increased Sirt1 and Bcl-2 expression levels relative to the DPMCs of WT littermates (Fig. 5B).
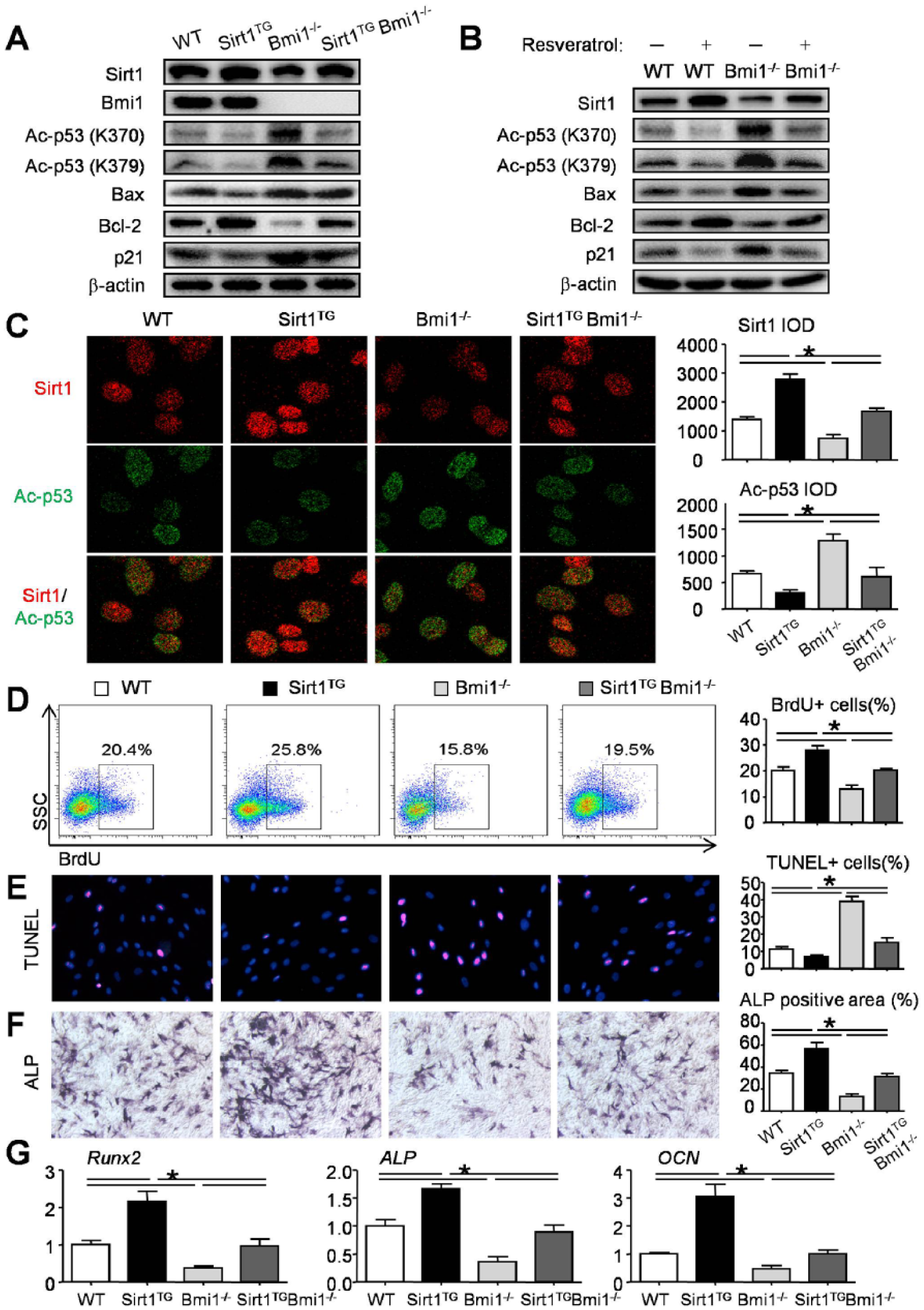
Effect of Sirt1 overexpression in mesenchymal stem cells on p53 deacetylation, cell proliferation, apoptosis, and differentiation in Bmi1−/− mice. (
Furthermore, the immunofluorescence assay was performed with DPMCs from WT and mutant mice with anti-Sirt1 and anti-Ac-p53 (K370) antibodies. The results showed that Sirt1 expression level was increased in the Sirt1TG DPMCs and reduced in the Bmi1−/− DPMCs but not significantly altered in the Sirt1TGBmi1−/− DPMCs relative to its expression in the DPMCs from the WT littermates (Fig. 5C). In contrast, Ac-p53 expression was reduced in the Sirt1TG DPMCs and increased in the Bmi1−/− DPMCs but not significantly altered in the Sirt1TGBmi1−/− DPMCs as compared with its expression in the DPMCs from the WT littermates (Fig. 5C).
To investigate whether the overexpression of Sirt1 affects cell proliferation and apoptosis, we examined DPMCs from 4 genotype mice with BrdU incorporation, flow-cytometric analysis, and a TUNEL assay. The percentage of BrdU-positive cells was increased in the DPMCs from the Sirt1TG mice and reduced in the Bmi1−/− DPMCs but not significantly altered in the Sirt1TGBmi1−/− DPMCs relative to its percentage from the WT littermates (Fig. 5D). In contrast, the percentage of TUNEL-positive cells was reduced in the DPMCs from the Sirt1TG mice and increased in the Bmi1−/− DPMCs but not significantly altered in the Sirt1TGBmi1−/− DPMCs as compared with its percentage from the WT littermates (Fig. 5E). We also examined cell differentiation using DPMCs from 4 genotype mice with osteoblast induction medium. The percentage of ALP-positive cells was increased in the Sirt1TG mice and reduced in the Bmi1−/− mice but not significantly altered in the Sirt1TG Bmi1−/− mice relative to its percentage from the WT littermates (Fig. 5F). These data were further confirmed by expressions of the osteoblast marker genes Runx2, ALP, and OCN (Fig. 5G). All the parameters were improved in the Sirt1TGBmi1−/− mice as compared with the Bmi1−/− littermates (Fig. 5D–G).
Discussion
Prx1 encodes a homeobox transcription factor that is mesenchymally expressed in tissues that develop into cartilage (Blaney Davidson et al. 2014), bone (ten Berge et al. 1998), or tooth structures (Mitchell et al. 2006). The Prx1 promoter is widely used to study mesenchymal cells and to generate mesenchymal lineage cells for specific knockout or overexpression mouse models (Elefteriou and Yang 2011). However, there is lack of clear evidence that Prx1 transcripts are expressed in postnatal dental tissue. In this study, by using Prx1-Cre/ROSAnTnG mice, we demonstrated for the first time that there are many Prx1+ cells within dental pulp, mainly located in pulp horns, whether on postnatal day 3 or postnatal week 4. Blood vessels and nerves are present in the central pulp region. In the peripheral region, there are 3 zones: odontoblastic zone, cell-free zone, and cell-rich zone. The cell-rich zone contains fibroblasts and undifferentiated mesenchymal cells. It looks like Prx1 cells mainly locate in the cell-rich zone of pulp horns. We speculate that these Prx1 cells facilitate the recruitment of newly differentiating cells to form odontoblasts when dentin is grinded. Furthermore, in the apical papilla, which harbors MSCs (Sohn et al. 2014; Sonoyama et al. 2008), there was a lack of expression of Prx1. We speculate that Prx1 is not a good marker for apical papilla cells. More experiments are needed to delineate the precise cell marker to define stem cells within apical papilla.
Bmi1 is required for the self-renewal of stem cells, including hematopoietic stem cells (Iwama et al. 2004; Park et al. 2003), neural stem cells (Molofsky et al. 2005), lung stem cells (Zacharek et al. 2011), and intestinal stem cells (López-Arribillaga et al. 2015). We previously reported that Bmi1 maintains the self-renewal of bone marrow MSCs by inhibiting the expression of p27, p16, and p19 and alters the fate of bone marrow MSCs by enhancing osteoblast differentiation and inhibiting adipocyte differentiation, at least in part by stimulating Sirt1 expression (Zhang et al. 2010). We then reported that Bmi1 deficiency leads to dentin formation defects (Yin et al. 2016). Therefore, in this study, we used a genetic method to overexpress Sirt1 in the MSCs of Bmi1−/− mice to determine whether these dentin formation defects could be rescued. Our data suggested that the dental volume, odontoblastic gene expression levels, and percentage of proliferating BrdU-positive cells were significantly increased, whereas the percentage of TUNEL-positive apoptotic cells was significantly reduced in the Sirt1TGBmi1−/− mice when compared with the Bmi1−/− mice. All these parameters were almost completely corrected in the Sirt1TGBmi1−/− mice versus the Bmi1−/− mice.
p53 is a protein that regulates the cell cycle and has been shown to participate in cell proliferation and cell apoptosis (Alenzi 2004). Acetylation is required for p53 function (Appella and Anderson 2001). Our data showed that Sirt1 deacetylated the p53 protein. It has been reported that Sirt1 deacetylates p53 and attenuates its ability to transactivate its downstream target genes, such as p21 for cell-cycle arrest and Bax for apoptosis (Luo et al. 2001; Vaziri et al. 2001). We also observed that p21 and Bax were reduced in the Sirt1TG mice and increased in the Bmi1−/− mice but were not significantly altered in the Sirt1TGBmi1−/− mice relative to the levels in WT littermates. Bmi1 deficiency leads to increased p53 accumulation on its target gene promoters (Chatoo et al. 2009). We also observed increased acetylated p53 expression in the dental extracts and DPMCs of the Bmi1−/− mice. Furthermore, the overexpression of Sirt1 rescued the abnormal cell apoptosis and proliferation of the Bmi1−/− mice by inhibiting acetylated p53 expression. Consequently, our results indicate that overexpression of Sirt1 in MSCs stimulates dentin formation, at least partially through p53 deacetylation.
In summary, using a transgenic mouse model overexpressing Sirt1 in its MSCs, we demonstrated that Sirt1 overexpression increased dental volume, promoted predentin maturation and dentin formation, rescued tooth mineralization and dentin maturation defects caused by Bmi1 deficiency. Using DPMCs, we found that the overexpression of Sirt1 in MSCs promoted p53 deacetylation and cell proliferation and differentiation and inhibited cell apoptosis in WT and Bmi1−/− mice. Thus, this study demonstrates that Sirt1 is a potential target for promoting dentin formation in an anabolic approach to dental developmental defects.
Author Contributions
H. Wang, D. Miao, W. Sun, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; C. Lv, contributed to conception, design, data acquisition, and interpretation, critically revised the manuscript; Y. Gu, contributed to conception, data acquisition, and interpretation, critically revised the manuscript; Q. Li, contributed to conception, data analysis, and interpretation, critically revised the manuscript; L. Xie, contributed to design, data analysis, and interpretation, critically revised the manuscript; H. Zhang, contributed to conception, design, and data interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
DS_10.1177_0022034518781509 – Supplemental material for Overexpressed Sirt1 in MSCs Promotes Dentin Formation in Bmi1-Deficient Mice
Supplemental material, DS_10.1177_0022034518781509 for Overexpressed Sirt1 in MSCs Promotes Dentin Formation in Bmi1-Deficient Mice by H. Wang, C. Lv, Y. Gu, Q. Li, L. Xie, H. Zhang, D. Miao and W. Sun in Journal of Dental Research
Footnotes
A supplemental appendix to this article is available online.
This research was supported by grants from the National Natural Science Foundation of China (81670965 to W.S., 81500815 to H.W., and 81230009 to D.M.), the Natural Science Foundation of Jiangsu Province in China (BK20150999 to H.W.), the Natural Science Fund for Colleges and Universities of Jiangsu Province in China (15KJB320004 to H.W.), and the Open Project from Jiangsu Key Laboratory of Oral Diseases, Nanjing Medical University (JSKLOD-KF-1707 to C.L., JSKLOD-KF-1706 to Q.L.).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.