
Abstract
Oral cancer has a high annual incidence rate all over the world, and the tongue is the most frequently affected anatomic structure. The current standard care is ablative surgery of malignant neoplasm, followed by tongue reconstruction with free flap. However, such reconstructive modalities with postsurgery radiotherapy or chemotherapy can hardly support the functional recovery of the tongue—particularly, functional taste bud regeneration—in reconstructed areas, thus seriously affecting patients’ prognosis and life quality. Using a critical-sized tongue defect model in rats, we show that combinatory transplantation of small intestinal submucosa–extracellular matrix (SIS-ECM) with gingival mesenchymal stem cells (GMSCs) or their derivative exosomes promoted tongue lingual papillae recovery and taste bud regeneration as evidenced by increased expression of CK14, CK8, and markers for type I, II, and III taste bud cells (NTPdase 2, PLC-β2, and AADC, respectively). In addition, our results indicate that GMSCs or their derivative exosomes could increase BDNF expression, a growth factor that plays an important role in the proliferation and differentiation of epithelial basal progenitor cells into taste bud cells. Meanwhile, we showed an elevated expression level of Shh—which is essential for development, homeostasis, and maintenance of the taste bud organ—in wounded areas of the tongue among animals treated with GMSC/SIS-ECM or exosome/SIS-ECM as compared with SIS-ECM control. Moreover, our data show that GMSCs or their derivative exosomes promoted innervation of regenerated taste buds, as evidenced by elevated expressions of neurofilament and P2X3 at the injury areas. Together, our findings indicate that GMSC/SIS-ECM and exosome/SIS-ECM constructs can facilitate taste bud regeneration and reinnervation with promising potential application in postsurgery tongue reconstruction of patients with tongue cancer.
Introduction
There are >30,000 new cases of oral cancer diagnosed every year across the world, while the tongue is the most frequently affected anatomic structure, especially the anterior two-thirds of the dorsal tongue (Kim et al. 2012). Tongue cancer is usually first diagnosed at advanced stages, when it has already involved a large volume of tongue tissue despite the availability of many advanced diagnostic technologies, including computed tomography and magnetic resonance imaging (Kim et al. 2012). The current standard care for these patients is glossectomy, which requires surgical removal of malignant neoplasm (Shanti and O’Malley 2018). Resection of a large volume of tongue tissue leads to anatomic deformity, functional debilitation, as well as psychological impact. Tongue reconstruction is highly demanded after glossectomy to ensure patients’ prognosis and postsurgery life quality.
Through decades of development in tissue reconstruction, microvascular-free tissue transfer has evolved as the reconstructive technique of choice following surgical removal of malignant neoplasms (Engel et al. 2010). However, those skin tissue–derived tongue reconstructive modalities with postsurgery radiotherapy or chemotherapy can hardly support any functional taste bud regeneration in reconstructed areas and can even cause taste bud loss in normal tongue surfaces, which may seriously affect patients’ taste sensation and subsequently tamp down their appetite (Feller et al. 2010; Mannelli et al. 2018). Therefore, securing patients’ taste sensory function and meeting their nutrition requirements after tongue cancer therapy are essential for patients’ recovery, physically and psychologically (Jensen and Peterson 2014).
Exosomes, with a size range of 30 to 150 nm, are formed by the fusion of multivesicular bodies within cell cytoplasm and plasma membrane and are released into the extracellular milieu (Katsuda et al. 2013). Accumulating evidence has shown that mesenchymal stem cell (MSC)–derived exosomes exhibit therapeutic effects in a variety of disease models (Furuta et al. 2016) comparable to the capacity of MSCs (Lankford et al. 2018). Therefore, MSC-derived exosomes may represent a novel cell-free therapeutic paradigm for tissue regeneration and treatment of a variety of diseases, bypassing reported drawbacks associated with the direct use of MSCs (Lou et al. 2017).
We have successfully established a critical-sized tongue defect model in rats and have demonstrated that the transplantation of small intestinal submucosa–extracellular matrix (SIS-ECM) constructs seeded with gingival MSCs (GMSCs) could promote muscle regeneration and reduce fibrosis during tongue reconstruction (Xu et al. 2017). Considering the multilayered structure of ablative tongue tissue as well as its specialized taste sensory function, we herein explored whether GMSCs and their derivative exosomes laden with SIS-ECM matrix could facilitate taste bud regeneration during tongue reconstruction.
Materials and Methods
Extensive experimental details are included in the Appendix.
Animals
Six- to 7-wk-old Sprague-Dawley female rats weighing 200 to 250 g (Charles River Laboratories) were used in this study. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania. A prospective randomized controlled animal model design was adopted according to all the recommendations of the ARRIVE guidelines (Animal Research: Reporting In Vivo Experiments). Rats were group housed in polycarbonate cages (2 animals per cage) with controlled temperature (18 to 23 °C ), 40% to 65% of humidity, and a 12-h light/dark cycle. Rats were acclimatized for 1 wk before the experiment, fed with a standard laboratory diet, and allowed ad libitum access to drinking water.
Cell Cultures and Exosome Purification
According to the approved Institutional Review Board protocol at the University of Pennsylvania, gingival tissues were obtained as remnants of discarded tissues from healthy human subjects who underwent routine dental procedures. Detailed protocols for GMSC culture (Zhang et al. 2009) and exosome purification are available in the Appendix.
GMSC/SIS-ECM Construct Preparation and Animal Surgery
Strips (6 × 6 mm) of decellularized porcine SIS-ECM 2.0 (Cook Biotech, Inc.) was prepared as described previously (Xu et al. 2017). For detailed procedures of animal surgery and transplantation of SIS-ECM with GMSCs or exosomes, see the Appendix.
Histologic and Immunofluorescence Studies
The tongue samples were fixed in 10% neutralized formalin for 48 h, and 5-µm-thick paraffin sections and 8-µm-thick cryosections were cut. Detailed procedures for hematoxylin and eosin and immunofluorescence staining can be found in the Appendix.
Statistical Analysis
Differences between the experimental and control groups were analyzed by the paired Student’s t test. One-way analysis of variance was used to test the statistical significance of multiple group differences, unless otherwise indicated. P < 0.05 was considered statistically significant. SPSS software (version 18.0; IBM) was used for all the analyses.
Results
Purification of GMSC-Derived Exosomes
Initially, we showed that GMSCs harbored abundant CD63 signals in the cytoplasmic compartment (Fig. 1A), a popularly used marker for identification of exosomes (Khan and Kishore 2017; Phinney and Pittenger 2017). By using centrifuge filtration and ExoQuick-TC Exosome Precipitation—a well-established method for purifying exosomes from different types of cells (Khan and Kishore 2017; Phinney and Pittenger 2017; Pu et al. 2017)—we purified and isolated a fraction of extracellular vesicles from the culture supernatants of GMSCs. NanoSight analysis showed that the GMSC-derived extracellular vesicle fraction exhibited a mean ± SD size of 95.8 ± 5.8 nm (Fig. 1B), a nanoparticle-size range that is in agreement with exosome size distribution as previously reported (Yue et al. 2017). Western blotting confirmed that the purified exosomes from different strains of GMSCs expressed exosomal surface markers CD63 and CD9 (Fig. 1C).
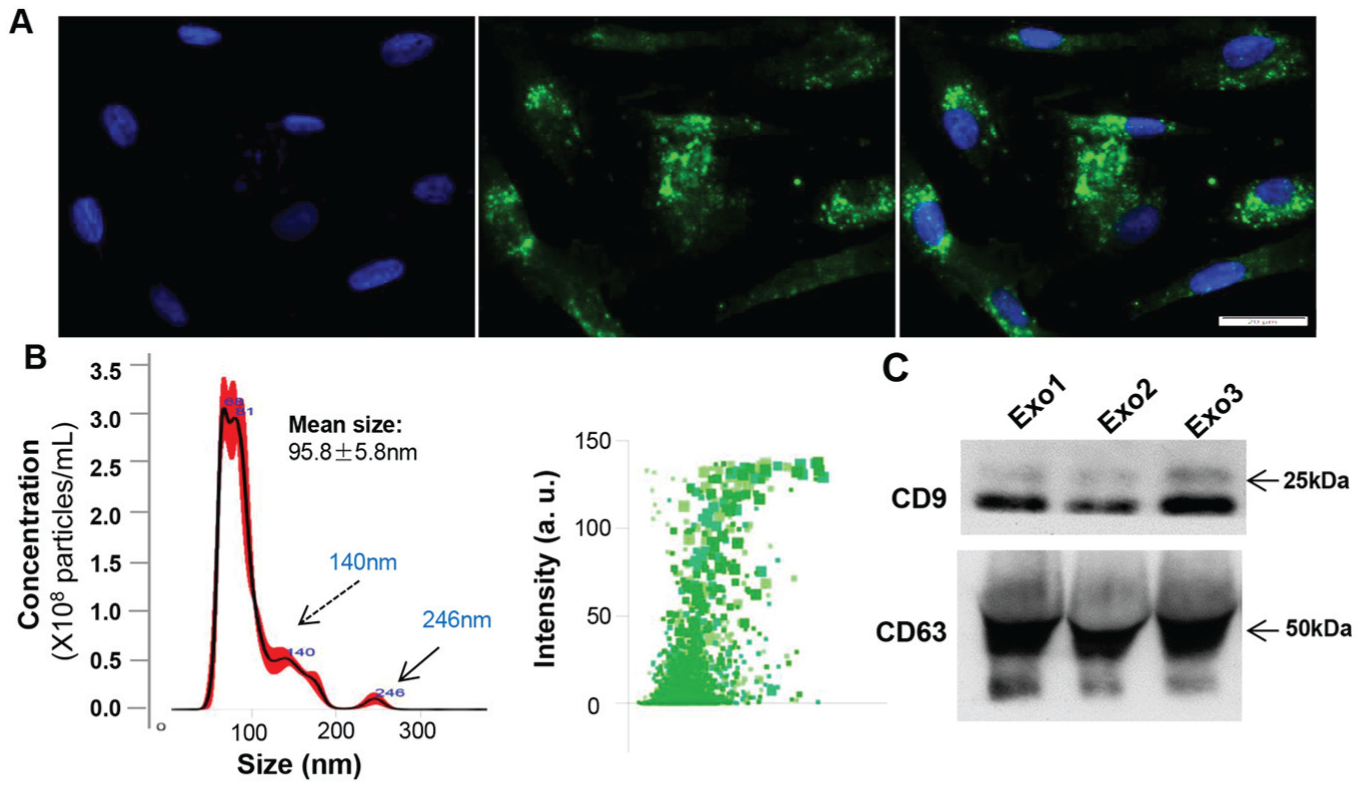
Purification and characterization of exosomes from culture media of gingival mesenchymal stem cells (GMSCs). (
GMSC/SIS-ECM and Exosome/SIS-ECM Promoted Recovery of Tongue Epithelium Papillae
To determine the effects of combinatorial GMSC/SIS-ECM and exosome/SIS-ECM on lingual papillae regeneration, SIS-ECM, GMSC/SIS-ECM, and exosome/SIS-ECM were transplanted onto the critical-sized tongue defects in 3 groups of animals. The healing of the tongue defects was individually observed at different time points. Similar to our previous report (Xu et al. 2017), all rats were able to survive after the surgery without obvious body weight loss during the experimental process (data not shown), and the transplanted SIS-ECM scaffold was completely resorbed by day 7 after transplantation (Fig. 2A). Until day 7 postsurgery, the wound margins in the GMSC/SIS-ECM group exhibited faster epithelial migration and wound closing and less scarring as compared with the SIS-ECM and exosome/SIS-ECM groups (P < 0.05; Fig. 2A, B). By day 14 postsurgery, clinical morphology of the tongue indicated that the GMSC/SIS-ECM and exosome/SIS-ECM groups showed less contraction and maintained the overall tongue shape without deformity as compared with the SIS-ECM control group. By day 28 postsurgery, the GMSC/SIS-ECM and exosome/SIS-ECM groups demonstrated better reepithelialization than the SIS-ECM control group, whereas the GMSC/SIS-ECM group appeared to display better epithelial restoration than that of the exosome/SIS-ECM group. By day 56 postsurgery, the GMSC/SIS-ECM and exosome/SIS-ECM groups showed comparably complete epithelial regeneration, while the SIS-ECM group still maintained some immature repaired epithelial tissue (Fig. 2A).
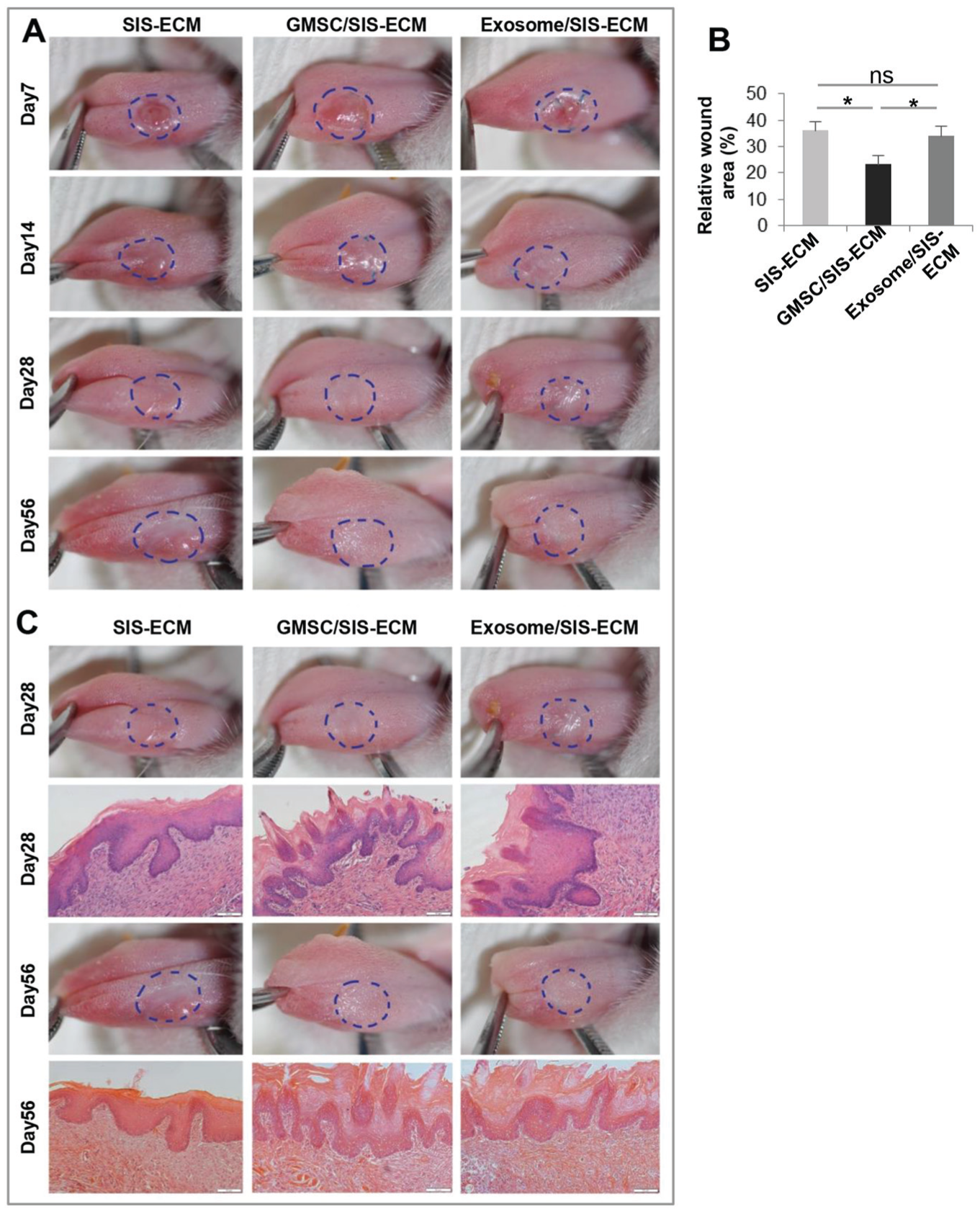
Integration of GMSC/SIS-ECM and exosome/SIS-ECM promoted regeneration of tongue lingual papillae. (
Histologic analysis indicated that the GMSC/SIS-ECM and exosome/SIS-ECM groups displayed more restoration of papillae and keratinized tissues in comparison with the SIS-ECM control (Fig. 2C). By day 56 postsurgery, the regenerated papillae in the GMSC/SIS-ECM and exosome/SIS-ECM groups became more regularly arranged than those at day 28 postsurgery. However, no obvious differences in the histology of the defect areas between day 28 and day 56 postsurgery were observed in the SIS-ECM control group (Fig. 2C). These findings suggest that GMSC/SIS-ECM and exosome/SIS-ECM groups were able to promote the reepithelialization of tongue dorsal defect and stimulate the lingual papillae regeneration.
GMSC/SIS-ECM and Exosome/SIS-ECM Promoted Taste Bud Regeneration
One of the key functions of dorsal tongue is to detect and conduct taste stimulation to the central nervous system, which is based on the mature structure and innervation of taste buds that reside in lingual papillae. Type II taste cells are critical for taste sensation and can respond to most taste stimuli and use ATP as a neurotransmitter to signal to sensory nerves (Barlow 2015). We then determined the expression of phospholipase C-β2 (PLCβ2), a marker for type II taste bud cells, and UCH-L1(PGP9.5), a pansensory neuronal marker (Yoshida et al. 2017), in the regenerated lingual papillae. Our results showed that the expression levels of UCH-L1 and PLCβ2 in the regenerated epithelial layers of rats treated with GMSC/SIS-ECM and exosome/SIS-ECM were significantly increased as compared with those in the SIS-ECM control group at day 28 postsurgery (P < 0.01), wherein the elevated expression of UCH-L1 and PLCβ2 induced by GMSC/SIS-ECM was more pronounced than that by exosome/SIS-ECM (P < 0.05; Fig. 3A, B).
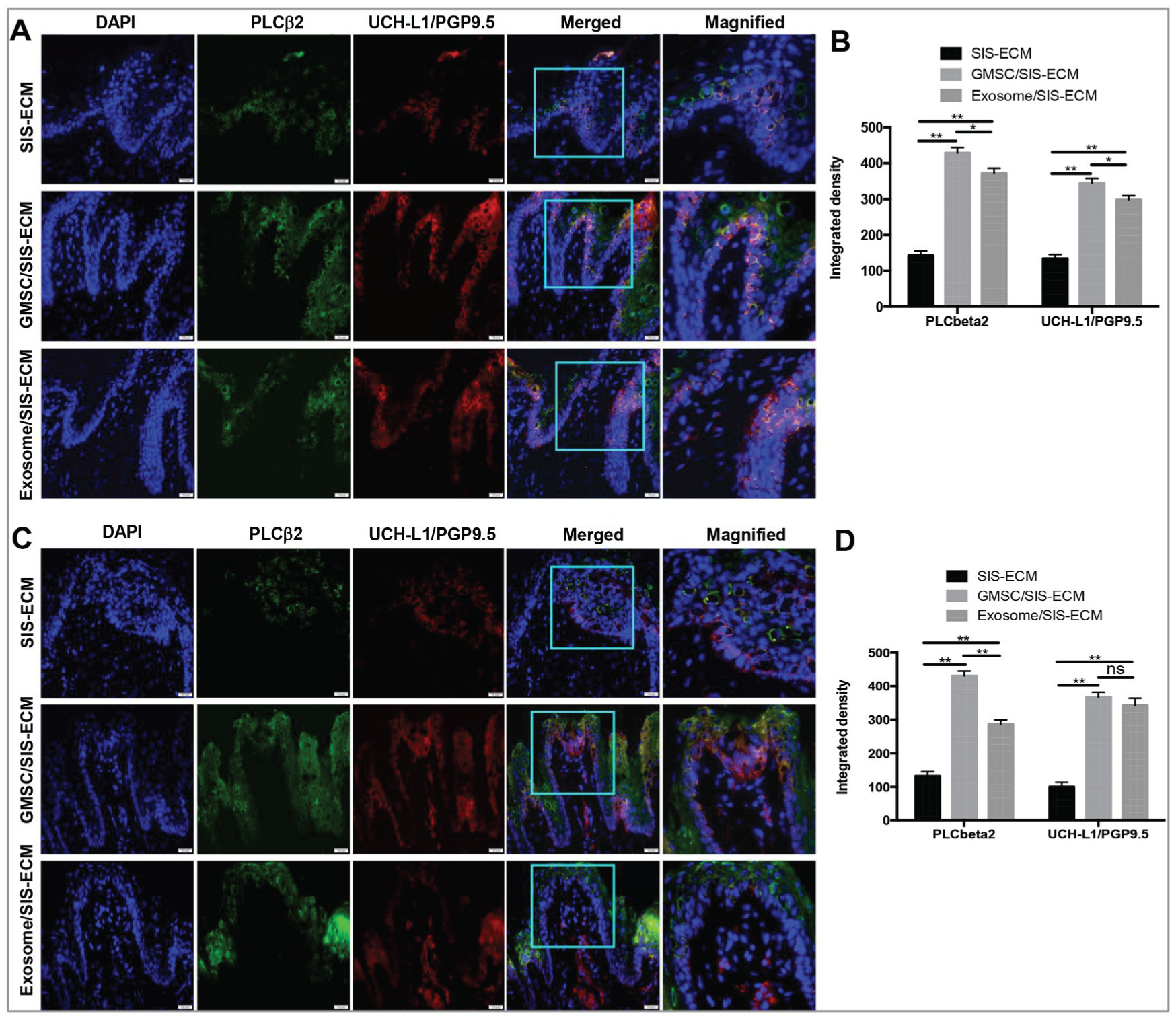
GMSC/SIS-ECM and exosome/SIS-ECM promoted taste bud regeneration. Coexpression of PLCbeta2 (green) and UCH-L1/PGP9.5 (red) at local defect areas at (
By day 56 postsurgery, the signals of PLCβ2 and UCH-L1 were significantly enhanced in the underlying connective tissue in the GMSC/SIS-ECM and exosome/SIS-ECM groups versus the SIS-ECM control group (P < 0.01), whereby the inductive effect of GMSC/SIS-ECM on PLCβ2 expression was relatively more robust than that of exosome/SIS-ECM (P < 0.05; Fig. 3C, D). In addition, we determined the expression of nucleoside triphosphate diphosphohydrolase-2 (NTPDase 2), a marker for type I taste bud cells (Bartel et al. 2006; Vandenbeuch et al. 2013), and aromatic L-amino acid decarboxylase (AADC), a marker for type III taste bud cells (Kotani et al. 2013). Our results indicated that the GMSC/SIS-ECM and exosome/SIS-ECM groups, at day 56 postsurgery, showed increased immunoreactivity signals of NTPDase 2 and AADC expressions in comparison with the those of the SIS-ECM group (P < 0.01; Appendix Fig. 1A-D). Of note, GMSC/SIS-ECM exhibited a relatively stronger effect on induction of NTPDase 2 than exosome/SIS-ECM (P < 0.05) (Appendix Fig. 1A, B). These results indicated that implantation of GMSC/SIS-ECM or exosome/SIS-ECM could facilitate restoration of epithelial layers and lingual papillae and regeneration of taste buds within the defect areas, whereas GMSC/SIS-ECM appeared to exhibit an overall better regenerative effect than exosome/SIS-ECM.
GMSC/SIS-ECM and Exosome/SIS-ECM Promoted Taste Bud Reinnervation and BDNF Expression
Our recent studies showed that GMSCs have the potential to promote regeneration of facial nerve defects in rats (Zhang et al. 2018). We then determined whether GMSCs or GMSC-derived exosomes could promote peripheral nerve regeneration within tongue defect areas, an essential component for functional innervation of regenerated taste buds. Our results showed that by day 56 postsurgery, the expression level of neurofilament, a neuronal marker of axons, was significantly increased in areas of tongue defect healing of the GMSC/SIS-ECM and exosome/SIS-ECM groups as compared with the SIS-ECM control group (P < 0.01; Fig. 4A, B). Of note, the expression signals of neurofilament were organized in a more linear structure/shape in the GMSC/SIS-ECM group, suggesting that the regenerated nerves were more mature than those of other 2 groups (Fig. 4A). According to previous studies, most fungiform papillae nerve fibers with P2X2 and P2X3 receptors were from the chorda tympani nerve (Ishida et al. 2009), which typically innervates taste bud cells (K8+P2X3+; Kumari et al. 2017). Our results further showed increased immunoreactivity of P2X3+ and numbers of K8+P2X3+ taste bud cells in regenerated taste buds from groups implanted with GMSC/SIS-ECM and exosome/SIS-ECM as compared with the SIS-ECM control group (P < 0.01; Appendix Fig. 2), suggesting that the regenerated nerves reinnervating taste bud cells were derived from the chorda tympani nerve.
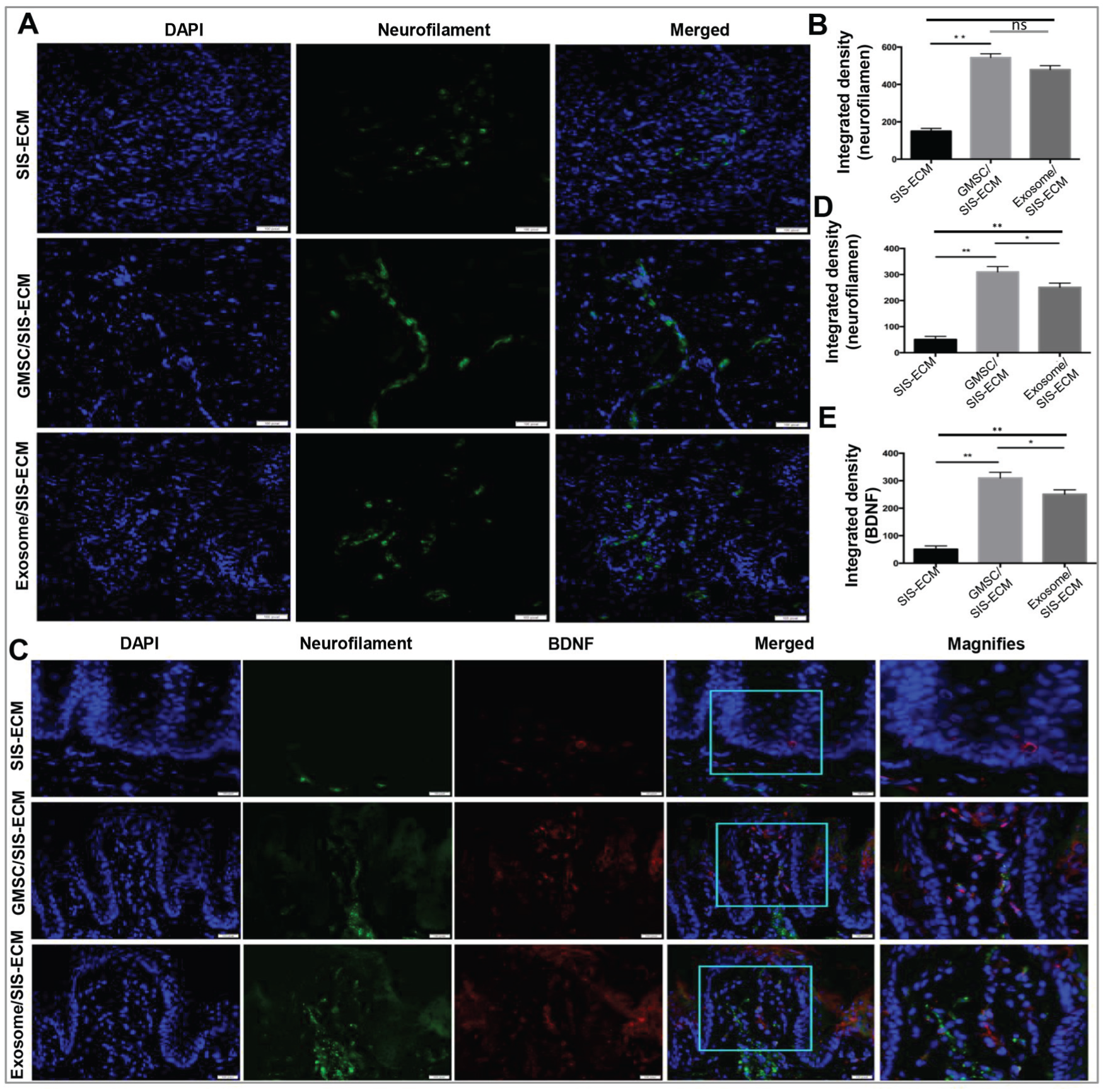
GMSC/SIS-ECM and exosome/SIS-ECM promoted nerve regeneration within defect areas. (
Previous studies indicated that nerve-derived BDNF plays an important role in the development and maintenance of taste buds. We then determined the expression of BDNF in nerve fibers in areas of tongue defect healing. By day 56 postsurgery, the expression levels of neurofilament and BDNF were increased in regenerated papillae in the GMSC/SIS-ECM and exosome/SIS-ECM groups as compared with the SIS-ECM group (P < 0.01; Fig. 4C–E), and the overlapped signals of neurofilament and BDNF expressions within the regenerated papillae were also extended into the underlying connective tissues (Fig. 4C). Of note, GMSC/SIS-ECM exhibited relatively stronger inductive effects on the expression of neurofilament and BDNF than did exosome/SIS-ECM (P < 0.05; Fig. 4D, E).
Most recently, several lines of evidence demonstrated the critical role of the Hedgehog (Hh) signaling pathways—particularly, Sonic Hedgehog (Shh) ligand signaling—in the development, homeostasis, and maintenance of taste bud organs (Castillo-Azofeifa et al. 2017; Kumari et al. 2017; Mistretta and Kumari 2017). We then determined the expression of the Shh ligand in the tongue defect areas at 14 d postsurgery. Our results showed that the expression levels of Shh were elevated in the tongue defect areas of animals implanted with either GMSC/SIS-ECM or exosome/SIS-ECM as compared with SIS-ECM (P < 0.01), whereby the increased immunoreactivity signal of Shh was mainly localized within the submucosal layers (Appendix Fig. 3).
Increased BDNF Expression in Epithelial Progenitor and Taste Bud Cells
Since the proliferation and self-renewal of CK14+ epithelial stem cells at the basal layer and CK8+ progenitor cells within a taste bud are essential for the development and regeneration of taste buds (Okubo et al. 2009; Li and Zhou 2012), we then determined the expression of CK14 and CK8 in the regenerated epithelium. We found that by day 7 postsurgery, the expression of CK14 within the basal epithelial layer in the GMSC/SIS-ECM and exosome/SIS-ECM groups was significantly higher than that in the SIS-ECM control group (P < 0.01; Fig. 5A, B). Accordingly, the expression level of BDNF in the epithelial layers was higher in the GMSC/SIS-ECM and exosome/SIS-ECM groups than in the SIS-ECM control group (P < 0.01). By day 14 postsurgery, the expression of CK8, a general marker for intragemmal taste bud cells, was significantly increased in the GMSC/SIS-ECM and exosome/SIS-ECM groups versus the SIS-ECM group (P < 0.01; Fig. 5C, D). Of note, GMSC/SIS-ECM exerted even stronger inductive effects on the expression of BDNF, CK14 (P < 0.01), and CK8 (P < 0.05) than did exosome/SIS-ECM (Fig. 5B, D). These findings suggest that an elevated expression of BDNF within the epithelial layers is closely associated with the increased expression of CK14 and CK8, thus suggesting that the increased BDNF expression mediated by GMSCs or GMSC-derived exosomes may contribute, at least in part, to their regenerative effects on taste bud regeneration in the tongue defect model in rats.
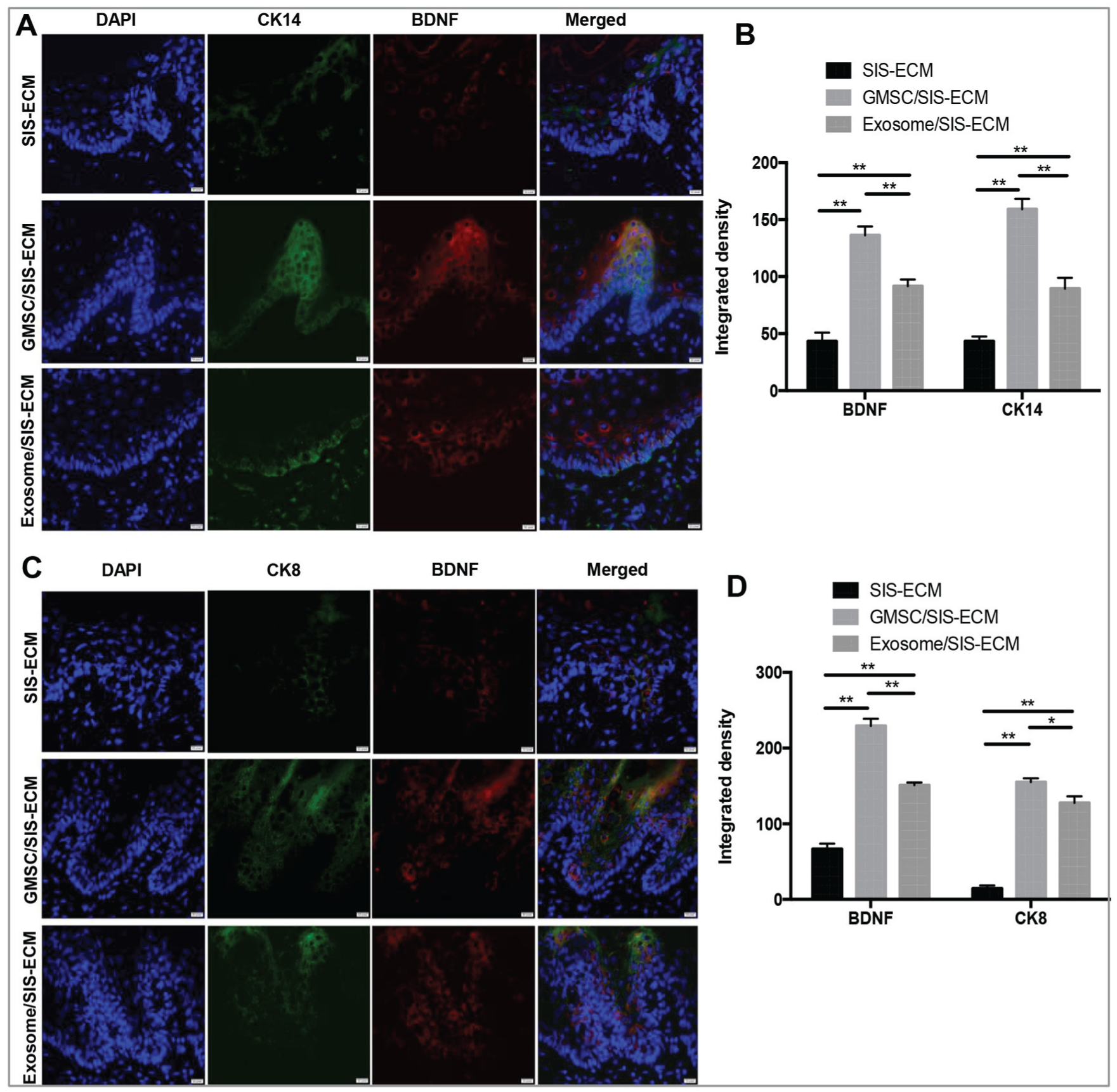
GMSC/SIS-ECM and exosome/SIS-ECM promoted regeneration of CK14+ basal epithelial cells and CK8+ taste bud progenitor cells coexpressing BDNF. (
Discussion
In recent years, tissue engineering and regenerative medicine (TE/RM) have shown great promises in tissue reconstruction (Kozlik and Wojcicki 2014). The TE/RM approaches usually involve the combinatorial use of various types of biomaterials, stem cells, or cell products (Rose and De Laporte 2018). Among the biomaterials, decellularized small intestine submucosa is one of the most commonly used acellular scaffolds in tissue regeneration (Andrée et al. 2013). Among the different types of stem cells, MSCs represent a subpopulation of postnatal progenitor cells of different tissue origins, such as bone marrow, adipose, and dental tissues (Zhang, Nguyen, Yu, and Le 2012; Loebel and Burdick 2018). Due to their multipotent differentiation capability, secretion of a myriad of bioactive factors, and immunomodulatory functions, MSCs have been widely explored in preclinical and clinical settings as cell-based therapies for a wide spectrum of diseases and tissue regeneration (Xu et al. 2017; Zhang et al. 2018). GMSCs, a subpopulation of MSCs derived from the easily accessible gingival tissues, are also multipotent and possess immunomodulatory functions (Zhang, Nguyen, Yu, and Le 2012). Our previous studies demonstrated that GMSCs could promote skin and oral wound healing in mice by facilitating epithelial regeneration (Zhang et al. 2010; Zhang, Nguyen, Shi, et al. 2012). Most recently, we showed that local implantation of GMSC/SIS-ECM promoted tongue muscle regeneration in a critical-sized tongue defect model in rats (Xu et al. 2017). Using the same tongue defect model in rats, we herein further showed that implantation of SIS-ECM scaffolds laden with GMSCs obviously facilitated lingual papillae reconstruction and taste bud regeneration, as evidenced by increased expression of NTPdase 2, PLC-β2, and AADC—3 markers for type I, II, and III taste bud cells, respectively (Bartel et al. 2006; Kotani et al. 2013; Vandenbeuch et al. 2013). Additionally, we found that implantation of GMSC/SIS-ECM significantly elevated the expression of the general neural fiber markers, UCH-L1/PGP9.5, and neurofilaments, as well as the expression of the chorda tympani nerve marker P2X3 receptor (Ishida et al. 2009), in areas of tongue defect healing. These findings suggest that the regenerated taste bud cells were reinnervated by the chorda tympani nerve (Ishida et al. 2009; Kumari et al. 2017).
CK14+ epithelial progenitor cells represent the pool for differentiated taste bud cells as well as keratinocytes (Okubo et al. 2009; Li and Zhou 2012), and CK8 is a general marker for intragemmal cells (Venkatesan et al. 2016), while both types of cells play an essential role in the development and regeneration of taste buds. In the current study, we showed that local implantation of GMSC/SIS-ECM could significantly increase the expression of CK14 and CK8 in the regenerated papillae as compared with SIS-ECM control, suggesting that GMSCs facilitated taste bud regeneration possibly by promoting the regeneration of these epithelial progenitor compartments.
Previous studies implicated an important role of BDNF expression in the development and maintenance of taste buds, which can be detected within the epithelial layer and underlying nervous tissues of taste buds (Farbman et al. 2004; Meng et al. 2015; Meng et al. 2017). Most recently, several lines of evidence showed that Hh signaling, particularly the Shh from nerves and epithelium, plays a key role in taste bud development, homeostasis, and maintenance (Castillo-Azofeifa et al. 2017; Kumari et al. 2017; Mistretta and Kumari 2017). In the present study, we showed that when compared with SIS-ECM control, implantation of GMSC/SIS-ECM obviously increased BDNF expression not only in CK14+ basal epithelial progenitor cells and CK8+ intragemmal cells but also in neurofilament+ nerve fibers in the submucosal connective tissues within areas of tongue defect healing. Meanwhile, we found that implantation of GMSC/SIS-ECM clearly increased Shh expression, whereby the increased Shh signals were mainly localized within the submucosal connective tissues. These findings support the notion that GMSCs enhanced taste bud regeneration and innervation, at least in part, by upregulating BDNF and Shh expressions in taste bud cells and non–taste bud cells. However, further mechanistic studies—for example, by the use of 2- and 3-dimensional organoid models of epithelial or taste bud cells—are warranted to explore the mechanisms whereby GMSCs upregulate BDNF and Shh expressions.
Most recently, a growing body of evidence has shown that exosomes, one of the key secretory products of MSCs, exert therapeutic effects comparable to those of parental MSCs in tissue repair via intercellular transfer of various bioactive components to modulate the activity of recipient cells (Phinney and Pittenger 2017). Additionally, exosomes are relatively stable and can easily pass the biological barriers, thus avoiding the risks associated with the use of stem cells (Nooshabadi et al. 2018). Therefore, MSC-derived exosomes may provide a novel paradigm for cell-free therapy in TE/RM. Interestingly, when compared with SIS-ECM control, local implantation of SIS-ECM scaffolds laden with GMSC-derived exosomes also significantly enhanced the expression CK14 and CK8, taste bud cell markers (type I, II, and III: NTPdase 2, PLC-β2, and AADC, respectively), nerve fiber markers (UCH-L1/PGP9.5, neurofilaments, and P2X3 receptor) as well as the expression of 2 key trophic factors (BDNF and Shh) in the reconstructed lingual papillae layers and the submucosal connective tissues within the areas of tongue defect healing. These findings suggest that GMSC-derived exosomes also possess regenerative potentials on reconstruction of lingual papillae and taste buds. However, it is noteworthy that GMSC/SIS-ECM appeared to exert an overall better beneficial effect on the reconstruction of lingual papillae structure and taste bud regeneration, suggesting that some nonexosomal factors secreted by GMSCs may also play a role in facilitating taste bud regeneration. The relatively lower efficacy of GMSC-derived exosomes on taste bud regeneration than that of GMSCs might also be due to the short half-life of exosomes. Therefore, further studies are warranted 1) to determine whether multiple treatments with exosomes can improve their therapeutic effects on taste bud regeneration and 2) to identify which types of nonexosomal factors and components existing within GMSC-derived exosomes have contributed to their regenerative potentials, respectively.
In summary, we have shown that transplantation of combinatorial GMSC/SIS-ECM and exosome/SIS-ECM facilitated lingual papillae reconstruction and bud regeneration in rats. However, the present study has several limitations: a lack of electrophysiologic or behavioral analysis of the regenerated taste buds; a lack of serial observations on the dynamic changes in the expression of key genes involved in taste bud regeneration; a lack of a mechanistic study on how GMSCs and exosomes regulate the expression of BDNF and Shh in taste bud cells and other types of cells; and last, the necessity to optimize the dose and frequency for exosome treatment. Future studies are warranted to further address these important points. Taken together, our current findings support the notion that GMSCs represent an easily accessible source of stem cells for fabricating TE/RM products with a great promise for clinical application in tongue reconstruction and taste bud regeneration after ablative surgery of tongue cancer.
Author Contributions
Y. Zhang, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; S. Shi, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; Q. Xu, contributed to conception, design, data acquisition, analysis, and interpretation, critically revised the manuscript; Q. Zhang, A.D. Le, contributed to conception, design, and data interpretation, drafted and critically revised the manuscript; R.M. Shanti, contributed to conception, design, and data interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
DS_10.1177_0022034518804531 – Supplemental material for SIS-ECM Laden with GMSC-Derived Exosomes Promote Taste Bud Regeneration
Supplemental material, DS_10.1177_0022034518804531 for SIS-ECM Laden with GMSC-Derived Exosomes Promote Taste Bud Regeneration by Y. Zhang, S. Shi, Q. Xu, Q. Zhang, R.M. Shanti and A.D. Le in Journal of Dental Research
Footnotes
This work was supported by Peter Geistlich Research Awards of Osteo Science Foundation (Q.Z. and A.D.L.), the Schoenleber funding support (A.D.L.), Oral and Maxillofacial Surgery Foundation Research Grant (Q.Z., R.M.S.).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.